Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Hücre Zarında Morfolojik Farklılaşmalar

2
Hücreler yaptıkları emme, salgılama v. s
Hücreler yaptıkları emme, salgılama v.s. görevleriyle ilgili olarak, bulundukları yerlerde göre bazı morfolojik yüzey değişiklikleri gösterirler. Bu değişiklikler hücrelerin bazal, lateral ve apikal yüzeylerinde meydana gelir.

3
Bazal Plâzma Zarı Çoğunlukla hücrenin kaidesine rastlayan bazal yüzey, bazal lamina üzerine oturmuş olup düz bir şekilde uzanır. Fakat bazı hücrelerde göreve uygun değişiklikler meydana gelebilir. Bazal plâzma zarında en çok görülen değişiklik zarın içeri doğru meydana getirdiği katlanmalardır. Bu katlanmalar hücrenin 1/3 derinliğine kadar gider. Deniz kuşlarının tuz bezlerinde hücrelerin tepesine kadar, Rhodnius 'un Malpighi hücrelerinde 3-5 mikron yüksekliğe kadar uzanır.

4
Katlanmalar bölme halindedir. Bölmeler dallanabilir ve birleşebilir.
Bölmelerin arasında sitoplâzma kompartımanları meydana gelmiştir. Bu kompartımanlarda bol miktarda mitokondri bulunur. Mitokondriler bölme yüzeyine paralel uzanırlar. Bölme sisteminin gelişme derecesi hücre ile çevresi (kan, doku sıvısı) arasındaki madde taşınmasının önemine bağlıdır. Bu katmanlarla hücrenin kaide yüzeyi genişletilmiş olur.
 arasındaki madde taşınmasının önemine bağlıdır. Bu katmanlarla hücrenin kaide yüzeyi genişletilmiş olur.")
5
M M Bazal lamina Bazal zar katlanmaları ve aralarında mitokondriler

6
Lateral Plâzma Zarı Plâzma zarları, yan yana duran komşu hücreler arasında çok bitişik değildir. Aralarında bir hücreler arası (intersellüler) açıklık bulunabilir. Hücreler arası glikokaliks ile dolu olup bu madde polisakkaritlerden, siyalik asitten ve asit mukopolisakkaritlerden (glükozamino-glikanlar) oluşur. Yan yana gelen hücrelerin zarları hücrelerin birbirine bağlanmasını sağlamaya yarayan katlanmalar. Yan yana gelen hücreler arasında gerçek hücreler arası boşluklar da meydana gelebilir.
 açıklık bulunabilir. Hücreler arası glikokaliks ile dolu olup bu madde polisakkaritlerden, siyalik asitten ve asit mukopolisakkaritlerden (glükozamino-glikanlar) oluşur. Yan yana gelen hücrelerin zarları hücrelerin birbirine bağlanmasını sağlamaya yarayan katlanmalar. Yan yana gelen hücreler arasında gerçek hücreler arası boşluklar da meydana gelebilir.")
7
Hücre Hücre Hücre zarı Hücreler arası aralık Hücre zarı İki hücre zarı ve aralarında hücreler arası aralık

8
Lateral zar katlanmaları

9
Hücreler arası aralık Zar Zar M İki hücre zarı ve aralarında hücreler arası aralık

10
Zar kıvrımları Yan yana hücreler ve zarları, zar kıvrımları

11
Lateral Plâzma Zarında Birleşme Kompleksleri
Hücreler daimi olarak birarada bulundukları zaman bazen iki hücrenin temas ettiği yerlerdeki plâzma zarları arasında çeşitli özel birleşme yerleri teşekkül eder. Omurgalı hücrelerinde; Dezmozom, Kapalı birleşme yeri, Aralıklı birleşme yeri (gap junction) gibi üç özel tip birleşme görülür. Bitki hücrelerinde plâzmodezma denilen bir birleşme tipi görülür. Hücrelerin birbirlerine mekanik yapışıklılığının sebebi aralarındaki dezmozomlardır.
 gibi üç özel tip birleşme görülür. Bitki hücrelerinde plâzmodezma denilen bir birleşme tipi görülür. Hücrelerin birbirlerine mekanik yapışıklılığının sebebi aralarındaki dezmozomlardır.")
12
İki hücrenin birbirine mekanik olarak bağlanması Düğme (nokta) dezmozomlar (macula adherens) olarak bilinen yapılardır. Bir nokta dezmozom, her biri iki komşu hücre zarı içinde bulunan iki stoplazmik plakadan oluşur. Hem plakalardan hem de flamentlerden oluşan Kemer dezmozomlar (zonula adherens) ile nokta dezmozomlar arasında yüzeysel bir benzerlik vardır. Hücre-hücre tutunmasında bir rolü yoksa da hücrenin iç kısmını çepe çevre saran ve kasılabilir nitelikte fibril içeren plakalar, hücre için bir iç destek oluşturur.
 ile nokta dezmozomlar arasında yüzeysel bir benzerlik vardır. Hücre-hücre tutunmasında bir rolü yoksa da hücrenin iç kısmını çepe çevre saran ve kasılabilir nitelikte fibril içeren plakalar, hücre için bir iç destek oluşturur.")
13
Hücreler, komşu hücre zarlarındaki özel transmembran proteinlerinin birbirlerine bağlanmasıyla sıkı bağlantılarla da (zonula occludens) birbirine tutunur. Hücreler arasında böylece hiç boşluk kalmaz. Hücreler bunlardan başka delik-geçit bölgeleri (neksus) aracılıyla da birbirlerine bağlanabilir. Bu iki hücrenin sıralı özdeş kanal oluşturmasıyla olur.
 aracılıyla da birbirlerine bağlanabilir. Bu iki hücrenin sıralı özdeş kanal oluşturmasıyla olur.")
14
Delik-geçit bölgeleri iki sinir hücresini elektriksel olarak bağlayabilir. Bu hücreler sadece sinyal oluşturan elementlerin geçişini sağlayacak şekilde işlev görürler. Gelişmekte olan hücrelerde çok yaygındır ve başlangıçta hücrelerin birbirine yapışmalarında ve düzenlenmelerinde önemli rol alır. Sonuçta, hem mekanik kuvvet hem de hücreler arasında bazı özel maddelerin paylaşılması söz konusudur.

15
Bitkilerde ise bitki hücre duvarında komşu hücre zarının birbiriyle birleştikleri bölgelerde plazmodezmata denen özelleşmiş açıklıklar vardır. Bunların bazıları açıkken, bazıları çift zarlı bir engel olarak kalıp, çözücü ve çözünen maddelerin hücreler arasındaki hareketlerini kontrol etmede önemli rol oynar.

16
Plazmodezma Hücre duvarı Vakuol Sitoplazma Plazmodezma Bitkilerde hücre duvarı

17
GER Hücre duvarı Dezmotüpçük Bitki hücre duvarında plazmodezmata

18
Dezmozomlar iki tip olur. Bunların ince yapıları aynıdır.
Nokta dezmozomu‘lar daha çok görülürken, kemer dezmozomu’lara daha az rastlanır. Nokta dezmozomu iki hücrenin plâzma zarları arasında yer yer görülür. Dezmozomlar yan yana iki hücreyi sıkıca birarada tutar. Mekanik baskılara maruz dokularda sık rastlanırlar. İki hücrenin plâzma zarları değişmemiştir ve zarlar arasında A° kadar olan bir aralık bulunur. Burası ipliksi bir madde ile doludur ve hücreler arası bir çimento vazifesi görür.

19
Dezmozomlar, madde protein ve karbohidrat kapsar
Dezmozomlar, madde protein ve karbohidrat kapsar. Muhtemelen glikoproteinler ve glikoz-aminoglikanlar bulunur. Nokta dezmozomun sitoplâzma tarafında amorf bir matriks şeklinde elektronca yoğun bir madde bulunur, bu sitoplâzmik plâk’ tır. 100 A° çapında iplik demetleri bu yapıdan zarın iç kısmına yaklaşır, ilmik yapıp sitoplâzmaya geri döner. Bunlar keratin'den yapılmış Tonofilament denir Bazı hücrelerde tonofilamentler sitoplâzmayı geçerek dezmozomun iç tarafından diğerinin iç tarafına uzanır (Transmembran bağ).
.")
20
Kemer dezmozomları basit yapılıdır
Kemer dezmozomları basit yapılıdır. Aktin'den yapılmış 70 A° çapındaki tonofilamentlere sahiptir. Tonofilamentler kemer dezmozomun bulunduğu yerde plâzma zarının hemen altındaki sitoplâzmada görülür. Bazı dezmozom yapıları tek taraflı olur. Buna yarım dezmozom (hemidezmozom) denir.
 denir.")
21
Sitoplazmik plak Tonoflamentler Dezmozom

22
Kaderin Epidermiste dezmozom Tutunma proteinleri Hücreler arası aralık
Sitoplazmik plağa bağlanmış keratin iplikler Hücreler arası aralık Epidermiste dezmozom Hücrelerin zarları

23
Dezmozomlar

24
Dezmozomlar

25
Dezmozomlar

26
Dezmozomlar

27
Tonoflament Yarım dezmozom (Hemidezmozom)
")
28
Kapalı birleşme yerlerinde yan yana gelen hücrelerin plâzma zarları çok sıkı bir şekilde temas halindedir. Temas yerinde hiçbir aralık kalmamıştır. Bu şekilde bir tıkanma meydana gelir. Kapalı birleşme yerleri hücreyi çepeçevre kemer gibi sarar. Bu yapıya zonula occludens denir. Bu yapılar, meselâ mesane gibi bir organın iç yüzünü kaplayan epitel hücrelerinin apikal tarafa bakan lateral zarları arasında meydana gelir.

29
Kapalı temas yerleri tek bir nokta halinde olmayıp aralıklı bir durum gösterebilir.
Birçok bezin ve bağırsak epitelinde olduğu gibi, apikal bazal yönde teşekkül eden bu kompleksin başında bir kapalı birleşme yeri bulunur. Kapalı birleşme bölgesi hücreyi kemer gibi sarar ve uzunluğu A° kadardır. Bu bölgeyi A° uzunluğunda olan bir kemer dezmozom bölgesi izler. Hücreleri kemer gibi saran bu bölgeden sonra tipik dezmozom olan nokta dezmozomu yer alır.

30
Kapalı birleşme bölgeleri (Tight junctions)
")
31
Kapalı birleşme bölgeleri (Tight junctions)
")
32
Kapalı birleşme yerleri

33
Aralıklı birleşme yerleri yan yana gelen zarlar arasında yer yer görülür.
Bu bölgede iki hücrenin zarları hiçbir zaman tamamen birleşmezler. Daima aralarında A° olabilen bir aralık kalır. Bu aralık ince bağlantılarla birleşmiştir. İnce bağlantı yerlerinde 70 A° çapında konnekson denen partiküller bulunur (konneksin denilen transmembran –integral- proteindir). Ortasında 15 A° çapında annulus denen açıklık bulunur.
. Ortasında 15 A° çapında annulus denen açıklık bulunur.")
34
Konneksonun altı alt birimi bir zarda, altı alt birimi diğer zarda yer alır.
Konnekson iki zarda karşılıklı zar boyunca yer alır ve her iki tarafta sitoplâzmaya ve ekstrasellüler aralığa uzanırlar. Ortalarındaki açıklık bir hücrenin sitoplâzmasından diğerine uzanan devamlı porlar meydana getirir.

35
Aralıklı birleşme yerleri (Gap junction)
Konnekson Zar Zar Aralıklı birleşme yerleri (Gap junction)
")
36
Konneksin Annulus Aralıklı birleşme yerleri (Gap junctions) kanalları
 kanalları")
37
Hücre yüzeyi Bazal lamina

38
Apikal Plâzma Zarı Hücrelerin apikal yüzeyinde sadece görev sırasında yani hücre aktif halde iken meydana gelen modifikayonlar olabilir ve bunlar sabit değildir. Fakat sabit olan farklılaşmalar da görülür. Hiçbir hücrenin serbest yüzeyi düz değildir. Kıvrımlara genel olarak mikrovillüs adı verilir. Mikrovillüsler birkaç tane veya çok fazla sayıda olabilir.

39
Mikrovillüs sayısı az olduğu zaman uzunluğu, çapı ve şekli düzgün olmayan uzantılar halinde görülür.
Mikrovillüslerin sayısı artınca boyu ve çapı düzgün bir durum alır. Mikrovillüsler uçları yuvarlak olan ve kenarları birbirine paralel uzanan çıkıntılardır. Uzunlukları birkaç yüz Angströmden 10 mikrona kadar değişir. Çapları A° olur.

40
Mikrovillüsler Plazma zarı

41
Mikrovillüsler

42
Bazı mikrovillüslerinin kaidesinde küçük keseler şeklinde çöküntüler olur.
Bu çöküntülerden pinositoz yolu ile fazla miktarda sıvı içeri alınır Mikrovillüsleri örten plâzma zarı da birim zar yapısında olur. Kalınlığı A° kadardır. Mikrovillüslerin üzerinde düzgün ve yoğun bir şekilde dağılmış ipliksi bir yapıda, glikoprotein makromolekülleri halinde bulunan dış örtüler bulunur.

43
Mikrovillüslerin ortasında mikrofilamentler'den oluşan bir göbek kısmı bulunur.
Bu yapılar hücrenin apikal kısmında bulunan aktin filament ağına enine filamentlerle bağlanır. Göbek filamentleri ayrıca mikrovillüslerin lateral zarlarına ince ipliklerle bağlıdırlar. Mikrovillüslerin ortasındaki yapılar aktin filamentleri olduğu için kasılma özelliği vardır. Mikrovillüslerle hücrenin emme yüzeyi genişler.

44
Mikrovillüslerin boyuna ve enine kesitleri

45
Mikrovillüsler Mikrovillüsler Mikrofilamentler (Aktin filamentleri)
İntermedyer filamentler Mikrovillüsler

46
Siller (Kirpikler) ve Flagellumlar (Kamçılar)
Hücre zarından meydana gelen bir yapılardır. Bazı omurgalı spermatozoonlarının kamçıları hariç çeşitli hayvan türlerinde yapılışları aynıdır. Siller genellikle hücre zarında tek olarak, bazı bir hücrelilerde ise (Clamydomonas reinbardtti ) aynı yerden çift olarak çıkar.
 aynı yerden çift olarak çıkar.")
47
Sillerden Meydana Gelen Yapılar
Sillerin en fazla değişmiş şekillerine omurgalıların fotoreseptör hücrelerinde rastlanır Boyun içinde bazal cisimcik uzanır ve dokuz çift periferal tüpçükten yapılmıştır. Merkez telleri yoktur. Periferal tüpçükler dış segmente geçtikten sonra sonlanırlar. Boyun bölgesinin kaidesinde bir sentriyol, iç segmentte ikinci bir sentriyol bulunur.

48
Miyelinli aksonlarda, başlangıçta aksonun çevresinde miyelin yoktur.
Hücre zarından farklılaşarak gelişen diğer bir yapı sinir hücre aksonunda görülen Miyelin’dir. Sinir aksonları miyelinli veya miyelinsiz olurlar. Miyelinsiz aksonlar Schwann hücresi içinde gömülü bulunurlar. Schwann hücre zarı aksonun etrafını çevirir ve iki uçtan, zarın karşılaştıkları yerde birleşip bir boyun halinde kalır. Bu boyuna mezakson denir. Miyelinli aksonlarda, başlangıçta aksonun çevresinde miyelin yoktur. Gelişme ilerledikçe Schwann hücresinin zarında meydana gelen mezakson, aksonun çevresinde dönmeye başlar ve Schwann hücresinin sitoplâzmasını iterek ortak merkezli silindirler halinde tabakalar meydana getirir

49
Akson etrafında miyelin kılıf

50
Akson etrafında Schwann hücre zarı tarafından miyelin oluşturulması

51
Akson

52
Makromoleküllerin ve Partiküllerin
Hücreye Alınması Hücre, plâzma zarı ile hem katı parçacıkların yutulmasını hem de makromoleküllerin bir çeşit içilmesini yapar. Bu iki olaya fagositoz ve endositoz denir. Hücreden büyük miktarda madde çıkarılması ise ekzositoz adı verilen bir görevdir. Bu iki olay birbirinin zıddıdır. Endositozla hücre içine alınan parçalar daima bir hücre içi vaküole girerler. Hücrede teşekkül eden bu sitoplâzmik vaküole endozom denir.

53
Özelleşmiş bazı hücreler tarafından mikroorganizmaların veya katı parçacıklar gibi spesifik olmayanların yutulması veya yenmesi olayı fagositoz'dur. Fagositoz olayında, yutulacak parçacık plâzma zarı üzerine gelir. Zarda psödopod denilen yalancı ayaklar parçacığın üzerini çevirir. Karşılıklı gelen plâzma zarları birleşerek içinde yabancı parçacık bulunan bir vaküol teşekkül eder. Fagositozla teşekkül eden bu fagositik vaküol bir fagozom'dur. Fagositoz olayında, önce parçacıklar hücre zarına yapışır, sonra içeri alınır.

54
Katı besin maddesi FAGOSİTOZ Çekirdek Yalancı ayak Özel granüller

55
Fagositoz

56
Fagositoz olayı 4 aşamada tamamlanır.
Bu aşamalar kemotaksi (yönelme), yapışma, öldürme ve sindirme’dir Öldürülen obje daha sonra sekonder lizozom enzimleriyle parçalanır. Fagositoz olayında objenin partikül yüzeyinin ve kimyasal yapısının önemi büyüktür. Ancak patojen bakterilere karşı konukçu yüzeyinde serum antikorları meydana getirirler.
, yapışma, öldürme ve sindirme’dir. Öldürülen obje daha sonra sekonder lizozom enzimleriyle parçalanır. Fagositoz olayında objenin partikül yüzeyinin ve kimyasal yapısının önemi büyüktür. Ancak patojen bakterilere karşı konukçu yüzeyinde serum antikorları meydana getirirler.")
57
Fagositoz

58
Fagositoz yapan birçok hücrenin zarında antikorun kolayca bağlanabileceği Fc reseptörleri vardır.
Ayrıca konukçu tarafından serum proteini olarak meydana getirilen kompleman komponentleri fagositozu hızlandırır. Örnek, makrofajlar Amiplerde fagositoz olayı ile hayvanın amöboyid hareketi sıkıca bağlıdır. Amipte hücrenin beslenmesi bu yolla olur.

59
Birçok amipin yüzeyi dışardan ince filamentlerle örtülüdür veya glikozaminoglikandan oluşan bir örtü bulunur. Besin parçacıkları önce bu ekstrasellüler materyale yapışır, sonra yutulur. Bir çok alçak çok hücrelilerde, özel hücreler plâzma zarı ile fagosite ettikleri besini hücre içine alarak sindirirler ve böylece beslenirler. Fakat yüksek yapılı çok hücrelilerde fagositoz bir savunma olayıdır. Memelilerde granüllü lökositler ve makrofaj denilen fagositik hücreler bu görevi yapmaktadır.

60
Endositoz, pinositoz ve reseptör aracılığı ile endositoz olarak iki şekilde olmaktadır.
Bitki hücrelerinde endositoz görülmez. Sıvıların alınmasına pinositoz denir. Fagositozla bazen pinositoz arasındaki fark çok bariz değildir. Pinositik vaküoller, çapı yaklaşık 0.1 mikron kadar büyüklüklerde olur. Bu vaküollere pinozom denir. Pinositozla hücreye alınan materyal spesifik değildir.

61
Pinositoz

62
Çekirdek

63
Reseptör aracılığı ile yapılan endositozda materyalin spesifik olması gerekmektedir. Hücre zarında bulunan reseptörler hücre içine alınacak meteryali tanımak zorundadır. Ligand adını alan ekstrasellüler bir materyal hücre zarındaki reseptörle ligand-reseptör kompleksi'ni oluşturur. Ekstraselüler proteinler ve küçük partiküller, spesifiklik durumuna göre liganda sıkıca bağlanır.

64
Bu tip endositoz sırasında meydana gelen kesecik, Örtülü kesecik dıştan iki tip protein içeren bir örtü ile çevrelenmiştir. Bu kesecikler sitozol içine taşınır. Kesecik etrafındaki bu örtü daha sonra çözünerek sitozol’e karışır. Örtülü keseciğin esas proteini Klatrin’dir. Klatrin üçlü yapıya sahiptir. Klatrinin zara bağlanmasında Adaptinler adı verilen ikinci tip proteinler rol oynar. Adaptinler aynı zamanda trans-Golgiden ayrılacak keseciklerin yapısına girecek moleküllerin seçiciliğinde de rol oynar.

65
Reseptör aracılığı ile yapılan endositoz ve örtülü kesecikler
Örtü proteini Reseptör Örtülü kesecik Örtülü çukur Sitoplazma Reseptör aracılığı ile yapılan endositoz ve örtülü kesecikler

66
Endositozla kolesterolün hücreye alınması
Hücre dışı Apoprotein Kolesterol Örtülü çukur LDL kompleksi LDL reseptörü Sitoplazma Kesecik Klatrin Endositozla kolesterolün hücreye alınması

67
Plâzma zarı ile, aynı zamanda, ekzositoz yapılmaktadır.
Hücrede, bir zarla çevrili olarak salgı granülleri teşekkül eder ve salgılanır. Buna salgılama (sekresyon) denir. Fagositoz ve pinositoz olayları sonucu meydana gelen vaküol ve keseciklerde bulunan maddelerin, enzimlerle sindirilmesi sonucunda meydana gelen artık ürünler dışarı boşaltılır. Buna da boşaltım (ekskresyon) denir.
 denir. Fagositoz ve pinositoz olayları sonucu meydana gelen vaküol ve keseciklerde bulunan maddelerin, enzimlerle sindirilmesi sonucunda meydana gelen artık ürünler dışarı boşaltılır. Buna da boşaltım (ekskresyon) denir.")
68
Ekzositozda, dışarı verilecek maddenin çevresindeki zar, plâzma zarı ile birleşerek muhtevasını dışarı çıkarır. Golgi ile paketlenen salgı maddesi küçük kesecikler halinde Golgi sisternlerinden ayrılınca, önce yoğunlaşma vaküolleri'ni oluştururlar. Salgı granülleri yavaş yavaş hücrenin serbest yüzeyine yaklaşır. Bu arada, granülün içinde salgı olgunlaşır. Granülün zarı ile plâzma zarı yan yana gelirler. Zarların dış lipit tabakaları önce lipit miselleri teşkil eder ve sonra iki yapının zarı birleşir ve salgının dışarı atılacağı yerde bir kanal teşekkül eder.

69
Ekzositoz

70
Ekzositozun çeşitleri vardır.
Holokrin tipi salgılama yapan hücrelerde, önce hücre salgı granülleri ile dolar ve bütün hücre salgı haline dönüşür. Bu olayda hücre yozlaşır ve içindekinin tamamı dışarı bırakılır. Memeli derisinde, kıllara bağlı olan yağ bezleri bu tip salgılama yaparlar. Bu tip Dejeneratif salgılamadır. Birde hücrenin parçalanmadan kendisinin ürün olduğu salgılama şekli olan aktif holokrin salgılama vardır. Testis ve ovaryum bu aktif holokrin bezlerdir.

71
Merokrin tipi salgılama çok incelenmiştir
Merokrin tipi salgılama çok incelenmiştir. Sentezlendikten sonra bir zarla çevrilen proteinler veya glikoproteinler, bu zarın hücre zarı ile birleşmesi sonucu, içindekilerini hücrenin dışına bırakırlar. Birçok endokrin ve ekzokrin bezlerin salgı yapan hücreleri salgılarını bu tipte çıkarırlar. Apokrin tipi salgılamada sitoplâzmanın apikal yüzeyinden bir parçası kopup ayrılır. Önce apikal plâzma zarı büyür ve salgılama yapılınca apikal yüzeyde bir azalma meydana gelir. Süt ve ter bezlerinde salgılama bu tipte olur.

72
Sitopempis olarak adlandırılan diğer bir olayda, pinositozla teşekkül eden vaküol olduğu gibi değişikliğe uğramadan dışarı atılır. Bir taraftan alınan madde, öbür tarafa nakledildikten sonra, keseciğin zarı öbür taraftaki plâzma zarı ile birleşerek içindekini dışarı atmaktadır. Bu şekilde hücre bazı maddelerin taşınmasında aracı olmaktadır.

73
Maddelerin Zardan Geçişi

74
Hücre zarının, hücrenin korunmasını sağlama görevinin yanında, en önemli görevi maddelerin hücreye giriş ve çıkışını düzenlemesidir. Zarın lipit tabakaları zarın geçirgenliğinde önemli rol oynar.

75
DANIELLI ve DAVSON, zarda porlar bulunduğunu belirterek bu porların proteinle kaplı olduğunu ve maddelerin bu porlardan geçtiğini ileri sürmüşlerdir. Özellikle su ve suda çözünen küçük moleküller bu porlardan geçebilmektedir. Sıvı mozayik zar modelinde diasil gliseridlerin yağ asidi kuyruklarının termal olarak hareketli olduğu ve küçük polar maddelerin buralara girerek zardan geçebileceği düşünülmüştür.

76
Su girişi hipertonik veya hipotonik oluşuna göre,
Diğer maddeler ise, hücrenin fonksiyonlarının gerekliliğine göre seçici olarak hücreye girer ya da çıkar. Zar seçici geçirgen (selektif permeabl)’dir.
’dir.")
77
Yağlarda erimeyen maddelerin veya küçük polar maddelerin zarlardan geçişi 3 metoda göre yapılmaktadır.

78
1. Difüzyon Difüzyonda maddeler, zardan pasif olarak, yani bir enerji sarfı gerekmeksizin taşınırlar. Difüzyonun çeşitleri vardır.

79
I. Yoğunluğu yüksek olan bölgeden yoğunluğu düşük olan bölgeye geçiş
Genellikle suda çözünen bütün maddeler düşük konsantrasyonlu bölgelere doğru yayılma eğilimindedirler. Akciğerlerin alveollerinde oksijenin difüzyonu bunun en iyi bilinen örneğidir. Yoğun bir şeker çözeltisinin su ile temas edince, şekerin suya geçmesi verilebilir. Konsantrasyon ne kadar yüksek ise difüzyon da o kadar hızlı olur.

80
Zar kanalları ve taşıyıcılar
İyonlar Nörotransmitterler gibi Daha büyük moleküller Zar kanalları ve taşıyıcılar

81
Difüzyon sırasında, maddenin, zardan geçişi enerji kullanılmasını gerektirmeyen pasif bir taşıma olayıdır. Küçük ve yüklü iyonlar zarlardan difüzyonla geçerler. Fakat bu geçiş çok yavaş olur. Anyonlar katyonlara göre daha kolay geçerler. Monovalent maddelerin geçişi bivalentlerden daha kolaydır. Na+, K+, Cl–, H+, Mg+2, Ca+2 zarlardan geçişi en çok incelenen yüklü iyonlardır ve hücrenin fizyolojik aktivitesinde çok önemlidir.

82
Biyolojik sistemlerde difüzyon olayının devam ettirilebilmesi için zarı geçen maddenin geçtiği yerde birikmemesi gerekir. Meselâ, böceklerde, glukoz yağ cisimciklerinde bir disakkarit olan trehaloz şekline çevrilerek depolanır.

83
II. Elektrik yüküne göre difüzyon
İnorganik iyonların difüzyonu daha zordur ve sadece iyonların yoğunlukları geçişte yeterli olmayıp elektrik yükü farkları da rol oynamaktadır.

84
Elektrik yüklü madde, ancak yeterince elektrik yükü farkının olduğu yerde, iyon yoğunluk farkına göre difüzyon yapar Elektrik potansiyeli farkının –100 mv olması hücre zarları için önemli olup bu fark yüklü maddelerin geçişine yön verir. Potansiyel farkları ile zarın iki yanında bazı iyonların konsantrasyonlarında önemli farklar sağlanır. Meselâ, birçok hücrelerin iç çevresinde Cl– iyonu konsantrasyonunun az olması bu yolla temin edilir.

85
Tablo. Kasta iyon konsantrasyonu ve elektrik potansiyeli
————————————————————————— İyonlar Hücre İçinde (K+) Hücre Dışında (Na+, Cl–) Na K Cl– HCO– Elektrik potansiyeli –90 V ————————————————————————
 Hücre Dışında (Na+, Cl–) Na K Cl– HCO– Elektrik potansiyeli –90 V 0. ————————————————————————")
86
III. Mübadele (değiş-tokuş) difüzyonu
Zarın bir yanındaki iyon, zarın diğer yanında bulunan aynı kimyasal tipteki bir iyonla değiştirilir. Ortamdaki uygun iyonun konsantrasyonu arttıkça iyon değiştirilmesi azalır.

87
IV. Yığın olarak geçiş Maddeler, hidrostatik veya ozmotik kuvvetlerle zarlardan geçer Meselâ, omurgalıların böbreğinde, glomerulusdaki kanın hidrostatik basıncı ozmotik basıncından yüksek olunca, Bowmann kapsülü'nün duvarından fazla kan süzülerek ürik asitle birlikte proksimal dalgalı kanal'a geçer.

88
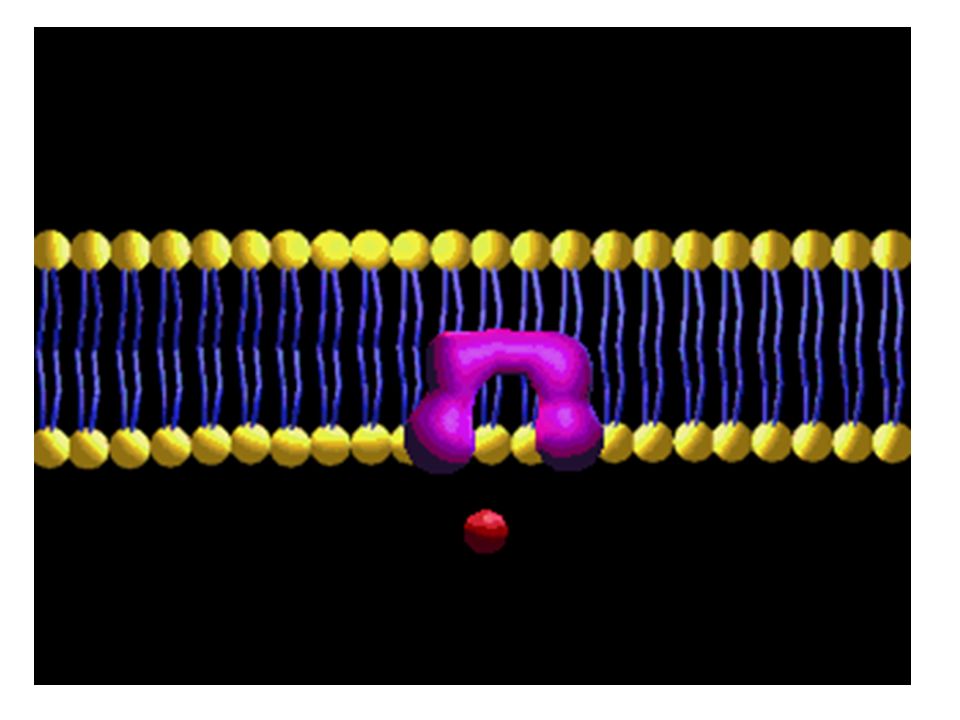
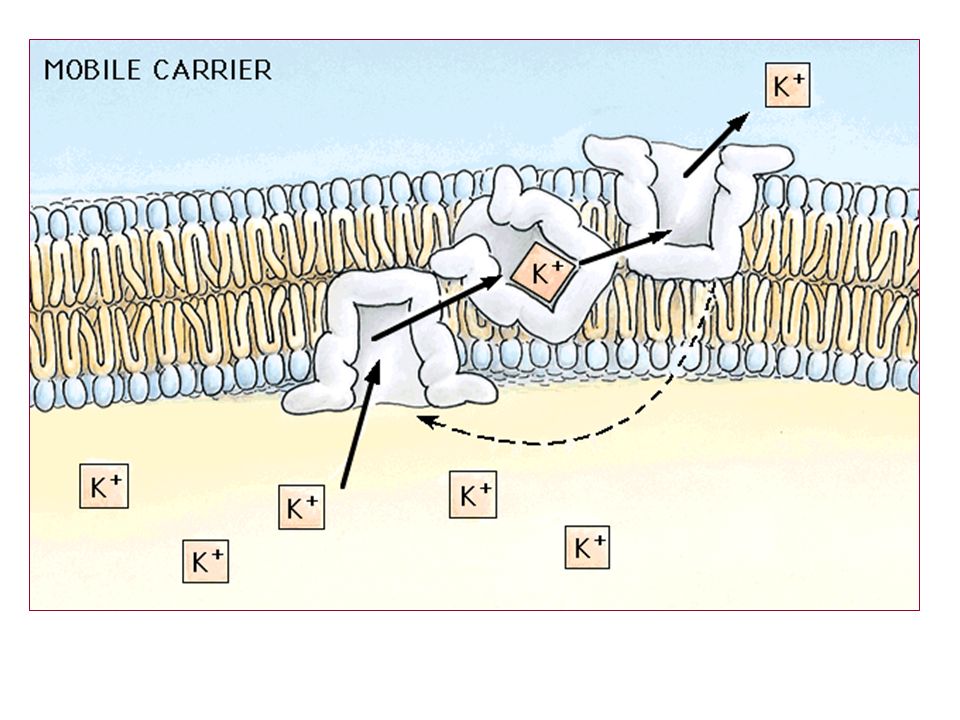
2. Kolaylaştırılmış Difüzyon
Bu tip difüzyonda zarda permeaz olarak bir proteinin bulunması gerekir. Eritrosit zarlarında Band III proteini olarak tanınan integral bir protein bu olay için örnek verilebilir. Şekerlerin ve amino asitlerinin zardan geçişi Proteinin bulunuşu enerji sarfını azaltmaya yarar. Şekerin zar yüzeyine bağlanması proteazda bir konformasyon değişmesine yol açar.

90
Glucose is moved across cell membranes

92
3. Aktif Taşıma Pasif taşımaya zıt olan bir taşıma şeklidir.
Aktif taşıma enerji sarfını gerektirmektedir. Aktif taşıma için gerekli enerji ATP’den sağlanır. Taşıma işi zarda bulunan bir protein ile yapılır. Aktif taşıma sırasında taşınacak madde proteine bağlanır. Zarın iç yüzeyinde proteine yüksek enerji bağlanması ile taşınan madde iç tarafa bırakılır.

93
Bazı proteinler Sodyum/Potasyum pompasında olduğu gibi molekülleri taşırken ATP den enerji kullanırlar. Burada fosfat bağındaki enerji sodyum atomlarının (Kırmızı) potasyumla (Sarı) değiştirilmesinde kullanılır.
 potasyumla (Sarı) değiştirilmesinde kullanılır..")
94
P sınıfı ATPaz'lar: Transmembran polipeptittirler.
ATP enerjisine bağlı olarak yapılan iyon taşınmasında görev yapan üç grup ATPaz vardır. Bunlar P, V ve F sınıfı ATPaz'lardır. P sınıfı ATPaz'lar: Transmembran polipeptittirler. Taşıma sırasında fosforlaşarak aktive olurlar. Yüksek ökaryotlarda Na+ ve K+ Memelilerin asit salgılayan hücrelerinde H+ ve K+ Sarkoplâzmik retikulum ve ökaryot plâzma zarında Ca taşırlar.

95
V sınıfı ATPaz'lar: H+ taşıyan ATPaz'lardır.
Mantar ve bitki vaküol zarında bulunurlar. Hayvan hücrelerinde lizozom, endozom ve salgı kesecikleri zarlarında bulunurlar. Bu enzimler 3-5 farklı polipeptitten meydana gelmiştir. Çalışması sırasında fosforlaşmaya uğrarlar.

96
F sınıfı ATPaz'lar: Birçok bakteride, bütün ökaryot hücrelerin mitokondri iç zarında H+ taşırlar. Taşıma sırasında fosforlaşarak aktive olan bu enzimler 8-13 polipeptitten meydana gelmişlerdir.

97
Aktif taşımanın ilginç bir örneği iyonların pompalanmasıdır.
yıllarında memeli eritrositlerinde Na’un hücre zarından dışarı, K’un ise içeri geçişi için bir enerji gerektiğini, bu enerjinin ATP tarafından verildiğini ve Na+K+ATPaz olarak bilinen bir enzimin (proteaz) görev yaptığını, açıklanmıştır.
 görev yaptığını, açıklanmıştır.")
98
Ayrıca Na+ iyonlarının dışarı çıkarılması elektrik potansiyeli farkına da zıt olan bir olaydır.
Diğer taraftan K+ iyonları, hücrede daha yoğun bulunduğu için, pasif olarak hücreden dışarı çıkmaya zorlanmaktadır. Durum Cl– iyonları için de Na+ iyonlarında olduğu gibidir.

99
+ + + + + + + + + + + - + - - - - - - - - - - -
Na+ Na+ Na+ Cl- Cl- Cl- K+ + + + + + + + + + + + - + - - - - - - - - - - - K+ K+ K+ Na+ Cl- Zarın elektrik potansiyel derecesi ile iyon konsantrasyon derecesi farkına rağmen Na+ iyonlarının hücre dışına pompalanması

100
Bu işlem sodyum pompası olarak bilinir
Bu işlem sodyum pompası olarak bilinir. Sodyum-potasyum ATPaz taşıma proteininin fosforilasyonu ile yapısında meydana gelen konformasyon değişmesi sonucu bu aktif taşıma yapılmaktadır. Enzimin fonksiyonu için Na+ ve K+ iyonları ile beraber Mg+2 iyonlarının da bulunması gerekmektedir. Na+, ancak zarın iç tarafında, K+ iyonları da sadece zarın dışında bulunduğunda pompayı aktive eder ve pompa çalışır. Na+K+ATPaz fonksiyonel bağlanma yerlerine sahip integral bir proteindir.

101
Zara sıkıca bağlı bir protein olan Na+K+ATPaz enzimi
iki alfa ve iki beta alt biriminden oluşmuş bir tetramer yapısındadır. Her bir beta alt birimi molekül ağırlığında olup glikoprotein yapısındadır. Her bir alfa alt birimi ise molekül ağırlığındadır ve karbohidrat kapsamaz.

102
Eritrosit zarlarında bu enzimden sadece molekül bulunduğu ve bir dakikada K+ iyonunu içeri taşıdığı hesaplanmıştır. Her devirde 3 Na+ iyonu dışarı, 2 K+ iyonu içeri taşınır. Enzime Na+ ve K+ iyonlarının bağlanması için enzimde meydana gelen konformasyon değişiklikleri E1 ve E2 olarak gösterilir. Hücrenin iç tarafında, enzime Na+ iyonu bağlanınca enzim fosforile olur. Dış darında K+ iyonu bağlanınca enzim defosorile olur.

103
Şekil. Na+ iyonu pompası.
ATP ADP + 3 Na+ DIŞ E1 Na+ K+ -P Şekil. Na+ iyonu pompası.

104
+2 K+ DIŞ Na+ K+ +2K+ +3Na+ iP - P E1 E2

105
Na+K+ATPaz'ın sinir, beyin ve böbrek hücre zarlarında çok aktif olarak bulunduğu gözlenmiştir.
Hücrede ozmotik dengenin devamlılığı için aktif taşıma temel bir olaydır. Sarkoplâzmik retikulum zarında bulunan Ca+2Mg+2ATPaz enziminin varlığı ve olaya bir fosfolipidin katıldığı gösterilmiştir. Glükoz, diğer monosakkaritler, amino asitleri, fenol kırmızısı, pirimidinler, safra tuzları, penisilin v.s. de hücre zarlarından, konsantrasyon derecelerine zıt olan bir yönde taşınırlar.

106
Taşınmanın yapılabilmesi için, organik maddenin taşınmak üzere zardan alınacağı tarafta, daima Na+ konsantrasyonunun zarın diğer tarafına göre daha yüksek olmasının gerektiği ortaya çıkarılmıştır. Potasyum, lityum, amonyum iyonlarının hücre dışında yüksek oranda bulunması, organik maddelerin hücre içine taşınmasına engel olur. Zarlarda maddelerin taşınması işinde, taşıyıcı adı verilen proteinin zarda hareket etmediği gösterilmiştir. Sıvı-mozayik zar modeli, zarlarda sabit porlar olduğu fikrini desteklemektedir.

107
Suyun Zardan Geçişi Su zarlardan süratle her iki yönde difüzyonla geçer. Bu olay ozmoz’dur. Her iki çözeltinin kapsadıkları su miktarına göre bir hidrostatik basınç'ları vardır. Hidrostatik basınç farkı bir ozmotik basınç meydana getirir.

108
Hücre zarı suya karşı çok permeabldir.
Suyun fazlası hücrelerin içinde ve aralarında dağılır ve hücrede bir hacım artması olmaz. Vücutta NaCl fazla olduğu zaman, Vücut, normal şartlarda, çeşitli kısımlarındaki suyu dengede tutmak için ozmotik basıncı bir düzenleme sistemi olarak kullanır.

109
Eritrositlerin bulunduğu dış çevrede % 0
Eritrositlerin bulunduğu dış çevrede % 0.9 kadar NaCl bulunması hücrelerin canlılığının devamı için gereklidir. Hücreler ancak % 0.9 NaCl'lü, kendi ozmotik basınçlarına denk olan bir ozmotik basınçlı, yani aynı tuz yoğunluğundaki çevrede (izotonik) canlı kalabilmektedir. Memeli hücreleri için % 0.9 tuz yoğunluğu izotonik bir çevre sağlar. Omurgasız hücreleri için bu tuz yoğunluğu kabaca % 0.6 olarak tespit edilmiştir.
 canlı kalabilmektedir. Memeli hücreleri için % 0.9 tuz yoğunluğu izotonik bir çevre sağlar. Omurgasız hücreleri için bu tuz yoğunluğu kabaca % 0.6 olarak tespit edilmiştir.")
110
Hipotonik ortam İzotonik ortam Hipertonik ortam

111
OZMOZ Ozmoz özel bir tip diffüzyondur
Su yarı geçirgen bir zardan geçerek yüksek konsantrasyonlu bir bölgeden düşük konsantrasyonlu bölgeye geçer Çözünen moleküller (İyonlar, organik komponentler vs.) solut olarak adlandırılır Solutlerin çoğu zarı geçemez Solut konsantrasyonu arttıkça serbest su konsantrasyonu azalır OZMOZ Hipotonik solüsyon Hipertonik solüsyon Solut molekülü Seçici geçirgen zar Hipotonik solüsyon Hipertonik solüsyon Su molekülü Seçici Geçirgen Zar Su ve solut molekül kümeleri Suyun geçişi
 solut olarak adlandırılır. Solutlerin çoğu zarı geçemez. Solut konsantrasyonu arttıkça serbest su konsantrasyonu azalır. OZMOZ. Hipotonik. solüsyon. Hipertonik. solüsyon. Solut. molekülü. Seçici. geçirgen zar. Hipotonik solüsyon. Hipertonik solüsyon. Su molekülü. Seçici. Geçirgen. Zar. Su ve solut. molekül kümeleri. Suyun geçişi.")
112
a. Ozmotik basınç deneyinin başlangıcı b. Dengede
Cam tüp Lastik tıkaç Lastik bant Sükroz molekülü Selofan kese Su molekülü a. Ozmotik basınç deneyinin başlangıcı b. Dengede

113
Ayrıntılı deneyler için izotonik çözeltilere çok az miktarda başka tuzların da eklenmesi gerekir. Böylece meydana getirilen özel çözeltiye fizyolojik çözelti denir Çeşitli bilim adamlarının geliştirmiş olduğu özel fizyolojik çözeltiler vardır. Tablo. Ringer çözeltisinin kimyasal karışımı ——————————————————————————— Maddenin adı Gram olarak miktarı NaCl KCl CaCl Na H CO EDTA (Etilen diamin tetra asetik asit) 20 Saf su litre
 20. Saf su 1 litre.")
114
Organizmanın bütün olarak ozmotik basıncının ayarlanması böbreklerle sağlanarak doku sıvısı ile hücre içi sıvısının ozmotik basıncı eşit tutulmaya çalışılır. Bitki hücrelerinde, hücre içi sıvısının ozmotik basıncının hücre dışı sıvısının ozmotik basıncından yüksektir ancak, selüloz duvar hücrelerin patlamasını önler. Özellikle tatlı sularda yaşayan bir hücreli organizmalarda, ozmotik basınç, kontraktil vaküol denen bir organel aracılığı ile düzenlenir.

115
Hücrelerin Birbirini Tanıması ve Hücre Adezyonu
Hücrelerin birbirini tanımasını sağlayan proteinler hücre zarlarında bulunurlar ve zarın proteinlerine bağlı olan karbohidratlar da bu mekanizmaya girerler. Eğer bu tanıtıcı moleküller bulunmazsa, organizmaya giren diğer proteinler v.b. yabancı kabul edilirler ve vücudun antikor savunma sistemi tarafından hücuma uğrar.

116
Antikor sistemi organizmayı korur.
İnsanda günde kadar mutasyon olmaktadır. Bunların çoğu hücre için öldürücüdür. Fakat, mutasyona uğrayan hücre yaşama imkânı bulursa, bu hücreden işleyişi bozuk, kötü huylu (habis) yeni hücrelerin meydana gelmesi mümkün olur.
 yeni hücrelerin meydana gelmesi mümkün olur.")
117
Zarda meydana gelen protein değişmesi antikor tarafından yok edilir
Zarda meydana gelen protein değişmesi antikor tarafından yok edilir. Aksi takdirde kanser çok daha yaygın olurdu. Doku naklinde, yabancı proteinli dokunun uyum sağlaması için antikor sistemini engellemek gerekir. Kan hücrelerinin antigen proteinleri tanıtıcı proteinlerdir. Bunlar kan grubu faktörleri'dir ve siyalik asit ile amino şekerleri kapsayan glikozaminoglikanlardır (mukopolisakkaritler).
.")
118
Rhesus D (Rh) faktörü için kırmızı kan hücre zarı yüzeyinde 3. 000-20
Rhesus D (Rh) faktörü için kırmızı kan hücre zarı yüzeyinde tanıma yeri vardır. A faktörü için ise bunun misli tanıma yeri bulunur. Organizmada bir organda yan yana bulunan komşu hücreler aynı tip veya farklı tip hücrelerdir. Embriyo gelişirken, her organdaki birbirine benzeyen ve benzemeyen hücreler arasında seçici interaksiyonlar olur. Hücre diğer hücrelerin yüzeyini tanıma yeteneğindedir.
 faktörü için kırmızı kan hücre zarı yüzeyinde tanıma yeri vardır. A faktörü için ise bunun misli tanıma yeri bulunur. Organizmada bir organda yan yana bulunan komşu hücreler aynı tip veya farklı tip hücrelerdir. Embriyo gelişirken, her organdaki birbirine benzeyen ve benzemeyen hücreler arasında seçici interaksiyonlar olur. Hücre diğer hücrelerin yüzeyini tanıma yeteneğindedir.")
119
Gerek embriyo ile gerekse süngerlerle yapılan deneyler hücrelerin yapışma özelliği için çok iyi delillerdir. Hücreleri birbirine bağlayan hücre adezyonu mekanizması, muhtemelen iki hücrenin karşılıklı birbirini tanıyan tamamlayıcı molekülleri arasında yüksek çekimli kovalent olmayan bağlar kurulur. Hücre adezyonunda, muhtemelen plâzma zarından glikoproteinlerin dışa çıkan uzantıları birbiri ile tamamlayıcı şekilde interaksiyon yapar.

120
Hücre Adezyon Molekülleri
Hücrenin komşu hücre ile ve hücre dışı ortam ile adezyonu, embriyonun gelişimi sırasında ve erginde hücrenin vücudun bir yerinden başka bir bölgesine göç etmesi sırasında çok önemlidir. Hücre adezyon molekülleri transmembran proteinlerdir. Bu moleküller beş grupta incelenmektedir: a. İmmünoglobulin grubu moleküller, b. Kaderin grubu moleküller, c. İntegrin grubu moleküller. d. Selektin grubu moleküller e. Musin grubu moleküller

121
İmmünoglobulin grubu hücre adezyonu molekülleri
Bu gruptaki moleküller, N-CAM (nöral hücre adezyon molekülleri), miyeline bağlı glikoproteinler, CD tipi antijenler, I-CAM-1 (Interselüler hücre adezyon molekülleri) vb. lerdir. N-CAM, daha çok nöral hücrelerde bulunan glikoprotein yapısındaki moleküllerdir. Embriyoda KD, ergin birey hücrelerinde KD arasındadır. N-CAM molekülleri üç tip olur: Id, Sd ve SSd. Bunlardan Id ve Sd N-CAM molekülleri transmembran glikoprotein yapısındadır. SSd N-CAM molekülleri ise zara fosfolipit inozitol ile bağlıdır.
, miyeline bağlı glikoproteinler, CD tipi antijenler, I-CAM-1 (Interselüler hücre adezyon molekülleri) vb. lerdir. N-CAM, daha çok nöral hücrelerde bulunan glikoprotein yapısındaki moleküllerdir. Embriyoda KD, ergin birey hücrelerinde KD arasındadır. N-CAM molekülleri üç tip olur: Id, Sd ve SSd. Bunlardan Id ve Sd N-CAM molekülleri transmembran glikoprotein yapısındadır. SSd N-CAM molekülleri ise zara fosfolipit inozitol ile bağlıdır.")
122
Her 3 molekülün zar dışında kalan kısımları birbirine benzer ve molekül zinciri üzerinde 5 adet yarı halkasal şekilde yapılara sahiptirler. N-CAM moleküllerinin esas görevlerinden biri diğer hücrenin yüzey moleküllerini tanımaktır. Diğer bir önemli görevi de kas sinir bağlantısını sağlamaktır. Miyeline bağlı glikoproteinler (MAG) miyelinde görülen ve 100 KD molekül ağırlığındaki moleküllerdir. MAG molekülleri yalnız kollagen ve heparinle bağlantı yapabilir.
 miyelinde görülen ve 100 KD molekül ağırlığındaki moleküllerdir. MAG molekülleri yalnız kollagen ve heparinle bağlantı yapabilir.")
123
CD tipi antijenler (CD: Cluster Designation) özellikle T lenfositleri alt populasyonlarında hücre yüzeyi marker'ı olarak bulunurlar. Hedef hücrenin tanınmasında rol oynar

124
I CAM-1 (Intercellular Adhesion Molecule-1) 76-114 KD molekül ağırlığındadır.
I CAM-1 adezyon molekülleri fibroblast, endotelyum ve keratinosit tipi hücrelerde bulunur. Sitotoksik T ve helper T lenfositlerinin görevlerini yapmalarında ve NK (Natural Killer) hücreleri ile granülostlerin antikor aracılığı ile yaptıkları öldürme faaliyetlerinde önemli rolü olmaktadır.
 hücreleri ile granülostlerin antikor aracılığı ile yaptıkları öldürme faaliyetlerinde önemli rolü olmaktadır.")
125
Kaderin grubu hücre adezyonu molekülleri
Bu grupta bulunan moleküller Ca+2' a bağlı hücre-hücre adezyonunda görev yapan proteinlerdir. Hücreler arası adezyonun Ca+2' a bağlı olan veya olmayan şekillerde olabilir. Ca+2' a bağlı adezyon molekülleri de transmembran protein yapısında olur. Kaderin moleküllerinin sitoplâzma içinde kalan kısımları aktin filamentleri ile birleşmiştir. Kaderin grubu moleküller morfogenezde düzenleyici rol oynamaktadır.

126
İntegrin grubu hücre adezyon molekülleri
Bu moleküller de transmembran polipeptitleridir. Hem matriks komponentine hem de sitoplâzmik komponente bağlıdırlar. Böylece hücre dışı ve hücre içi komponentleri birbirine yaklaştırırlar. VLA antijenleri (Very Late Antigens) yine bu gruptan olup uzun ömürlü T lenfositlerinde vardır. VLA 1 den VLA 5 e kadar beş tipi bulunmaktadır.
 yine bu gruptan olup uzun ömürlü T lenfositlerinde vardır. VLA 1 den VLA 5 e kadar beş tipi bulunmaktadır.")
127
Selektin grubu moleküller
Zar glikoproteinlerindendir. Ucunda bulunan lektin benzeri kısımları ile belirli karbohidratları tanır. Bunların L, E ve P olmak üzere üç tipi tanımlanmıştır. L-Selektinler dolaşan lökositlerde, E ve P tipleri de endotel hücrelerinde bulunur. Bunlar lökositlerin endotelden damar dışına çıkmasında (diyapedez) görev yapar.
 görev yapar.")
128
Musin grubu moleküller
Karbohidratça çok zengin, polipeptit zincirinde serin ve treonin amin amino asitleri bol bulunan musin benzeri maddelerdir. Bu moleküller lökositlerdeki selektinler tarafından tanınır.

129
Kanser Hücreleri Kanser, hücrelerin kontrolsüz büyümesi, dağılması ve uygun bulduğu yeni yerlerde yerleşip tekrar çoğalması ile (metastaz) oluşur.
 oluşur.")
130
Temas engellemesi (kontak inhibisyon)
Normal kültür hücreleri genel olarak her 24 saatte bir defa bölünür. Kültür kabında tek bir hücre tabakası oluşana kadar bölünme devam eder ve sonra durur. Hücreler arasında bir işaretle bu düzenlenmektedir. Kanserli hücrelerde mitoz bölünme engellenmez ve çoğalan hücreler yığılır ve birkaç hücre tabakasından olaşan düzensiz kitleler oluşur.

131
Normal Kanserli Normal ve kanserli hücrelerin kültür kabında teşkil ettiği tabakanın şematik durumu. Normal hücreler tek tabakadan, kanserli hücreler birkaç tabakadan oluşur.

132
Kanserli hücrelerin zemine yapışması zayıftır ve hareketlilikleri çok fazladır.
Kanserli hücrelerin yüzey özellikleri değişir. Kanserli hücreler arasında aralıklı temas yerleri kaybolur. Glikoprotein reseptörlerine bağlanan lektinler kullanılarak bu reseptörlerin normal hücrelere göre zarın lipit tabakaları içine daha kolayca nüfuz ettiği görülmektedir.

133
Kanserli hücrelerin şeker alışı artar ve plâzma zarında meydana gelen sızıntılar ile hücre içi enzimler ortama çıkar Kanserli hücreler hücre faktörü denilen molekül ağırlıklı bir proteaz salgılar. Bu enzim plâzminojenler'i etkileyerek plâzmin'e çevirir. Bu enzim kan pıhtılarını eritir ve kanserli hücrenin yüzeyinde bulunan protein gruplarını uzaklaştırır. Çoğunlukla kanserli hücreler normal hücrelerde bulunmayan yeni antijenlere sahip olurlar.

134
Zarla Taşınan Sinyaller
Bir organizmadaki iletişim (komünikasyon) o organizmanın bir hücresinden çıkarılan bir kimyasal maddenin başka bir hücreyle etkileşim (interaksiyon)'u ile sağlanır. Bunları dört grupta toplamak mümkündür. A.Endokrin tipi iletişimde endokrin bezlerin kana salgıladığı kimyasal maddeler daha uzak bir bölgedeki hedef hücre ile etkileşime girerek bir cevap meydana gelmesini sağlar.
 o organizmanın bir hücresinden çıkarılan bir kimyasal maddenin başka bir hücreyle etkileşim (interaksiyon) u ile sağlanır. Bunları dört grupta toplamak mümkündür. A.Endokrin tipi iletişimde endokrin bezlerin kana salgıladığı kimyasal maddeler daha uzak bir bölgedeki hedef hücre ile etkileşime girerek bir cevap meydana gelmesini sağlar.")
135
B. Parakrin tipi iletişimde hücreden salgılanan maddeler sinir hücrelerinde olduğu gibi hemen yakınındaki hücrelerin etkileşimini sağlar. C. Otokrin tipi iletişimde T lenfositlerinin sitokin salgılaması gibi hücrenin salgıladığı madde kendisiyle etkileşime girer. D. Hücre-hücre iletişiminde farklılaşmakta olan embriyonik hücreler arasında görüldüğü gibi hücreler komşu hücrenin salgıladığı maddelerle etkileşim yerine o hücrenin zarında yer alan moleküllerle doğrudan etkileşime girer.

136
Hücrelerin birbirleriyle iletişimini sağlayan kimyasal maddelere sinyal molekülleri veya haberci moleküller denir. Hedef hücrelerin plâzma zarının dış tabakasında bulunan spesifik reseptöre tutunurlar. Bu kimyasal habercilere birinci haberciler denir. Bu moleküller reseptöre bağlandıklarında reseptörün ligandı olurlar ve hücrede bir seri reaksiyonun başlamasına sebep olurlar. Bazı birinci haberciler hücreye kolaylıkla girerek sitoplazmadaki reseptörlere bağlanır ve reaksiyonları başlatırlar.

137
Gaz olarak bulunan sinyal moleküllerine nitrik oksit (NO) ve karbon monoksit (CO) örnek verilebilir.
Atmosferde kirletici olarak bulunan NO düşük konsantrasyonlarda olduğu zaman sinyal görevi yapan önemli bir moleküldür Düz kasın gevşemesinde kaslara bitişik endotel hücrelerine bağlanan asetil kolin endotel hücrelerine NO sentezini uyarmaktadır. NO guanilat siklaz enzimini aktive ederek GTP den cGMP oluşmasını ve bunu takip eden basamakta kasın gevşemesini sağlamış olur. Böylece kan damarları da genişlemiş olur.

138
CO de NO gibi guanilat siklazı uyararak cGMP oluşmasını sağlayan kimyasal bir haberci moleküldür.
Steroit grubuna giren haberci moleküller zarlardan kolaylıkla geçerler.

139
Birinci haberci moleküller arasında yer alan nörotransmitter maddeler nöronlar arasındaki sinaps bölgelerinde veya nöronlar ile kas hücreleri arasında iletişimi sağlayan moleküllerdir. Peptit hormonlar (insülin, glukagon) ve büyüme faktörleri (Epidermal büyüme faktörü, EGF) de birinci haberci moleküller arasında yer alır. Çeşitli lipit molekülleri “Eicosanoid” hücre yüzey reseptörlerine bağlanan sinyal molekülleri olarak steroit tipi moleküllerden farklılık gösterirler.
 ve büyüme faktörleri (Epidermal büyüme faktörü, EGF) de birinci haberci moleküller arasında yer alır. Çeşitli lipit molekülleri Eicosanoid hücre yüzey reseptörlerine bağlanan sinyal molekülleri olarak steroit tipi moleküllerden farklılık gösterirler.")
140
Bitki hormonları bitkinin büyüme ve gelişmesinde görev yapan birinci haberci moleküller arasında sayılabilirler. Bunlar arasında oksinler, absisik asit, giberellinler, sitokininler ve etilen bulunur. Bu hücre dışı haberciler hücre zarındaki reseptöre bağlanarak zarda bir seri fiziksel değişmeye yol açar veya zardaki iyon kanallarının açılmasına sebep olur. Bu değişmelerle yeni bir sinyal meydana gelmekte ve bu sinyal, bilgiyi hücre içindeki komponente ulaştırmaktadır. Bu yeni sinyallere ikinci haberciler denmektedir. (cAMP, Ca+2, inozitolfosfat ve 1,2-diasilgliserol)
")
141
Kanda şeker seviyesinin ayarlanmasında üç hormon görev yapar.
Bunlar böbrek üstü bezinin medulla bölgesinden salınan epinefrin hormonu ile pankreastan salınan glukagon ve insülin hormonlarıdır.

142
Glikojenin parçalanması fosforilaz enzimi ile, sentezi ise glikojen sentetaz enzimi ile katalizlenir. Epinefrin ve glukagon hormonları fosforilazı aktive eder, glikojen sentezini engeller ve glikojen parçalanmasını ilerletir (glikojenoliz). İnsülin glükoz salınmasını uyarır, glikojen sentezini aktive eder, glikojen teşekkülünü ilerletir. Bu işlemlerde ikinci haberci olarak siklik adenozin monofosfat (c-AMP) denilen bir maddenin olaya girdiği görülmüştür.
. İnsülin glükoz salınmasını uyarır, glikojen sentezini aktive eder, glikojen teşekkülünü ilerletir. Bu işlemlerde ikinci haberci olarak siklik adenozin monofosfat (c-AMP) denilen bir maddenin olaya girdiği görülmüştür.")
143
Adenozin trifosfat (ATP)
Adenin Üç fosfat Riboz Adenozin trifosfat (ATP)
")
144
Adenin Riboz O Adenilat siklaz enziminin katalizlemesi ile ATP den c-AMP meydana gelmesi

145
Siklik AMP ile zardan haberin alınmasında üç ayrı integral zar komponenti olaya girer.
Birincisi bir reseptör'dür. İkincisi zarın iç yüzeyinde yer alan ve olayı katalizleyen adenilat siklaz enzimi'dir. Bu katalizleme ile ATP'den cAMP teşekkül etmektedir. Üçüncüsü bir düzenleyici (regülatör) protein'dir. Bu da zarın iç tarafında yer alır. G proteini denen bu düzenleyici protein hem adenilat siklaz molekülü ile hem de çeşitli reseptörlerle reaksiyona girer.
 protein dir. Bu da zarın iç tarafında yer alır. G proteini denen bu düzenleyici protein hem adenilat siklaz molekülü ile hem de çeşitli reseptörlerle reaksiyona girer.")
146
Adenilat siklazın aktive olması için, aynı zamanda, guanozin trifosfat (GTP)'ın bulunması gerekmektedir. GTP regülatör proteine bağlanır.

147
Sinyalin zarı geçmesinden sorumlu komponentlerin şeması
Hormon (Birinci haberci) A D ZAR R GTP MgATP Siklik AMP (İkinci haberci) Sinyalin zarı geçmesinden sorumlu komponentlerin şeması
 A. D. ZAR. R. GTP. MgATP. Siklik AMP. (İkinci haberci) Sinyalin zarı geçmesinden sorumlu komponentlerin şeması.")
148
İkinci haberci olarak c-AMP'nin rolü bütün hücrelerde aynıdır ve çok spesifiktir.
Hücre içinde c-AMP seviyesinin artması protein kinaz denilen diğer bir spesifik enzimi aktive eder. Kinaz bir fosfat molekülünü bir vericiden alıcıya transfer eden bir enzimdir. Verici molekül ATP'dir. Alıcı da diğer bir proteindir. Bu proteinin fosforile olması temel ve tek bir düzenleyici mekanizmadır.

149
Hücrede c-AMP'den başka siklik guanozin monofosfat (cGMP) ve kalsiyum iyonları da ikinci haberci moleküller olarak görev yaparlar. Guanilat siklaz GTP'den c-GMP teşkilini sağlar. Bu enzim de zarın iç tarafında bulunmaktadır.

150
Zarın Ca+2 iyonlarına karşı permeabilitesini geçici olarak yükseltilmek suretiyle bazı maddeler kalsiyumun ikinci haberci olarak görev almasını sağlarlar. Meselâ, sekresyon, birçok hücrede Ca+2 ile uyarılır. Uyarıcı gelince, cevap olarak, hücre, plâzma zarındaki kalsiyum kapıları'nı açar, hücrenin çevresindeki yüksek Ca+2 yoğunluklu çevreden hücre içine süratle Ca+2 girer. Hücreye birden Ca+2 dolması plâzma zarı ile salgı granülünün birleşmesini sağlar. Böylece salgı dışarı çıkar.

Benzer bir sunumlar








 Nedir? Lipitlerdir.>")








>")

