Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Kalp Yetersizliğinde Yeni Ufuklar: “Mikro RNA”
Mehmet Birhan YILMAZ, MD, FESC Cumhuriyet Üniversitesi Tıp Fakültesi Kardiyoloji AD, Sivas

3
Mikro RNA=miRNA İlk kez 1993 yılında keşfedildi.
İsimlendirilmesi 2001 yılında oldu. Keşfedilmesi son 10 yılın en önemli buluşlarından biri olarak kabul ediliyor

4
Mikro RNA (miRNA) 20-25 nükleotid uzunlukta tek sarmal RNA molekülleridir. Tüm ökaryotik hücrelerde bulunurlar. miRNA’lar, DNA’dan transkribe edilen genler tarafından (intron bölgesi) kodlanır. Proteine translasyon olmaz ve bu yüzden “non-coding RNA” parçacığı olarak bilinirler.
 kodlanır. Proteine translasyon olmaz ve bu yüzden non-coding RNA parçacığı olarak bilinirler.")
5
Genlerin protein ürünlere dönüşmesini düzenledikleri fark edilmiştir
Genlerin protein ürünlere dönüşmesini düzenledikleri fark edilmiştir. mRNA (mesajcı RNA)’ların bir bölgesine benzer nükleotid dizilimleri içerir. Bu sayede mRNA’ların protein ürün oluşturmasını engeller, hatta mRNA’ya tamamlayıcı zincirler içerdiği için mRNA’nın yıkımını hızlandırabilirler Nadiren “histon modifikasyonu” (DNA’yı paketleyen yapı)ve DNA metilasyonu üzerinden de etki göstererek hedef genlerin üretimini düzenler.
’ların bir bölgesine benzer nükleotid dizilimleri içerir. Bu sayede mRNA’ların protein ürün oluşturmasını engeller, hatta mRNA’ya tamamlayıcı zincirler içerdiği için mRNA’nın yıkımını hızlandırabilirler. Nadiren histon modifikasyonu (DNA’yı paketleyen yapı)ve DNA metilasyonu üzerinden de etki göstererek hedef genlerin üretimini düzenler.")
6
mRNA’nın tersine (çok kararsız bir molekül) endojen ribonükleaz aktivitesine karşı oldukça dayanıklı ve kararlı moleküllerdir dolayısıyla sağlıklı ve hastalıklı durumlarda düzeyleri kolaylıkla tespit edilebilir. Topkara VK et al. Cardiovasc Drugs Ther 2011

7
miRISC: RNA induced silencing complex
Exportin: nucleo sytoplasmic shuttle

8
İnsanda muhtemelen 1000’den fazla miRNA var
Bu genler tüm protein kodlayan genlerin yaklaşık 1/3’ünün fonksiyonunu düzenler. Bazı insan miRNA’ları RNA polimeraz III ile transkribe olur. Her biri farklı genomik organizasyona ve farklı biyogenetik mekanizmalara sahip çok sayıda miRNA altsınıfı var. Guarnieri and Dileone Ann.Med. 40:

9
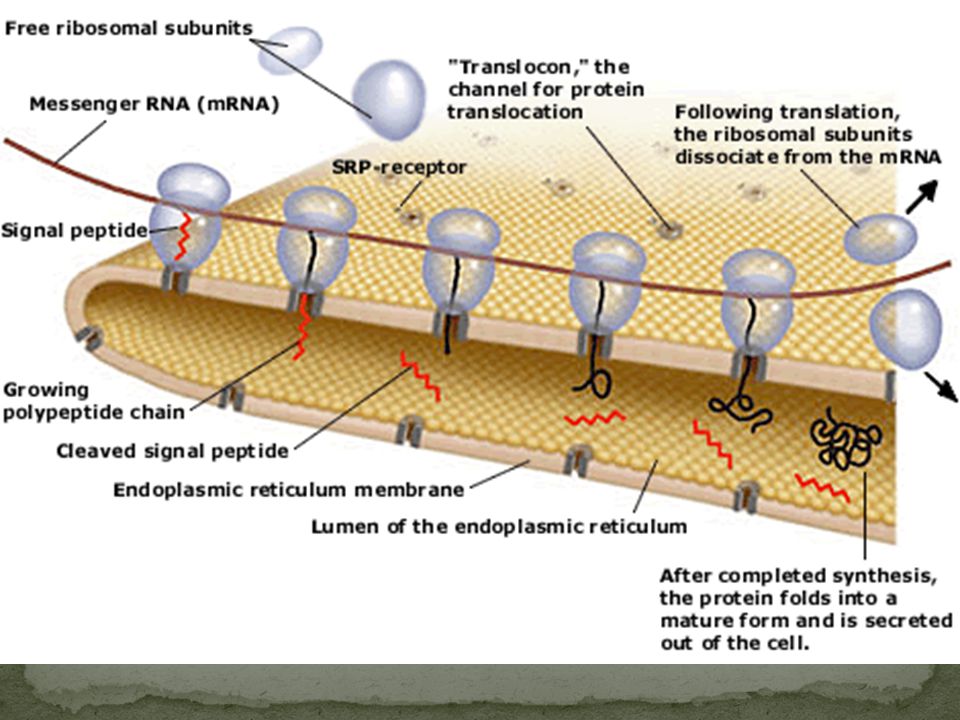
SRP (Sinyal Tanıyan Partikül): bir ribonukleoproteindir (protein-RNA)
SRP (Sinyal Tanıyan Partikül): bir ribonukleoproteindir (protein-RNA). Spesifik proteinleri ökaryotlarda ER’a, prokaryotlarda plazma membranına taşır. Ökaryotlarda sitosolde bulunur. ER’daki sinyal diziye (peptide) bağlanır ve sentezlenen proteini ribozom boyunca ER içine bırakır.
: bir ribonukleoproteindir (protein-RNA). Spesifik proteinleri ökaryotlarda ER’a, prokaryotlarda plazma membranına taşır. Ökaryotlarda sitosolde bulunur. ER’daki sinyal diziye (peptide) bağlanır ve sentezlenen proteini ribozom boyunca ER içine bırakır.")
12
miRNA’ların tespiti Küçük RNA klonlaması Northern blots Microarray
QPCR Websites:

13
Eric M. Small & Eric N. Olson; Nature 2011
miRNA işlevi 1 2 3 i): bir miRNA pek çok mRNA hedefine sahip olabilir ii): birden fazla miRNA bir arada çalışıp bir fonksiyonu düzenleyebilir. iii): miRNAs bazı aktivatör ve inhibitörler üzerine etkiyebilir (indirekt etki). Eric M. Small & Eric N. Olson; Nature 2011
: bir miRNA pek çok mRNA hedefine sahip olabilir. ii): birden fazla miRNA bir arada çalışıp bir fonksiyonu düzenleyebilir. iii): miRNAs bazı aktivatör ve inhibitörler üzerine etkiyebilir (indirekt etki). Eric M. Small & Eric N. Olson; Nature")
14
Hayvan Çalışmalarında miRNA Fonksiyonları
miRNA’lar dokuya spesifik ekspresyon paternleri gösterir. Örneğin, miR-1 kalp, miR-122 karaciğer, miR-124a beyinde bulunur. 1. miRNA gelişimsel zamanlamayı (embriyogenez) düzenler: 2. miRNA gelişimsel süreçte önemli: miRNA’lar kök hücreleri, embriyo, beyin, kalp, KC de dahil olmak üzere tüm hayvan dokularının normal gelişiminden sorumludur. Spesifik miRNA’lar spesifik gelişimsel süreçte spesifik dokuların gelişimini kontrol eder. Örneğin, Dicer mutantı zebrafish embriyoları başlangıçta normal olarak gelişir ancak fertilizasyondan yaklaşık 8 gün sonra arrest olur. HOX (hayvan gelişiminde önemli bir gen) miR-196 tarafından negatif olarak regüle edilir. Bu miRNA’nın eksik ekspresyonu anormal HOX geni ekspresyonuna neden olur ve gelişimsel anomaliler ortaya çıkar. Drosophila’da miR-1 mutasyonları kardiyogenez ve kas gelişiminde defektler oluşturabilir.
 düzenler: 2. miRNA gelişimsel süreçte önemli: miRNA’lar kök hücreleri, embriyo, beyin, kalp, KC de dahil olmak üzere tüm hayvan dokularının normal gelişiminden sorumludur. Spesifik miRNA’lar spesifik gelişimsel süreçte spesifik dokuların gelişimini kontrol eder. Örneğin, Dicer mutantı zebrafish embriyoları başlangıçta normal olarak gelişir ancak fertilizasyondan yaklaşık 8 gün sonra arrest olur. HOX (hayvan gelişiminde önemli bir gen) miR-196 tarafından negatif olarak regüle edilir. Bu miRNA’nın eksik ekspresyonu anormal HOX geni ekspresyonuna neden olur ve gelişimsel anomaliler ortaya çıkar. Drosophila’da miR-1 mutasyonları kardiyogenez ve kas gelişiminde defektler oluşturabilir.")
15
3. miRNA metabolizmada önemli:
miR-155 immun yanıt ve lenfosit fonksiyonunda önemli. 4. miRNA hücresel yaşamda önemli: miR-14 de sineklerde hücre ölümünü baskılayıcı bir etkisi var. miR-375 beta hücrelerden insülin salgısını düzenliyor.

16
miRNA ve kanser İki önemli bulgu:
miRNA’ların büyük çoğunluğu kanser ile ilişkili genomik bölgelerde yerleşmiştir (1). miRNA ekspresyonları kanser hücrelerinde ve ilişkili normal doku hücrelerinde farklıdır (2). Neredeyse tüm kanserlerde miRNA’ların anormal ekspresyonları görülür: B cell kronik lenfositik lösemide mir-15a ve mir-16-1 silinmiştir (3). İnsan AC kanserlerinde let-7 downregulasyonu var. let-7’nin fazla ekspresyonu AC kanserini inhibe ediyor . 1. Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci U S A Mar 2;101(9): 2. Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA, Downing JR, Jacks T, Horvitz HR, Golub TR. MicroRNA expression profiles classify human cancers. Nature Jun 9;435(7043):834-8. 3. Guarnieri and Dileone MicroRNAs: A new class of generegulators. Ann.Med. 40:
. miRNA ekspresyonları kanser hücrelerinde ve ilişkili normal doku hücrelerinde farklıdır (2). Neredeyse tüm kanserlerde miRNA’ların anormal ekspresyonları görülür: B cell kronik lenfositik lösemide mir-15a ve mir-16-1 silinmiştir (3). İnsan AC kanserlerinde let-7 downregulasyonu var. let-7’nin fazla ekspresyonu AC kanserini inhibe ediyor . 1. Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci U S A Mar 2;101(9): Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA, Downing JR, Jacks T, Horvitz HR, Golub TR. MicroRNA expression profiles classify human cancers. Nature Jun 9;435(7043): Guarnieri and Dileone MicroRNAs: A new class of generegulators. Ann.Med. 40:")
17
SiRNA= small interfering RNA:
20-25 nükleotid uzunlukta çift sarmal yapıda (miRNA tek sarmal) RNA molekülleridir. “RNA interferans” yolağının (gen ekspresyonunu baskılama yolağı) temel ayağını oluşturur.
 RNA molekülleridir. RNA interferans yolağının (gen ekspresyonunu baskılama yolağı) temel ayağını oluşturur.")
18
miRNA ile siRNA arasındaki farklılık nedir?
Her iki tür de gen ekspresyonunun düzenlenmesinde rol alır. siRNA dsRNA’dan orijinlenir. siRNA büyük ölçüde yabancı RNA’ya (genellikle viral) etki gösterir ve hedefine %100 olarak bağlanır. miRNA saç tokası şeklinde ssRNA’dan orijinlenir, endojen kaynaklı miRNA, posttranskripsiyonel gen ekspresyonunu düzenler, %100’lük bir komplementerlik sağlamaz (biraz mismatch) miRNA, mRNA’yı ortadan kaldırmak yerine mRNA’nın translasyonunu modüle eder.
 etki gösterir ve hedefine %100 olarak bağlanır. miRNA saç tokası şeklinde ssRNA’dan orijinlenir, endojen kaynaklı. miRNA, posttranskripsiyonel gen ekspresyonunu düzenler, %100’lük bir komplementerlik sağlamaz (biraz mismatch) miRNA, mRNA’yı ortadan kaldırmak yerine mRNA’nın translasyonunu modüle eder.")
19
Tanımlanmış KV sistem miRNA’ları

20
MicroRNA-Kalp Yetersizliği
miRNA function during heart disease. Cardiac remodeling involves numerous disease processes. It is becoming increasingly clear that miRNAs fulfill specific functions during this process. Whereas miR-21, -133, -150, -195, and -214 influence cardiomyocyte hypertrophy, miR-1 and -133 have additionally been implicated in cardiac arrhythmogenesis. In addition, miR-21 and miR-195 also appear to influence apoptosis. miR-208 influences cardiac contractility by regulating myosin content and, by so doing, secondarily influences hypertrophy and fibrosis. The process of cardiac fibrosis is regulated by miR-21 and -29, both of which are highly expressed in cardiac fibroblasts. Neoangiogenesis post-MI relies on miR-126. Van Rooij ve arkadaşları TAC calcineurin A-mediated hypertrophy hayvan modellerinde 28 miRNA tanımlamışlardır ve bunların çoğunun düzeyi yetersizlikteki kalpte artmaktadır. (Van Rooij and al; 2006) Thum et al. KY’li hastaların miRNA profillerini incelemiş ve yüksek oranda fetal patern gözlemiştir. (Thum and al; 2007) Van Rooij & al, Circ Res 2008 20
 Thum et al. KY’li hastaların miRNA profillerini incelemiş ve yüksek oranda fetal patern gözlemiştir. (Thum and al; 2007) Van Rooij & al, Circ Res")
21
miRNAs and heart failure
All human data on miRNA function in heart disease to date have come from heart failure patients.19,22,23 Heart failure is defined as the inability of the heart to pump sufficient blood to the organism and is a frequent and fatal outcome of hypertrophy developed under pathological circumstances. For obvious reasons, it is problematic to obtain human cardiac tissue during the hypertrophic phase, before the onset of heart failure. However, our own data and that of others have indicated that there is at least a partial overlap between the miRNAs regulated during hypertrophy and heart failure, and the miRNA expression pattern seems to dictate the disease state.19 Northern blot analysis on both nonfailing and failing human samples indicated that several miRNAs are regulated in a comparable manner as in hypertrophic mouse models.23 In addition to our data, Ikeda et al19 compared miRNA expression in 3 different types of human heart disease (ischemic cardiomyopathy, dilated cardiomyopathy, and aortic stenosis) with normal heart. Among the 87 miRNAs detected in the heart, roughly half were differentially expressed in at least 1 disease group, whereas 7 miRNAs were regulated in the same direction in all 3 disease states.19 Although several studies already indicated miRNA expression to be regulated in human heart disease,22,23,25,37 this study is the first to show commonalities in expression between distinct disease etiologies. These divergent miRNA expression patterns point to miRNAs as biomarkers for subtle phenotypic differences and disease progression and imply that they are active participants in the disease processes. As during hypertrophy, a hallmark of heart failure is the reexpression of a fetal gene program. A study by Thum et al revealed similarities in the mRNA expression patterns of failing and fetal human heart, in that 353 mRNAs were found to be upregulated 2-fold in common in these 2 situations with respect to normal adult heart tissue.22 In addition, microarray analysis indicated that a large fraction of the miRNAs tested were either up- or downregulated in the same direction in fetal and failing heart with respect to normal adult cardiac tissue; the mRNAs that were upregulated in failing heart contained binding sites mainly for the downregulated miRNAs and vice versa.22 Thus, a shift toward fetal gene expression seems to be partly attributable to a change in cardiac miRNA levels occurring with hypertrophy and failure.
 with normal heart. Among the 87 miRNAs detected in the heart, roughly half were differentially expressed in at least 1 disease group, whereas 7 miRNAs were regulated in the same direction in all 3 disease states.19 Although several studies already indicated miRNA expression to be regulated in human heart disease,22,23,25,37 this study is the first to show commonalities in expression between distinct disease etiologies. These divergent miRNA expression patterns point to miRNAs as biomarkers for subtle phenotypic differences and disease progression and imply that they are active participants in the disease processes. As during hypertrophy, a hallmark of heart failure is the reexpression of a fetal gene program. A study by Thum et al revealed similarities in the mRNA expression patterns of failing and fetal human heart, in that 353 mRNAs were found to be upregulated 2-fold in common in these 2 situations with respect to normal adult heart tissue.22 In addition, microarray analysis indicated that a large fraction of the miRNAs tested were either up- or downregulated in the same direction in fetal and failing heart with respect to normal adult cardiac tissue; the mRNAs that were upregulated in failing heart contained binding sites mainly for the downregulated miRNAs and vice versa.22 Thus, a shift toward fetal gene expression seems to be partly attributable to a change in cardiac miRNA levels occurring with hypertrophy and failure.")
22
miRNA ve KY Kardiyak gen ekspresyonundaki değişimler
Fetal gen programları ile yetersizlikteki kalpteki genetik olaylar arasında çok ciddi benzerlikler bulunmaktadır. Bu gen ekspresyonundaki benzerlik %85.5 oranda aynı yöndedir.

23
miRNAhem hipertrofi hem de fibroziste önemli rol oynar
A transgenic approach revealed that myocardial overexpression of miR-195 in mice causes an increase in left ventricular wall size, upregulation of ANP, BNP and βMHC and reduced cardiac output (van Rooij and al; 2006) Cares et al. Focused their studies on miR-1 and miR-133 which had previously been shown to have key roles in skeletal myoblast differentiation and proliferation (Chen J.F and al;2006). They utilized 3 separate models of hypertrophy: TAC, transgenic expression of constitutively active Akt and exercise. In all 3 models both miR-1 and miR-133 were downregulated, a finding which was recapitulated in hearts from patients with DCM (Carè et al; 2007). Elegant work from the Olson lab demonstrated that in mice, co-ordinate regulation of α- and β-MyHC is controlled by miR-208 located in intron 27 of the α-MHC gene (Van Rooij; 2007). When these mice were subjected to TAC there was little sign of increased myocyte size or fibrosis. Moreover the miR-208-deficient mice failed to upregulate β-MHC but instead increased α-MHC expression to compensate. In contrast, ANP and BNP were upregulated as in WT mice, demonstrating that miR-208 is needed for β-MHC upregulation and cardiac remodelling but not for induction of other stress response factors. Intriguingly, miR-208’s role in hypertrophy may involve silencing of thyroid receptor-associated protein 1 (THRAP1) (van Rooij;2007). This silencing of THRAP1 modulates the activity of thyroid hormone and in turn expression of β-MHC. The adult heart responds to injury or hemodynamic overload by activating a variety of intracellular signaling pathways and transcriptional mediators that promote myocyte hypertrophy, reexpression of an embryonic gene program, and remodeling of the extracellular matrix. These events lead to left ventricular dilation and progressive myocardial fibrosis that can culminate in cardiac arrhythmias and failure.17 Several recent studies performed microarray analyses to determine whether miRNAs are dysregulated in hypertrophic and failing hearts.18–23 These studies have revealed signature patterns of miRNAs that are up- and downregulated during pathological cardiac remodeling in rodents and humans. Although miR-1, miR-29, miR-30, miR-133, and miR-150 have often been found to be downregulated, miR-21, miR-23a, miR-125, miR-195, miR-199, and miR-214 are upregulated with hypertrophy. Gain- and loss-of-function studies have implicated these miRNAs in the different aspects of the remodeling process during the progression of heart disease. The continuous discovery of new miRNAs suggests that the present collection of miRNAs implicated in cardiovascular disease is likely to be incomplete. Mechanism for miRNAmediated fibrosis in response to cardiac stress. Cardiac fibrosis is the result of both an increase in fibroblast proliferation and extracellular matrix deposition. Stress leads to upregulation of miR-21 and downregulation of miR-29 and miR-30 in cardiac fibroblasts, as well as downregulation of miR-30 and miR-133 in cardiomyocytes. miR-21 represses Spry1, a negative regulator of ERK-MAP kinase signaling, thereby leading to fibroblast proliferation and fibrosis. miR-29 represses expression of collagens, and miR-30 and -133 repress expression of CTGF, a positive regulator Van Rooij & Olson, J Clin Invest 2007
 Cares et al. Focused their studies on miR-1 and miR-133 which had previously been shown to have key roles in skeletal myoblast differentiation and proliferation (Chen J.F and al;2006). They utilized 3 separate models of hypertrophy: TAC, transgenic expression of constitutively active Akt and exercise. In all 3 models both miR-1 and miR-133 were downregulated, a finding which was recapitulated in hearts from patients with DCM (Carè et al; 2007). Elegant work from the Olson lab demonstrated that in mice, co-ordinate regulation of α- and β-MyHC is controlled by miR-208 located in intron 27 of the α-MHC gene (Van Rooij; 2007). When these mice were subjected to TAC there was little sign of increased myocyte size or fibrosis. Moreover the miR-208-deficient mice failed to upregulate β-MHC but instead increased α-MHC expression to compensate. In contrast, ANP and BNP were upregulated as in WT mice, demonstrating that miR-208 is needed for β-MHC upregulation and cardiac remodelling but not for induction of other stress response factors. Intriguingly, miR-208’s role in hypertrophy may involve silencing of thyroid receptor-associated protein 1 (THRAP1) (van Rooij;2007). This silencing of THRAP1 modulates the activity of thyroid hormone and in turn expression of β-MHC. The adult heart responds to injury or hemodynamic overload by activating a variety of intracellular signaling pathways and transcriptional mediators that promote myocyte hypertrophy, reexpression of an embryonic gene program, and remodeling. of the extracellular matrix. These events lead to left ventricular dilation and progressive myocardial fibrosis that can. culminate in cardiac arrhythmias and failure.17 Several recent studies performed microarray analyses to determine whether. miRNAs are dysregulated in hypertrophic and failing hearts.18–23 These studies have revealed signature patterns of. miRNAs that are up- and downregulated during pathological cardiac remodeling in rodents and humans. Although miR-1, miR-29, miR-30, miR-133, and miR-150 have often been found to be downregulated, miR-21, miR-23a, miR-125, miR-195, miR-199, and miR-214 are upregulated with hypertrophy. Gain- and loss-of-function studies have implicated these miRNAs in the different aspects of the remodeling process during the progression of heart disease. The continuous discovery of new miRNAs suggests that the present collection of miRNAs implicated in cardiovascular disease is likely to be incomplete. Mechanism for miRNAmediated. fibrosis in response to cardiac. stress. Cardiac fibrosis is the result of. both an increase in fibroblast proliferation. and extracellular matrix deposition. Stress leads to upregulation of miR-21. and downregulation of miR-29 and. miR-30 in cardiac fibroblasts, as well as. downregulation of miR-30 and miR-133. in cardiomyocytes. miR-21 represses. Spry1, a negative regulator of ERK-MAP. kinase signaling, thereby leading to. fibroblast proliferation and fibrosis. miR-29 represses expression of collagens, and miR-30 and -133 repress. expression of CTGF, a positive regulator. Van Rooij & Olson, J Clin Invest")
24
miR-208 We discovered that miR-208, a miRNA encoded within intron 27 of the MHC gene, plays a key role in the expression of MHC in response to cardiac stress.47 Although the expression level of miR-208 remains stable during cardiac stress, this miRNA appears to fulfill a dominant function in regulating cardiac hypertrophy and remodeling. In response to pressure overload by thoracic aortic constriction or signaling by calcineurin (a calcium, calmodulin-dependent phosphatase that drives pathological remodeling of the heart), miR-208–null mice showed virtually no hypertrophy of cardiomyocytes or fibrosis and were unable to upregulate MHC expression. In contrast, other stress responsive genes, such as those encoding ANF and B-type natriuretic peptide, were strongly induced in miR-208 mutant animals, demonstrating that miR-208 is dedicated specifically to the control of MHC expression, which can be uncoupled from other facets of the cardiac stress response. These data indicate that miR-208 regulates cardiac remodeling, at least in part, by regulating the stress-induced increase in MHC expression. Because miR-208 expression does not change in response to stress, its requirement for stress-dependent cardiac remodeling suggests that it cooperates with stress signaling to reprogram cardiac gene expression. Because even a subtle shift toward MHC reduces mechanical performance and efficiency of the adult heart,44–46,48 it might be of therapeutic value to exploit miR-208 regulation to prevent an increase in MHC expression during cardiac disease. The cardiac specificity, absence of overt maladaptive effects in the miR-208 mutant animals, and dedication of miR-208 to the cardiac stress response, but not to normal cardiac development, make miR-208 (and its downstream effectors) an attractive therapeutic Figure 8. Control of Myosin and Fast Versus Slow Muscle Gene Expression by the MyomiR Network In the adult heart, miR-208a encoded by the a-MHC gene is required for expression of b-MHC and Myh7b, which encode miR-208b and miR-499, respectively. Activation of Myh7b by miR-208a is constitutive, whereas activation of b-MHC also requires stress signals or absence of thyroid hormone. In slow skeletal muscle, Myh7b and b-MHC are regulated independently of miR-208a. miR-208a, miR-208b, and miR-499 repress the expression of a common set of transcriptional repressors that repress slow myosin and the slow myofiber gene program at the expense of fast muscle gene expression. Activation of the slow myofiber gene program also creates a positive feedback loop via the expression of miR-208b and miR-499, which further reinforce slow muscle gene program. Regulatory interactions within the blue box are shared by cardiac and slow skeletal muscle. miR-208, MHC genininin 27.intron bölgesinde kodlanmıştır. kardiyomiyosit hipertrofisi için gerekli olan ve yüklenmeye yanıt olarak salınımı düzenlenen βMHC ekspresyonunda gereklidir miRNA-208-eksik farelerde kardiyomiyositlerde hipertrofi oluşturulamaz. mekanik performansı ve erişkin kalbinin verimliliğini bozduğu için MHC’yi düzenleyen miRNA-208 düzenleyici (baskılayıcı) tedavi düşünülebilir.
, miR-208–null mice showed virtually no hypertrophy of. cardiomyocytes or fibrosis and were unable to upregulate MHC expression. In contrast, other stress responsive genes, such as those encoding ANF and B-type natriuretic peptide, were strongly induced in miR-208 mutant animals, demonstrating. that miR-208 is dedicated specifically to the control of MHC expression, which can be uncoupled from other facets of the cardiac stress response. These data indicate that. miR-208 regulates cardiac remodeling, at least in part, by regulating the stress-induced increase in MHC expression. Because miR-208 expression does not change in response to stress, its requirement for stress-dependent cardiac remodeling. suggests that it cooperates with stress signaling to reprogram cardiac gene expression. Because even a subtle shift toward MHC reduces mechanical performance and. efficiency of the adult heart,44–46,48 it might be of therapeutic value to exploit miR-208 regulation to prevent an increase in MHC expression during cardiac disease. The cardiac specificity, absence of overt maladaptive effects in the miR-208. mutant animals, and dedication of miR-208 to the cardiac stress response, but not to normal cardiac development, make miR-208 (and its downstream effectors) an attractive therapeutic. Figure 8. Control of Myosin and Fast Versus Slow Muscle Gene. Expression by the MyomiR Network. In the adult heart, miR-208a encoded by the a-MHC gene is required for. expression of b-MHC and Myh7b, which encode miR-208b and miR-499, respectively. Activation of Myh7b by miR-208a is constitutive, whereas activation. of b-MHC also requires stress signals or absence of thyroid hormone. In slow skeletal muscle, Myh7b and b-MHC are regulated independently of. miR-208a. miR-208a, miR-208b, and miR-499 repress the expression of a. common set of transcriptional repressors that repress slow myosin and the. slow myofiber gene program at the expense of fast muscle gene expression. Activation of the slow myofiber gene program also creates a positive feedback. loop via the expression of miR-208b and miR-499, which further reinforce slow. muscle gene program. Regulatory interactions within the blue box are shared. by cardiac and slow skeletal muscle. miR-208, MHC genininin 27.intron bölgesinde kodlanmıştır. kardiyomiyosit hipertrofisi için gerekli olan ve yüklenmeye yanıt olarak salınımı düzenlenen βMHC ekspresyonunda gereklidir. miRNA-208-eksik farelerde kardiyomiyositlerde hipertrofi oluşturulamaz. mekanik performansı ve erişkin kalbinin verimliliğini bozduğu için MHC’yi düzenleyen miRNA-208 düzenleyici (baskılayıcı) tedavi düşünülebilir.")
25
miR-208

26
miR-29 miR-29 kardiyak fibrozisin düzenleyicisidir.
Özellikle infarkt bölgesi ile sağlam bölge arasındaki sınır bölgede down-regüle olur ve kardiyak fibrozis indüklenir. Terapötik up-regülasyonu kardiyak fibrozisi baskılayabilir. MI also induces severe cardiac fibrosis in the infarcted area and hypertrophy and remodeling in the remote myocardium. Profiling of miRNA expression levels in the border zone of the infarct and remote myocardium both 3 days and 2 weeks post-MI revealed a striking miRNA expression pattern.34 Among the regulated miRNAs, the miR-29 family is dramatically downregulated in the border zone flanking the infarcted area. Our data indicate this downregulation of miR-29 to be responsible for the induction of collagens and additional extracellular matrix genes and, thereby, actively contributes to cardiac fibrosis in response to MI. miR-29 is also downregulated in disease models for cardiac hypertrophy and failure.23 These data imply that therapeutic upregulation of miR-29 in response to an ischemic event or cardiac stress might prevent the onset of cardiac fibrosis and thereby maintain cardiac function. A model for the role of miR-29 in cardiac fibrosis. In response to cardiac stress, TGF is activated and triggers the down-regulation of miR-29 in cardiac fibroblasts and consequent up-regulation of the expression of collagens and other ECM proteins involved in fibrosis. At the same time, stress induces the expression of anti-fibrotic BNP, which is secreted by cardiomyocytes and counteracts the activation and thereby profibrotic function of TGF.

27
Eric M. Small & Eric N. Olson; Nature 2011
miR-1, miR-133: Kasa özgü mikroRNAlar. Kardiyak hipertrofinin negatif düzenleyicileri; down-regülasyonu kardiyak hipertrofiye yol açarken, aşırı üretimi, endotelinle uyarılmış hipertrofik yanıtı baskılıyor. miR-21: (Kardiyak fibroblast havuzunun büyümesinden sorumlu olabilir) miR , beraber düzenlenirler ve hipertrofik yanıttan sorumludurlar. Özellikle miR-24, kardiyak gelişim için esansiyeldir. miR-21: KY’de up-regüle olur ve anti-apoptotik role sahiptir (strese maruz kalmış miyositlerin yaşama tutunmasını sağlama rolü) miR-1 pro-apopottik, miR-133 anti-apoptotik mikroRNA’lardır. miR-199a’nın down-regülasyonu hipoksik miyosit ölümünde çok önemlidir. miR-320, iskemi-reperfüzyon hasarında önemlidir. Aşırı üretilmesi hücre ölümüne yol açarken, baskılanması hücre korunumu sağlar. miR29 ailesi ve miR-25’in düzeylerinin artışı (transfeksiyon dahil) primer kardiyak fibroblastlarda kollajen gen ekspresyonunu arttırır. miR-21, KY’deki fibrozisin temel düzenleyicilerindendir (kardiyak fibroblastik hücrelerde bulunur) ve düzeyleri KY’de artar. Düzeyi, AMI sonrası arada kalan bölgede (borderline zone) de artar. miR-133a, kalpteki temel anti-fibrotik mikroRNA’dır. miR-208a, basınç yüküne karşı geliştirilen kardiyak fibrozis için gerekli moleküldür. Topkara V et al. Cardiovasc Drugs Ther 2011. Eric M. Small & Eric N. Olson; Nature 2011
 miR , beraber düzenlenirler ve hipertrofik yanıttan sorumludurlar. Özellikle miR-24, kardiyak gelişim için esansiyeldir. miR-21: KY’de up-regüle olur ve anti-apoptotik role sahiptir (strese maruz kalmış miyositlerin yaşama tutunmasını sağlama rolü) miR-1 pro-apopottik, miR-133 anti-apoptotik mikroRNA’lardır. miR-199a’nın down-regülasyonu hipoksik miyosit ölümünde çok önemlidir. miR-320, iskemi-reperfüzyon hasarında önemlidir. Aşırı üretilmesi hücre ölümüne yol açarken, baskılanması hücre korunumu sağlar. miR29 ailesi ve miR-25’in düzeylerinin artışı (transfeksiyon dahil) primer kardiyak fibroblastlarda kollajen gen ekspresyonunu arttırır. miR-21, KY’deki fibrozisin temel düzenleyicilerindendir (kardiyak fibroblastik hücrelerde bulunur) ve düzeyleri KY’de artar. Düzeyi, AMI sonrası arada kalan bölgede (borderline zone) de artar. miR-133a, kalpteki temel anti-fibrotik mikroRNA’dır. miR-208a, basınç yüküne karşı geliştirilen kardiyak fibrozis için gerekli moleküldür. Topkara V et al. Cardiovasc Drugs Ther Eric M. Small & Eric N. Olson; Nature")
28
Miyosit Kasılması ile İlgili miR
KY’de, α-MHC’nin ATP’yi daha yavaş hidrolize eden (enerj, verimliliği düşük) β-MHC’ye (fetal) dönüştüğü bilinmektedir. Sağlıklı kalpte MHC mikroRNA havuzunun %33’ünü α- MHC oluştururken, KY’de bu oran %2’ye düşmektedir. miR-208, birbirinden 3 nükleotid farkı olan iki form halinde bulunur (miR-208a ve b) miR-208a, α-MHC geninin intron bölgesinde kodlanmıştır. Mir-208, β-MHC gen ekspresyonunun kuvvetli pozitif bir düzenleyicisidir. miR-208a’nın uzun süreli genomik deplesyonu kontraktilitede azalmaya ve AF’ye yol açar (fare modeli)* *Callis TE, J Clin Invest. 2009;119:2772–86.
 β-MHC’ye (fetal) dönüştüğü bilinmektedir. Sağlıklı kalpte MHC mikroRNA havuzunun %33’ünü α- MHC oluştururken, KY’de bu oran %2’ye düşmektedir. miR-208, birbirinden 3 nükleotid farkı olan iki form halinde bulunur (miR-208a ve b) miR-208a, α-MHC geninin intron bölgesinde kodlanmıştır. Mir-208, β-MHC gen ekspresyonunun kuvvetli pozitif bir düzenleyicisidir. miR-208a’nın uzun süreli genomik deplesyonu kontraktilitede azalmaya ve AF’ye yol açar (fare modeli)* *Callis TE, J Clin Invest. 2009;119:2772–86.")
29
Eric M. Small & Eric N. Olson; Nature 2011

30
KY tanısında miR İnsan KY’de en azından 16 farklı mikro RNA’nın düzeyleri değişir. Serum miR-423 seviyesi, küçük bir çalışmada KY tanısında çok başarılı bulunmuştur (AUC:0.91) Başka bir çalışmada miR-499 düzeyleri ADKY’de yüksek bulunmuş, diyastolik KY’de normal olduğu saptanmıştır.
 Başka bir çalışmada miR-499 düzeyleri ADKY’de yüksek bulunmuş, diyastolik KY’de normal olduğu saptanmıştır.")
31
KY&inflamasyon ilişkisinde miRNA
Van de Vrie M et al. Cardiovasc Drugs Ther. 2011

32
Tedavi Yaklaşımında miRNA

33
Klinik Pratikte Yeri? miR-122, hepatit C virüsünün replikasyonu için gereklidir. Bu mikroRNA partikülünün, yine nükleik asit yapısıyla bloke edilmesinde son aşamaya gelinmiştir. Miravirsen, (SC injection, SPC3649), şu anda klinik çalışmada test edilmektedir (ClinicalTrials.gov identifier: NCT )
, şu anda klinik çalışmada test edilmektedir (ClinicalTrials.gov identifier: NCT )")
34
“MicroRNAs are small stars in the genome galaxy with many suprises still in store”
Sevignani C et al. Mamm Genome ;17(3):
:")
Benzer bir sunumlar

>")






 ZM106 Biyokimya 11. Hafta.>")







>")



