Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Santral Sinir Sistemi Nöromediyatörleri
Prof. Dr. Hakan KARADAĞ

2
Nöroregülatör Nöroregülatör Nörotransmiter Nöromodülatör Nörohormon
Endokrin iletişim Parakrin iletişim Otokrin iletişim

3
Nörotransmiter - Nöromodülatör
Presinaptik uçtan sinaps aralığına salıverilir. Postsinaptik membranı (nöron, efektör hücre) etkiler. Tek başlarına uyarı iletimini sağlar. NÖROMODÜLATÖR Presinaptik uçtan sinaps aralığına salıverilir. Postsinaptik membranı (nöron, efektör hücre) etkiler. Tek başlarına uyarı iletimini sağlayamaz; nörotransmiterin etkilerini modüle eder.
 etkiler. Tek başlarına uyarı iletimini sağlar. NÖROMODÜLATÖR. Presinaptik uçtan sinaps aralığına salıverilir. Postsinaptik membranı (nöron, efektör hücre) etkiler. Tek başlarına uyarı iletimini sağlayamaz; nörotransmiterin etkilerini modüle eder.")
4
Nöroregülatör Nöroregülatör Nörotransmiter Nöromodülatör Nörohormon
Endokrin iletişim Parakrin iletişim Otokrin iletişim Nöromediyatör

5
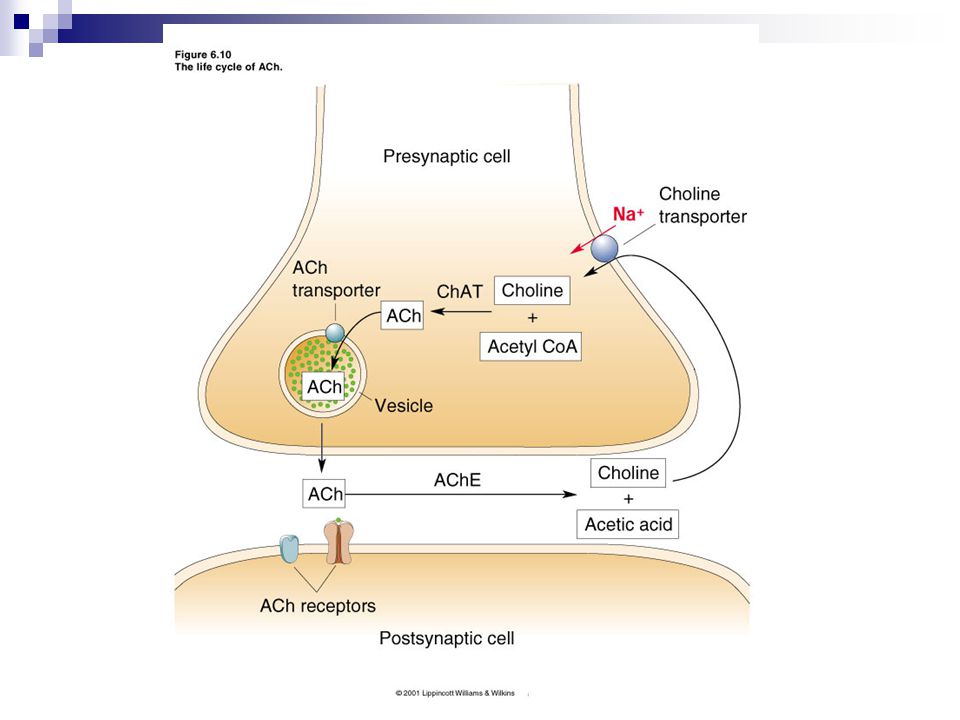
Nöromediyatörlerin sentez, depolanma ve salıverilmeleri

6
Küçük moleküllü nöromediyatörler
Peptidler

7
Nöroregülatör Nöroregülatör Nörotransmiter Nöromodülatör Nörohormon
Endokrin iletişim Parakrin iletişim Otokrin iletişim Nöromediyatör

8
Sinapslar D disinhibisyon B İnhibitör Eksitatör - + + E C A - + YANIT

9
Presinaptik reseptörler
Otoreseptör Heteroreseptör

10
Ko-transmiter A B Nörotransmiter Nöromodülatör Nöromediyatör

11
Küçük moleküllü mediyatörlerin ve nöropeptidlerin farklı uyaranlarla salıverilmeleri

12
Presinaptik reseptörler

13
Presinaptik reseptörler
HETERORESEPTÖR (-) OTORESEPTÖR Presinaptik reseptörler
 OTORESEPTÖR. Presinaptik reseptörler.")
14
SSS Nöromediyatörleri
Amin yapılı nöromediyatörler Asetilkolin Amino asit nöromediyatörler Peptid yapılı nöromediyatörler Diğerleri

15
Amin yapılı nöromediyatörler
Monoaminler Katekolaminler Dopamin Noradrenalin Adrenalin Serotonin Histamin

16
Amino asit nöromediyatörler
İNHİBİTÖR GABA Glisin EKSİTATÖR Glutamat Aspartat

17
Peptid yapılı nöromediyatörler (Nöropeptidler)
Opiodler P Maddesi (Substance P) Somatostatin VIP Kolesistokinin Nörotensin İnsülin Gastrin Glukagon TRH, GnRH, ACTH Anjiotensin II Bradikinin Vazopressin Oksitosin Motilin, Sekretin Nöropeptid Y, CGRP Nörokinin A ve B Galanin Karnozin
 Somatostatin. VIP. Kolesistokinin. Nörotensin. İnsülin. Gastrin. Glukagon. TRH, GnRH, ACTH. Anjiotensin II. Bradikinin. Vazopressin. Oksitosin. Motilin, Sekretin. Nöropeptid Y, CGRP. Nörokinin A ve B. Galanin. Karnozin.")
18
Diğerleri NO Adenozin Steroidler (Aldosteron, Kortizol, Progesteron, Estrojenler, Testosteron) Prostaglandinler
 Prostaglandinler.")
19
Dopaminerjik Sistem

20
Dopaminerjik Uç Tirozin HVA DOPA Dopamin (DA) Rezerpin DA DOPAC Kokain
Tirozin hidroksilaz DOPA HVA Aromatik amino asit dekarboksilaz KOMT Dopamin (DA) MAO Rezerpin DA DOPAC Kokain Amfetamin DA
 MAO. Rezerpin. DA. DOPAC. Kokain. Amfetamin. DA.")
21
Dopaminerjik sistem

25
The nigrostriatal dopamine pathway and the mesolimbic dopamine pathway
The nigrostriatal dopamine pathway (arrows showing connections from substantia nigra to caudate nucleus) and the mesolimbic dopamine pathway (arrows showing connections from ventral tegmentum to frontal cortex) are shown. The density of receptor types (D2 and D4 synapses shown) differ in these two pathways, and the regulation of synaptic dopamine by reuptake may differ, due to differences in density of dopamine transporters (shown as the primary site of action of methylphenidate, which blocks the reuptake process).
 and the mesolimbic dopamine pathway (arrows showing connections from ventral tegmentum to frontal cortex) are shown. The density of receptor types (D2 and D4 synapses shown) differ in these two pathways, and the regulation of synaptic dopamine by reuptake may differ, due to differences in density of dopamine transporters (shown as the primary site of action of methylphenidate, which blocks the reuptake process).")
30
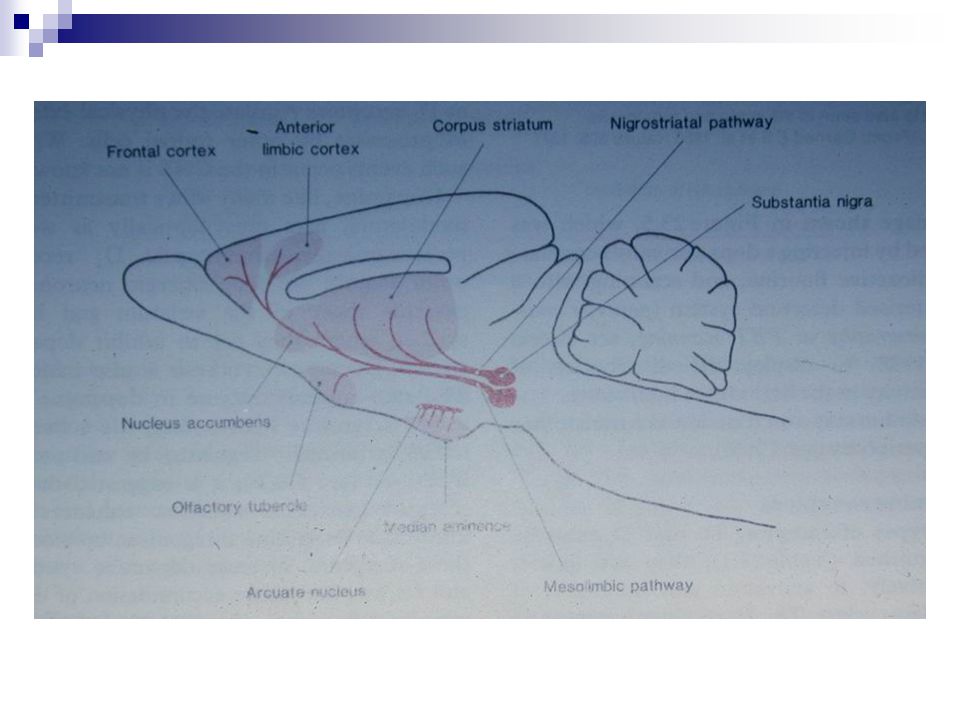
Fonksiyonları Nigrostriatal yolak
Parkinson hastalığında hipoaktivite Mezolimbik yolak ve mezokortikal yolak Şizofrenide artmış etkinlik (?) Pozitif pekiştiri (Nucleus accumbensteki uçlarda dopaminerjik etkinin güçlendirilmesine yol açan ilaç ve maddeler) Tuberoinfundibuler yolak Nöroendokrin fonksiyon Prolaktin, büyüme hormonu ve gonadotropinlerin salgılanmasında azalma UpToDate’den Dopamine agonists – Although dopamine does not decrease gonadotropin secretion to an appreciable degree in normal subjects, bromocriptine has been reported to reduce the secretion of intact gonadotropins and the free alpha subunit in a few patients and even to improve vision in one patient; however, it has not been shown to reduce adenoma size [14,15].
 Pozitif pekiştiri (Nucleus accumbensteki uçlarda dopaminerjik etkinin güçlendirilmesine yol açan ilaç ve maddeler) Tuberoinfundibuler yolak. Nöroendokrin fonksiyon. Prolaktin, büyüme hormonu ve gonadotropinlerin salgılanmasında azalma. UpToDate’den. Dopamine agonists – Although dopamine does not decrease gonadotropin secretion to an appreciable degree in normal subjects, bromocriptine has been reported to reduce the secretion of intact gonadotropins and the free alpha subunit in a few patients and even to improve vision in one patient; however, it has not been shown to reduce adenoma size [14,15].")
31
James Olds ve Peter Milner, 1954 Kaliforniya Teknoloji Enstitüsü
Thirty years ago the first complete report (Olds & Milner, 1954) of positive reinforcement from direct electrical stimulation of the brain was published. We are still a little stunned by that revelation. We were and are imbued with the idea that the brain is marvelously complicated. We are also faced with mounting evidence that it is even more complicated then we imagined (Snyder, 1980, for example, predicted over 200 neurotransmitters will be discovered). Our own experience tells us that pleasure and reinforcement processes are varied and subtle. Yet, a gross manipulation by way of stimulation with a macroelectrode can produce positive affect, and when there is a contingency, this can be reward or positive reinforcement. All that is necessary or positive reinforcement. All that is necessary is to step down the house current to a level that does not destroy neural tissue and to direct it, in brief spurts, to any number of sites in the brain and positive affect is initiated. Furthermore, it seems that in all vertebrates (from samples as diverse as goldfish to human beings) an effect can be elicited by brain stimulation that maintains behavior for that stimulation (Heath, 1964; Olds, 1962). Introduction *The term "abuse liability" is extraordinarily confusing, particularly in the context of this chapter. Discussing all of the problems with the term is beyond the scope of this chapter. It will be sufficient to say here, first, we are concerned with addiction liability. Second, the word liability has two meanings: likelihood and debt. In this context, and perhaps the entire book, it seems that we are attempting to assess addiction likelihood rather than addiction debt or abuse (albeit a likely consequence of addiction). Consequently, my topic is the use of procedures involving ICS in establishing likelihood of addiction. In accordance with modern theory of addiction, particularly opioid addiction (Smith & Lane, 1983), an addiction likelihood, in turn, is strongly related to the potential for a drug to be positively reinforcing. When the possibility of positive intracr When the possibility of positive intracranial reinforcement (ICR) was first announced, it was met, not surprisingly, with inordinate skepticism. That skepticism was extant even though the pioneering work of Hess had shown that intracranial stimulation (ICS) elicited a number of motivational and emotional sequences in freely moving animals. Since Olds used in his initial studies small experimental spaces with large manipulanda, it was suggested, for example, that the rats were not voluntarily pressing to get ICS but that they were merely being thrown on the manipulanda by ICS-elicited forced movements. Rats eventually ran mazes, crossed electrified grids, worked a variety of devices, and solved complex discrimination problems for ICS. So, it is no mere automatism that leads rats to work for ICS (Olds, 1962). The standard experimental space is a box similar to the one popularized by Skinner and colleagues. The rat is placed into a box that has a bar or lever extending through one wall. With depression ending through one wall. With depression of the lever, events can be programmed, such as delivery of food, water or ICS. Using such an experimental procedure, an extensive body of knowledge has been developed as variables, (e.g., degree of food deprivation and rate of food-delivery relative to lever-pressing) were manipulated. The effects of making ICS a contingency of lever-pressing, therefore, can be understood in the context of this extensive information. To observe ICR, subjects (usually rats) are fixed with chronically indwelling electrodes. The surgical procedures have become standard and electrodes and holders for electrodes are commercially available. In brief, all preparations involve putting wires into the brain so that electric current can stimulate a small amount of neural tissue making that tissue supra-active with respect to the rest of the brain in the freely moving subject. At a site of ICS, intense or prolonged current flowing in one direction destroys neural tissue. The typical ICS, then, is low intensity current with alternating polarity. Even if, however, the current is set so as to not damage tissue, ICS is unambiguously positive only when it is of brief duration. So, the standard experimental arrangement is to put a rat in a box and program it so that a lever-press yields a train of pulsate current with alternating polarity of low intensity lasting 0.5 seconds or less (i.e., train duration = 0.5 seconds). Sites of positive ICR differ. Once pressing test of positive ICR differ. Once pressing has been established, the differences are made obvious by changing intensity or duration of ICS. Also, for some sites rats more readily accept experimenter-imposed, frequently occurring trains of ICS whereas at other sites they will respond to terminate the imposed ICS. Sites are classed a "pure positive" when the rat works to initiate ICS and does little to escape it when imposed and "ambiguous" when it both initiates and escapes the ICS (Olds, 1962). This classification is highly dependent on the ICS used in testing. The ICS at all sites can be made too intense, too frequent, or too prolonged for optimal acceptance by the subject. With ICS set at safe values and brief train durations, sites of ICS were varied while observing the subjects’ responsiveness. With some sites rats do not learn to press for ICS. Using other experimental arrangements, it is possible to show that rats will work to avoid getting some ICS that was not positive. ICS, in a standard form, is either positive, neutral, or aversive in affective tone. Figure 1: The typical arrangement for observing intracranial reinforcement. We infer that the medial forebrain bundle (MFB) as it extends from the ventral midbrain through the lateral hypothalamus and lateral preoptic areas is particularly relevant to reinforcement processes because MFB ICS is often unambiguously positive. Rats emit many presses for hours each day for MFB ICS. With MFB sites the minimum intensity for eliciting pressing is low. Rats press tes the minimum intensity for eliciting pressing is low. Rats press for MFB ICS across an extensive range of features of the ICS itself (intensity, train duration, frequency of pulses, et cetera). And, rats do not escape imposition of MFB ICS across many variations of its imposition. One general feature of the behavior emitted for ICS is that the rats work more for greater intensity (more accurately, microcoulombs) of ICS (Keesey, 1964; McIntire & Wright, 1965). For many MFB sites the relationship between rate of pressing and intensity of current is linear up to the point of neural damage. For other sites the function reaches an asymptote so that further increases in intensity produce no further increases in pressing or may even produce a decrease. Observations make it apparent why some rates of pressing may decrease at higher intensities; the higher intensity may elicit seizures or forced movements interfering with pressing. It appears to be necessary to activate a rather large group of neurons and fibers to elicit ICR. The threshold for perception of ICS (i.e., intensity sufficient for a rat to use it as a conditional stimulus) is less than the intensity that will sustain a minimal rate of pressing. After the threshold for ICR is achieved, it seems that the recruitment of more and more activity in relevant tissue yields more and more intense positive affect which is manifested in greater rates of pressing. When eventually an intensity is reached that entually an intensity is reached that elicits activity leading to seizures or other side effects, the affective quality is diluted or the ability to press is hindered. With the initial studies there were a number of observations indicating that ICR had some peculiar features. In contrast to the behavior of rats working for small bits of food, for example, it was concluded that when a delay was programmed between trains of ICS, operant behavior deteriorated. These peculiarities of ICR, compared to behavior following conventional reinforcement, became known as the anomalies of ICR and were provided in lists to support various theories (Deutsch & Howarth, 1963; Kimble, 1961). Research on ICR involved an inspection of reputed anomalies (e.g., Reid, 1967). This research was a test, despite some limitations, of the general idea that certain ICS was indeed initiating activity ordinarily initiated by conventional reinforcers. Although it is impossible to prove an identity, the more the two kinds of reinforcers control behavior in the same way, the more confidence one has that they have considerable neural features in common. There is, of course, the obvious difference that one process follows ICS and one follows events such as eating or drinking. Also, with eating and drinking there is the eventual consequence of a full stomach that has no obvious match with ICR. Keeping these differences in mind, it seemed that if ICS was initiating, it seemed that if ICS was initiating the processes of reinforcement, then ICS should control behavior in nearly the same way as conventional reinforcers. The first finding of the systematic inspection of the anomalies was that some were not apparent when ICR was from MFB stimulation. For example, on the basis of observations of cats, it was concluded that cats did not show good signs of ICR and, therefore, ICS was not activating a universal reinforcement system present in different species. We (Schnitzer, Reid, & Porter, 1965) merely placed electrodes so that the MFB was stimulated and found that cats worked very hard for ICS. Also, direct comparisons were made between behavior maintained by food and water and that maintained by ICR. Prior to these experiments, the comparisons were made indirectly by comparing the behavior of rats working for ICR to the generalizations derived from studying rats working for conventional rewards. When rats press for food, the lever press activates a dispenser that delivers food in a dish usually slightly to the side of the lever. When rats pressed for ICS, the ICR was delivered with the depression of the lever. We (Reid & Porter, 1965) then arranged test conditions so that the lever press merely made it possible to get ICS at the dish that usually received food (i.e., the rat broke a photobeam across the dish to receive ICS). Under these comparable conditions of ways of delivering the tconditions of ways of delivering the two kinds of reinforcers and when the ICS was of the MFB, the behaviors maintained by ICS and by conventional reinforcers were indistinguishable in topography (e.g., Gibson, Reid, Sakai, & Porter, 1965). Rats have been shown to work on periodic schedules of delivery of ICS, to show no peculiarities of extinction (Gibson et al., 1965), and to press without decrement even when intervals between opportunities to press are great (Hunsicker & Reid, 1974; Wasden, Reid, & Porter, 1965). The anomalies of ICR are related to two features: the site of ICS and the comparisons of performance after unequal tasks (Reid, 1967). The site of ICS is a major consideration. There are no gross anomalies of ICR with certain sites of ICS, namely those of the MFB. Rats do show anomalies when tissue related to positive affect is activated concurrently with other tissue. This can be due to a slightly misplaced electrode or to features of the ICS such as high intensity, prolonged durations, or very brief surges in current that accompany generation of ICS by some stimulators. They perform like rats getting bitter food or food accompanied by low level foot shock. They show vacillation, performance decrements, rapid extinction, et cetera. More recent anatomical study of ICR has both expanded the borders of the MFB system and made more precise the cells and fibers of the system. It has become clear that among the critical tissues are dopaminergic neurons and processes (Wise, 1983). The MFB and adjacent tissue are places where dopaminergic fibers are reasonably densely packed (Jacobowitz & Palkovits, 1974), and ICR is achieved when ICS activates these dopaminergic elements (Wise, 1983). By observing when anomalies were and were not present, the features of the system became more defined. When ICS was exclusive to the MFB, anomalies were less apparent or weFB, anomalies were less apparent or were absent. Arrangements for the clearest cases of ICR supported the idea that activity of the MFB system was, indeed, an important element of the behavior of positive reinforcement. There are other lines of converging evidence to support a conclusion that the MFB system with its components of dopaminergic cells and fibers is critical to positive affective processes. For example, reward and positive reinforcement procedures fail when the MFB system is debilitated (Wise, 1982a). Experiments showing that the rewards of prototypic addictive drugs have localized, critical effects within the MFB system are, of course, particularly germane (Bozarth, 1983; Wise, 1983). These lines of research have received extensive review and discussion (e.g., Wise, 1980; 1982b; 1983) and this is not the place to repeat them. What is controversial is whether or not the MFB system of dopaminergic elements is part of the brain’s only system for expressi brain’s only system for expression of positive affect and the consequent events of positive reinforcement. If drugs of addiction are taken for their positively reinforcing effects and if the MFB system is the only system of positive affect, as retina, optic nerves and geniculate are exclusive to vision, then drugs of addiction must act, either directly or indirectly, on the MFB system. The problem of measuring addiction likelihood then becomes one of measuring drug-induced increases in that functional activity. The MFB system need not be, however, the only system of positive affect for measures of drug effects on the MFB system to be useful. The MFB system need only be a major part of the brain’s apparatus for positive affect. A test drug that would increase MFB’s functional activity would have high addiction likelihood. Along the same lines the ability of a drug to elicit positive affect need not be the only reason for a drug to be taken recreationally. All that is necessary is that the ability to elicit positivey is that the ability to elicit positive affect be a major component of the addiction syndrome. As the basic information of ICR accumulated, other events happened to produce an interest in the effects of psychotropic drugs on ICR. Most importantly, there was further confirmation of the idea that many, if not all, addictive drugs were taken for their positively reinforcing aspects (Deneau, Yanagita, & Seevers, 1969; Schuster & Thompson, 1969; Thompson & Schuster, 1964; Weeks, 1962). I doubt if anyone will challenge the conclusion that the MFB system is a major component of the brain’s apparatus for positive affect. (They may challenge the use of the term positive affect; however, they may substitute their favorite synonym without taking away from the conclusion.) I doubt if anyone will challenge in the 1980s that a drug’s ability to be positively reinforcing is a major component of a drug’s ability to become the focus of an addiction. Given this consensus, it follows that a drug increasing the functional activity of the MFB system is also a drug having high addiction likelihood. If the MFB system is not the only system of positive affect or if other factors besides the ability to elicit positive affect contribute to addiction likelihood, tests of functional activity of the MFB system may fail to index a drug’s addiction likelihood. Although there is a consensus concerning the primary conclusions, such a consensus does not translate directly into tests of addiction likelihood. Many problematic theoretical and practical issues remain, some of which, but surely not all, are discussed here. The result is the recommendation of a test, involving drug-induced changes in pressing for ICS by rats, as an initial screening procedure for likelihood of addiction. No one is satisfied with our current knowledge of the brain’s appour current knowledge of the brain’s apparatus of positive reinforcement. Little is known, for example, about the afferent patterns of activity that, in ordinary circumstances, set activity in the MFB system. Our lack of knowledge of the brain’s apparatus for reinforcement limits all approaches to assessing addiction likelihood. This obvious point is stated because it seems to be a covert criticism of methods using ICS. Perhaps the limitations to our knowledge are merely more focused when considering ICR. This is an advantage rather than a disadvantage. Two features of responsiveness for MFB ICS seem to index changes in functional activity: measures of threshold for elicitation of ICR and measures of work expended for a fixed intensity of ICS. For either of these indices of MFB activity to be a valid assessment of addiction likelihood, known addictive drugs must have common, systematic effects. Although one may develop techniques for measuring the MFB’s functional activity in preparations other than those involving behaving subjects (e.g., electrical recording; Nelsen & Kornetsky, 1972), measuring the behavior of subjects is likely to provide a more complete assessment of the relevant functional activity. This also follows from the realization that the MFB system itself is defined behaviorally as well as anatomically. Initial results with the effects of addictive agents on pressing for ICS were confusing. Morphine essing for ICS were confusing. Morphine decreased pressing but amphetamine, cocaine, and barbiturates increased it (Crow, 1970; Olds & Travis, 1960; Reid, Gibson, Gledhill, & Porter, 1964; Stein, 1962). Then, Adams, Lorens, and Mitchell (1972) reported that morphine was capable of increasing pressing. With reports verifying and extending that observation (Bush, Bush, Miller, & Reid, 1976; Holtzman, 1976; Koob, Spector, & Meyerhoff, 1975; Lorens & Mitchell, 1973; Marcus & Kornetsky, 1974; Pert, 1975), the idea was supported that drugs taken frequently for their recreational features by people share two properties. At some doses, these agents are self-administered by laboratory subjects and they facilitated responsiveness for rewarding ICS (Bozarth, 1978; Esposito & Kornetsky, 1978; Reid & Bozarth, 1978). There were a number of questions concerning morphine’s (the prototypic addictive agent) ability to increase pressing for ICS: (a) Was the increase a rebound from initial suppression of pressing? (b) Did the increase reflect morphine’s ability to suppress aversive concomitants that could easily accompany ICS? (c) Did the facilitated pressing reflect some increment in positive affect or merely some increased propensity to be active? Each of these questions was addressed experimentally. With large daily doses of morphine (e.g., 10 or 15 mg/kg/day), the period of facilitation moves forward and the initiaacilitation moves forward and the initial depression of press rates wanes. Figure 2 depicts the effects of smaller doses of morphine on pressing for ICS. It is apparent that the effects of morphine, compared to baseline, are characterized by a triple interaction of dose by time after dosing by days of daily dosing (see Figure 3). A further factor is the rate of pressing at baseline. When press rates at baseline are low, the relative increment in pressing can be great. When press rates at baseline are high, there are ceiling effects. The issue of tolerance to the facilitation effect has received considerable attention (Esposito & Kornetsky, 1978). It is clear that the facilitation in responsiveness to ICS shows nothing approaching complete tolerance (Esposito & Kornetsky, 1977). As daily dosing continues, there is clearly tolerance to the initial suppression (Adams et al., 1972; Bush et al., 1976). The facilitated pressing is paralleled by a decrease in the lower threshold for ICR, a topic reviewed extensively by Esposito and Kornetsky (1978; Esposito, Porrino, & Seeger, this volume). Morphine shifts the rate of pressing to intensity of ICS function to the left (Esposito & Kornetsky, 1977). Such findings provide an important confirmation for the idea that morphine is increasing the effectiveness of the ICS. Figure 2: The effects of small doses of morphine on pressing for hypothalamic ICS. There are a few remaining issues concerning tolerance of the facilitation effect. Quantifying the maximum extent of the facilitation is extraordinarily difficult, because the period of peak facilitation may differ with each dose. We do have enough comparisons to conclude that, in general, facilitation does not diminish much, if any, with repeated doses. In fact, peak effect may become larger with repeated doses. Along the same lines we do not have enough data to judge whether the period of enhanced positive affect due to morphine actually becomes briefer with repeated doses. There are reasons to suppose it does. With repeated injections the period of facilitated pressing moves forward closer and closer to the time of injections. Withdrawal symptoms do emerge and the events of withdrawal do diminish pressing (Bush et al., 1976). So, with the advent of withdrawal and the movement of the period of facilitation forward, the net result may be that the period of positive affect is shorter. The data with respect to tolerance provide the first good indication that morphine’s ability to induce analgesia and positive affect phine’s ability to induce analgesia and positive affect are separable. Figure 3: An attempt to depict the triple interaction of the effects of morphine, a standard large dose, on pressing for ICS. Reprinted with permission from Bozarth, Figure 4: Comparison of time-effect relationship for analgesia and pressing for ICS induced by a single administration of morphine (10 mg/kg). The analgesia data are adapted from Hipps, Eveland, Meyer, Sherman, and Cicero, 1976, and Kayan, Woods, and Mitchell, The ICR data are extrapolated from Adams, Lorens, and Mitchell, 1972, and Bush, Bush, Miller, and Reid, Reprinted with permission from Bozarth, Also, the time course of analgesia and positive affect (as indexed by pressing for ICS) are not the same (see Figure 4). There are also other kinds of evidence (to be summarized later) to indicate separation of morphine’s potential forevidence (to be summarized later) to indicate separation of morphine’s potential for analgesia and positive affect. So, morphine probably does not facilitate responding for ICS because it reduces some aversive concomitant of ICS that may accompany it. A direct test (Farber & Reid, 1976) of morphine’s ability to affect positive affect independent of its analgesic properties was done by assessing morphine’s ability to modify responding for positive ICS that was accompanied by a clearly aversive stimulus. Rats were fixed with two electrodes, one for stimulation of MFB and one for ICS that rats would escape. During one phase of testing, rats had only positive ICS as a contingency. During another phase rats had positive ICS followed immediately by aversive ICS as a single contingency of a lever press. Without drugs the programming of the aversive ICS reduced pressing compared to when only positive ICS was a contingency. From dose-response data and from data measuring pressing after self-administrated doses orasuring pressing after self-administrated doses or after equivalent small doses (Collaer, Magnuson, & Reid, 1977; Gerber, Bozarth, & Wise, 1981; also, see Bermudez-Rattoni, Cruz-Morales, & Reid, 1983), it is concluded that the facilitated pressing is not a rebound from any initial suppression that may accompany larger doses. This conclusion is compatible with the idea that morphine-induced facilitation of pressing is due to morphine’s ability to enhance activity in the MFB system. To confirm this notion an independent test of morphine’s capability to produce positive affect was developed. For 20 days rats pressed for positive ICS alone and for combinations of positive and aversive ICS following an injection of morphine. Morphine increased pressing for positive ICS as expected. After a few doses of morphine, morphine did not facilitate pressing for the combination of positive and aversive ICS. If morphine acted by way of diminishing aversiveness, just the opposite would be predicted. It was reasoned that if morphine was producing positive affect at the time it facilitated pressing for ICS, then that increment in positive affect should be manifested in some feature of the rat’s behavior. If the rat experienced that positive affect in a distinctive place, there is a good possibility that the stimuli of that place would be associated (classically conditioned) to the positive affect and would come to have a positive valence. When given a choice between the place of the drug experience and another place, the subject would choose the place where it had experienced the drug. Following this reasoning, rats were placed into one side of an alley while under the influence of morphine and, subsequently, were given the choice of being in that side or the other side (Rossi & Reid, 1976). The rats were confined to one side of the allets were confined to one side of the alley for conditioning at different times after injection of a large dose of morphine. Among the times chosen was the time that morphine readily facilitated pressing for ICS and times before and after morphine-facilitated pressing. The subjects receiving putative conditioning during the time when morphine facilitated pressing for ICS clearly spent more time on the side of the drug experience compared to rats given saline and treated the same way. Rats receiving putative conditioning at times when morphine did not produce clear signs of facilitated pressing did not show a conditioned preference for a side of the alley. The test of morphine’s ability to establish a preference for the place of a drug experience has come to be called the conditional place preference test (CPP test; Bozarth, this volume; van der Kooy, this volume). The test gave an independent verification that rats were experiencing something (positive affect) that established a preference for a place under the same dosing that facilitated pressing for ICS. Under dosing conditions that facilitate pressing for ICS, a CPP is established, the threshold for initiation of ICR is reduced, and the facilitation is difficult to explain by resorting to explanations other than those involving changing affective-reinforcing properties of ICS. Further, doses comparable to those that are self-administered produce facilitated pressing without an apparefacilitated pressing without an apparent interval of suppressed pressing. So, the conclusion is that morphine’s ability to be positively reinforcing is manifested in a number of indices of the rat’s behavior, including facilitating pressing for ICS.
 of positive reinforcement from direct electrical stimulation of the brain was published. We are still a little stunned by that revelation. We were and are imbued with the idea that the brain is marvelously complicated. We are also faced with mounting evidence that it is even more complicated then we imagined (Snyder, 1980, for example, predicted over 200 neurotransmitters will be discovered). Our own experience tells us that pleasure and reinforcement processes are varied and subtle. Yet, a gross manipulation by way of stimulation with a macroelectrode can produce positive affect, and when there is a contingency, this can be reward or positive reinforcement. All that is necessary or positive reinforcement. All that is necessary is to step down the house current to a level that does not destroy neural tissue and to direct it, in brief spurts, to any number of sites in the brain and positive affect is initiated. Furthermore, it seems that in all vertebrates (from samples as diverse as goldfish to human beings) an effect can be elicited by brain stimulation that maintains behavior for that stimulation (Heath, 1964; Olds, 1962). Introduction. *The term abuse liability is extraordinarily confusing, particularly in the context of this chapter. Discussing all of the problems with the term is beyond the scope of this chapter. It will be sufficient to say here, first, we are concerned with addiction liability. Second, the word liability has two meanings: likelihood and debt. In this context, and perhaps the entire book, it seems that we are attempting to assess addiction likelihood rather than addiction debt or abuse (albeit a likely consequence of addiction). Consequently, my topic is the use of procedures involving ICS in establishing likelihood of addiction. In accordance with modern theory of addiction, particularly opioid addiction (Smith & Lane, 1983), an addiction likelihood, in turn, is strongly related to the potential for a drug to be positively reinforcing. When the possibility of positive intracr. When the possibility of positive intracranial reinforcement (ICR) was first announced, it was met, not surprisingly, with inordinate skepticism. That skepticism was extant even though the pioneering work of Hess had shown that intracranial stimulation (ICS) elicited a number of motivational and emotional sequences in freely moving animals. Since Olds used in his initial studies small experimental spaces with large manipulanda, it was suggested, for example, that the rats were not voluntarily pressing to get ICS but that they were merely being thrown on the manipulanda by ICS-elicited forced movements. Rats eventually ran mazes, crossed electrified grids, worked a variety of devices, and solved complex discrimination problems for ICS. So, it is no mere automatism that leads rats to work for ICS (Olds, 1962). The standard experimental space is a box similar to the one popularized by Skinner and colleagues. The rat is placed into a box that has a bar or lever extending through one wall. With depression ending through one wall. With depression of the lever, events can be programmed, such as delivery of food, water or ICS. Using such an experimental procedure, an extensive body of knowledge has been developed as variables, (e.g., degree of food deprivation and rate of food-delivery relative to lever-pressing) were manipulated. The effects of making ICS a contingency of lever-pressing, therefore, can be understood in the context of this extensive information. To observe ICR, subjects (usually rats) are fixed with chronically indwelling electrodes. The surgical procedures have become standard and electrodes and holders for electrodes are commercially available. In brief, all preparations involve putting wires into the brain so that electric current can stimulate a small amount of neural tissue making that tissue supra-active with respect to the rest of the brain in the freely moving subject. At a site of ICS, intense or prolonged current flowing in one direction destroys neural tissue. The typical ICS, then, is low intensity current with alternating polarity. Even if, however, the current is set so as to not damage tissue, ICS is unambiguously positive only when it is of brief duration. So, the standard experimental arrangement is to put a rat in a box and program it so that a lever-press yields a train of pulsate current with alternating polarity of low intensity lasting 0.5 seconds or less (i.e., train duration = 0.5 seconds). Sites of positive ICR differ. Once pressing test of positive ICR differ. Once pressing has been established, the differences are made obvious by changing intensity or duration of ICS. Also, for some sites rats more readily accept experimenter-imposed, frequently occurring trains of ICS whereas at other sites they will respond to terminate the imposed ICS. Sites are classed a pure positive when the rat works to initiate ICS and does little to escape it when imposed and ambiguous when it both initiates and escapes the ICS (Olds, 1962). This classification is highly dependent on the ICS used in testing. The ICS at all sites can be made too intense, too frequent, or too prolonged for optimal acceptance by the subject. With ICS set at safe values and brief train durations, sites of ICS were varied while observing the subjects’ responsiveness. With some sites rats do not learn to press for ICS. Using other experimental arrangements, it is possible to show that rats will work to avoid getting some ICS that was not positive. ICS, in a standard form, is either positive, neutral, or aversive in affective tone. Figure 1: The typical arrangement for observing intracranial reinforcement. We infer that the medial forebrain bundle (MFB) as it extends from the ventral midbrain through the lateral hypothalamus and lateral preoptic areas is particularly relevant to reinforcement processes because MFB ICS is often unambiguously positive. Rats emit many presses for hours each day for MFB ICS. With MFB sites the minimum intensity for eliciting pressing is low. Rats press tes the minimum intensity for eliciting pressing is low. Rats press for MFB ICS across an extensive range of features of the ICS itself (intensity, train duration, frequency of pulses, et cetera). And, rats do not escape imposition of MFB ICS across many variations of its imposition. One general feature of the behavior emitted for ICS is that the rats work more for greater intensity (more accurately, microcoulombs) of ICS (Keesey, 1964; McIntire & Wright, 1965). For many MFB sites the relationship between rate of pressing and intensity of current is linear up to the point of neural damage. For other sites the function reaches an asymptote so that further increases in intensity produce no further increases in pressing or may even produce a decrease. Observations make it apparent why some rates of pressing may decrease at higher intensities; the higher intensity may elicit seizures or forced movements interfering with pressing. It appears to be necessary to activate a rather large group of neurons and fibers to elicit ICR. The threshold for perception of ICS (i.e., intensity sufficient for a rat to use it as a conditional stimulus) is less than the intensity that will sustain a minimal rate of pressing. After the threshold for ICR is achieved, it seems that the recruitment of more and more activity in relevant tissue yields more and more intense positive affect which is manifested in greater rates of pressing. When eventually an intensity is reached that entually an intensity is reached that elicits activity leading to seizures or other side effects, the affective quality is diluted or the ability to press is hindered. With the initial studies there were a number of observations indicating that ICR had some peculiar features. In contrast to the behavior of rats working for small bits of food, for example, it was concluded that when a delay was programmed between trains of ICS, operant behavior deteriorated. These peculiarities of ICR, compared to behavior following conventional reinforcement, became known as the anomalies of ICR and were provided in lists to support various theories (Deutsch & Howarth, 1963; Kimble, 1961). Research on ICR involved an inspection of reputed anomalies (e.g., Reid, 1967). This research was a test, despite some limitations, of the general idea that certain ICS was indeed initiating activity ordinarily initiated by conventional reinforcers. Although it is impossible to prove an identity, the more the two kinds of reinforcers control behavior in the same way, the more confidence one has that they have considerable neural features in common. There is, of course, the obvious difference that one process follows ICS and one follows events such as eating or drinking. Also, with eating and drinking there is the eventual consequence of a full stomach that has no obvious match with ICR. Keeping these differences in mind, it seemed that if ICS was initiating, it seemed that if ICS was initiating the processes of reinforcement, then ICS should control behavior in nearly the same way as conventional reinforcers. The first finding of the systematic inspection of the anomalies was that some were not apparent when ICR was from MFB stimulation. For example, on the basis of observations of cats, it was concluded that cats did not show good signs of ICR and, therefore, ICS was not activating a universal reinforcement system present in different species. We (Schnitzer, Reid, & Porter, 1965) merely placed electrodes so that the MFB was stimulated and found that cats worked very hard for ICS. Also, direct comparisons were made between behavior maintained by food and water and that maintained by ICR. Prior to these experiments, the comparisons were made indirectly by comparing the behavior of rats working for ICR to the generalizations derived from studying rats working for conventional rewards. When rats press for food, the lever press activates a dispenser that delivers food in a dish usually slightly to the side of the lever. When rats pressed for ICS, the ICR was delivered with the depression of the lever. We (Reid & Porter, 1965) then arranged test conditions so that the lever press merely made it possible to get ICS at the dish that usually received food (i.e., the rat broke a photobeam across the dish to receive ICS). Under these comparable conditions of ways of delivering the tconditions of ways of delivering the two kinds of reinforcers and when the ICS was of the MFB, the behaviors maintained by ICS and by conventional reinforcers were indistinguishable in topography (e.g., Gibson, Reid, Sakai, & Porter, 1965). Rats have been shown to work on periodic schedules of delivery of ICS, to show no peculiarities of extinction (Gibson et al., 1965), and to press without decrement even when intervals between opportunities to press are great (Hunsicker & Reid, 1974; Wasden, Reid, & Porter, 1965). The anomalies of ICR are related to two features: the site of ICS and the comparisons of performance after unequal tasks (Reid, 1967). The site of ICS is a major consideration. There are no gross anomalies of ICR with certain sites of ICS, namely those of the MFB. Rats do show anomalies when tissue related to positive affect is activated concurrently with other tissue. This can be due to a slightly misplaced electrode or to features of the ICS such as high intensity, prolonged durations, or very brief surges in current that accompany generation of ICS by some stimulators. They perform like rats getting bitter food or food accompanied by low level foot shock. They show vacillation, performance decrements, rapid extinction, et cetera. More recent anatomical study of ICR has both expanded the borders of the MFB system and made more precise the cells and fibers of the system. It has become clear that among the critical tissues are dopaminergic neurons and processes (Wise, 1983). The MFB and adjacent tissue are places where dopaminergic fibers are reasonably densely packed (Jacobowitz & Palkovits, 1974), and ICR is achieved when ICS activates these dopaminergic elements (Wise, 1983). By observing when anomalies were and were not present, the features of the system became more defined. When ICS was exclusive to the MFB, anomalies were less apparent or weFB, anomalies were less apparent or were absent. Arrangements for the clearest cases of ICR supported the idea that activity of the MFB system was, indeed, an important element of the behavior of positive reinforcement. There are other lines of converging evidence to support a conclusion that the MFB system with its components of dopaminergic cells and fibers is critical to positive affective processes. For example, reward and positive reinforcement procedures fail when the MFB system is debilitated (Wise, 1982a). Experiments showing that the rewards of prototypic addictive drugs have localized, critical effects within the MFB system are, of course, particularly germane (Bozarth, 1983; Wise, 1983). These lines of research have received extensive review and discussion (e.g., Wise, 1980; 1982b; 1983) and this is not the place to repeat them. What is controversial is whether or not the MFB system of dopaminergic elements is part of the brain’s only system for expressi brain’s only system for expression of positive affect and the consequent events of positive reinforcement. If drugs of addiction are taken for their positively reinforcing effects and if the MFB system is the only system of positive affect, as retina, optic nerves and geniculate are exclusive to vision, then drugs of addiction must act, either directly or indirectly, on the MFB system. The problem of measuring addiction likelihood then becomes one of measuring drug-induced increases in that functional activity. The MFB system need not be, however, the only system of positive affect for measures of drug effects on the MFB system to be useful. The MFB system need only be a major part of the brain’s apparatus for positive affect. A test drug that would increase MFB’s functional activity would have high addiction likelihood. Along the same lines the ability of a drug to elicit positive affect need not be the only reason for a drug to be taken recreationally. All that is necessary is that the ability to elicit positivey is that the ability to elicit positive affect be a major component of the addiction syndrome. As the basic information of ICR accumulated, other events happened to produce an interest in the effects of psychotropic drugs on ICR. Most importantly, there was further confirmation of the idea that many, if not all, addictive drugs were taken for their positively reinforcing aspects (Deneau, Yanagita, & Seevers, 1969; Schuster & Thompson, 1969; Thompson & Schuster, 1964; Weeks, 1962). I doubt if anyone will challenge the conclusion that the MFB system is a major component of the brain’s apparatus for positive affect. (They may challenge the use of the term positive affect; however, they may substitute their favorite synonym without taking away from the conclusion.) I doubt if anyone will challenge in the 1980s that a drug’s ability to be positively reinforcing is a major component of a drug’s ability to become the focus of an addiction. Given this consensus, it follows that a drug increasing the functional activity of the MFB system is also a drug having high addiction likelihood. If the MFB system is not the only system of positive affect or if other factors besides the ability to elicit positive affect contribute to addiction likelihood, tests of functional activity of the MFB system may fail to index a drug’s addiction likelihood. Although there is a consensus concerning the primary conclusions, such a consensus does not translate directly into tests of addiction likelihood. Many problematic theoretical and practical issues remain, some of which, but surely not all, are discussed here. The result is the recommendation of a test, involving drug-induced changes in pressing for ICS by rats, as an initial screening procedure for likelihood of addiction. No one is satisfied with our current knowledge of the brain’s appour current knowledge of the brain’s apparatus of positive reinforcement. Little is known, for example, about the afferent patterns of activity that, in ordinary circumstances, set activity in the MFB system. Our lack of knowledge of the brain’s apparatus for reinforcement limits all approaches to assessing addiction likelihood. This obvious point is stated because it seems to be a covert criticism of methods using ICS. Perhaps the limitations to our knowledge are merely more focused when considering ICR. This is an advantage rather than a disadvantage. Two features of responsiveness for MFB ICS seem to index changes in functional activity: measures of threshold for elicitation of ICR and measures of work expended for a fixed intensity of ICS. For either of these indices of MFB activity to be a valid assessment of addiction likelihood, known addictive drugs must have common, systematic effects. Although one may develop techniques for measuring the MFB’s functional activity in preparations other than those involving behaving subjects (e.g., electrical recording; Nelsen & Kornetsky, 1972), measuring the behavior of subjects is likely to provide a more complete assessment of the relevant functional activity. This also follows from the realization that the MFB system itself is defined behaviorally as well as anatomically. Initial results with the effects of addictive agents on pressing for ICS were confusing. Morphine essing for ICS were confusing. Morphine decreased pressing but amphetamine, cocaine, and barbiturates increased it (Crow, 1970; Olds & Travis, 1960; Reid, Gibson, Gledhill, & Porter, 1964; Stein, 1962). Then, Adams, Lorens, and Mitchell (1972) reported that morphine was capable of increasing pressing. With reports verifying and extending that observation (Bush, Bush, Miller, & Reid, 1976; Holtzman, 1976; Koob, Spector, & Meyerhoff, 1975; Lorens & Mitchell, 1973; Marcus & Kornetsky, 1974; Pert, 1975), the idea was supported that drugs taken frequently for their recreational features by people share two properties. At some doses, these agents are self-administered by laboratory subjects and they facilitated responsiveness for rewarding ICS (Bozarth, 1978; Esposito & Kornetsky, 1978; Reid & Bozarth, 1978). There were a number of questions concerning morphine’s (the prototypic addictive agent) ability to increase pressing for ICS: (a) Was the increase a rebound from initial suppression of pressing (b) Did the increase reflect morphine’s ability to suppress aversive concomitants that could easily accompany ICS (c) Did the facilitated pressing reflect some increment in positive affect or merely some increased propensity to be active Each of these questions was addressed experimentally. With large daily doses of morphine (e.g., 10 or 15 mg/kg/day), the period of facilitation moves forward and the initiaacilitation moves forward and the initial depression of press rates wanes. Figure 2 depicts the effects of smaller doses of morphine on pressing for ICS. It is apparent that the effects of morphine, compared to baseline, are characterized by a triple interaction of dose by time after dosing by days of daily dosing (see Figure 3). A further factor is the rate of pressing at baseline. When press rates at baseline are low, the relative increment in pressing can be great. When press rates at baseline are high, there are ceiling effects. The issue of tolerance to the facilitation effect has received considerable attention (Esposito & Kornetsky, 1978). It is clear that the facilitation in responsiveness to ICS shows nothing approaching complete tolerance (Esposito & Kornetsky, 1977). As daily dosing continues, there is clearly tolerance to the initial suppression (Adams et al., 1972; Bush et al., 1976). The facilitated pressing is paralleled by a decrease in the lower threshold for ICR, a topic reviewed extensively by Esposito and Kornetsky (1978; Esposito, Porrino, & Seeger, this volume). Morphine shifts the rate of pressing to intensity of ICS function to the left (Esposito & Kornetsky, 1977). Such findings provide an important confirmation for the idea that morphine is increasing the effectiveness of the ICS. Figure 2: The effects of small doses of morphine on pressing for hypothalamic ICS. There are a few remaining issues concerning tolerance of the facilitation effect. Quantifying the maximum extent of the facilitation is extraordinarily difficult, because the period of peak facilitation may differ with each dose. We do have enough comparisons to conclude that, in general, facilitation does not diminish much, if any, with repeated doses. In fact, peak effect may become larger with repeated doses. Along the same lines we do not have enough data to judge whether the period of enhanced positive affect due to morphine actually becomes briefer with repeated doses. There are reasons to suppose it does. With repeated injections the period of facilitated pressing moves forward closer and closer to the time of injections. Withdrawal symptoms do emerge and the events of withdrawal do diminish pressing (Bush et al., 1976). So, with the advent of withdrawal and the movement of the period of facilitation forward, the net result may be that the period of positive affect is shorter. The data with respect to tolerance provide the first good indication that morphine’s ability to induce analgesia and positive affect phine’s ability to induce analgesia and positive affect are separable. Figure 3: An attempt to depict the triple interaction of the effects of morphine, a standard large dose, on pressing for ICS. Reprinted with permission from Bozarth, Figure 4: Comparison of time-effect relationship for analgesia and pressing for ICS induced by a single administration of morphine (10 mg/kg). The analgesia data are adapted from Hipps, Eveland, Meyer, Sherman, and Cicero, 1976, and Kayan, Woods, and Mitchell, The ICR data are extrapolated from Adams, Lorens, and Mitchell, 1972, and Bush, Bush, Miller, and Reid, Reprinted with permission from Bozarth, Also, the time course of analgesia and positive affect (as indexed by pressing for ICS) are not the same (see Figure 4). There are also other kinds of evidence (to be summarized later) to indicate separation of morphine’s potential forevidence (to be summarized later) to indicate separation of morphine’s potential for analgesia and positive affect. So, morphine probably does not facilitate responding for ICS because it reduces some aversive concomitant of ICS that may accompany it. A direct test (Farber & Reid, 1976) of morphine’s ability to affect positive affect independent of its analgesic properties was done by assessing morphine’s ability to modify responding for positive ICS that was accompanied by a clearly aversive stimulus. Rats were fixed with two electrodes, one for stimulation of MFB and one for ICS that rats would escape. During one phase of testing, rats had only positive ICS as a contingency. During another phase rats had positive ICS followed immediately by aversive ICS as a single contingency of a lever press. Without drugs the programming of the aversive ICS reduced pressing compared to when only positive ICS was a contingency. From dose-response data and from data measuring pressing after self-administrated doses orasuring pressing after self-administrated doses or after equivalent small doses (Collaer, Magnuson, & Reid, 1977; Gerber, Bozarth, & Wise, 1981; also, see Bermudez-Rattoni, Cruz-Morales, & Reid, 1983), it is concluded that the facilitated pressing is not a rebound from any initial suppression that may accompany larger doses. This conclusion is compatible with the idea that morphine-induced facilitation of pressing is due to morphine’s ability to enhance activity in the MFB system. To confirm this notion an independent test of morphine’s capability to produce positive affect was developed. For 20 days rats pressed for positive ICS alone and for combinations of positive and aversive ICS following an injection of morphine. Morphine increased pressing for positive ICS as expected. After a few doses of morphine, morphine did not facilitate pressing for the combination of positive and aversive ICS. If morphine acted by way of diminishing aversiveness, just the opposite would be predicted. It was reasoned that if morphine was producing positive affect at the time it facilitated pressing for ICS, then that increment in positive affect should be manifested in some feature of the rat’s behavior. If the rat experienced that positive affect in a distinctive place, there is a good possibility that the stimuli of that place would be associated (classically conditioned) to the positive affect and would come to have a positive valence. When given a choice between the place of the drug experience and another place, the subject would choose the place where it had experienced the drug. Following this reasoning, rats were placed into one side of an alley while under the influence of morphine and, subsequently, were given the choice of being in that side or the other side (Rossi & Reid, 1976). The rats were confined to one side of the allets were confined to one side of the alley for conditioning at different times after injection of a large dose of morphine. Among the times chosen was the time that morphine readily facilitated pressing for ICS and times before and after morphine-facilitated pressing. The subjects receiving putative conditioning during the time when morphine facilitated pressing for ICS clearly spent more time on the side of the drug experience compared to rats given saline and treated the same way. Rats receiving putative conditioning at times when morphine did not produce clear signs of facilitated pressing did not show a conditioned preference for a side of the alley. The test of morphine’s ability to establish a preference for the place of a drug experience has come to be called the conditional place preference test (CPP test; Bozarth, this volume; van der Kooy, this volume). The test gave an independent verification that rats were experiencing something (positive affect) that established a preference for a place under the same dosing that facilitated pressing for ICS. Under dosing conditions that facilitate pressing for ICS, a CPP is established, the threshold for initiation of ICR is reduced, and the facilitation is difficult to explain by resorting to explanations other than those involving changing affective-reinforcing properties of ICS. Further, doses comparable to those that are self-administered produce facilitated pressing without an apparefacilitated pressing without an apparent interval of suppressed pressing. So, the conclusion is that morphine’s ability to be positively reinforcing is manifested in a number of indices of the rat’s behavior, including facilitating pressing for ICS.")
32
Skinner Box Operant conditioning chamber
From Wikipedia, the free encyclopedia Skinner box An operant conditioning chamber (sometimes skinner box) is a laboratory apparatus used in the experimental analysis of behavior to study animal behavior. The operant conditioning chamber was created by B.F. Skinner while he was a graduate student at Harvard University around It is used to study both operant conditioning and classical conditioning. Contents 1 Structure 2 Research Impact 3 Popular 'Extensions' 4 Skinner Box 5 See also Structure The structure forming the shell of a chamber is a box large enough to easily accommodate the organism being used as a subject. (Common model organisms used include rodents — usually lab rats — pigeons, and primates). It is often sound-proof and light-proof to avoid distracting stimuli. Operant chambers have at least one operandum (or "manipulandum"), and often two or more, that can automatically detect the occurrence of a behavioral response or action. Typical operanda for primates and rats are response levers; if the subject presses the lever, the opposite end moves and closes a switch that is monitored by a computer or other programmed device. Typical operanda for pigeons and other birds are response keys with a switch that closes if the bird pecks at the key with sufficient force. The other minimal requirement of a conditioning chamber is that it have a means of delivering a primary reinforcer or unconditioned stimulus like food (usually pellets) or water. It can also register the delivery of a conditioned reinforcer, such as an LED (see Jackson & Hackenberg 1996 in the Journal of the Experimental Analysis of Behavior for example) as a "token". With such a simple configuration, one operandum and one feeder, it is possible to investigate many psychological phenomena. Modern operant conditioning chambers typically have many operanda, like many response levers, two or more feeders, and a variety of devices capable of generating many stimuli, including lights, sounds, music, figures, and drawings. Some configurations use an LCD panel for the computer generation of essentially any stimulus. Operant chambers can also have electrified nets or floors so that electrical charges can be given to the animals; or lights of different colors that give information about when the food is available. Although the use of shock is not unheard of, Institutional Research Boards (IRB) approval is needed to avoid unnecessary harmful experimentation on animals. Skinner's work did not focus on punishment, and involved a "paw slap" which caused him to conclude, incorrectly, that punishment was ineffective. Works by Azrin, Sidman and others in the 60s and 70s proved this was not the case. Research Impact Skinner's operant chamber allowed him to explore the rate of response as a dependent variable, as well as develop his theory of schedules of reinforcement. The first operant chambers were attached to cumulative records on drums producing characteristic pauses, scallops, and other lines. Operant conditioning chambers have become common in a variety of research disciplines including behavioral pharmacology, and whose results inform many disciplines outside of psychology such as behavioral economics. Popular 'Extensions' Slot machines and online games are sometimes cited as examples of human devices that use sophisticated operant schedules of reinforcement to reward repetitive actions. Skinner's analysis of contingencies can be applied to almost any activity, however, including valuable ones like scientific creativity, writing novels, and artistic exploration and creativity. Skinner Box Skinner is noted to have said that he didn't want to be an eponym. The term Skinner Box is considered by some to be pejorative, and is probably most commonly used by those who are not in the discipline of Experimental analysis of behavior or in psychology.
 is a laboratory apparatus used in the experimental analysis of behavior to study animal behavior. The operant conditioning chamber was created by B.F. Skinner while he was a graduate student at Harvard University around It is used to study both operant conditioning and classical conditioning. Contents. 1 Structure. 2 Research Impact. 3 Popular Extensions 4 Skinner Box. 5 See also. Structure. The structure forming the shell of a chamber is a box large enough to easily accommodate the organism being used as a subject. (Common model organisms used include rodents — usually lab rats — pigeons, and primates). It is often sound-proof and light-proof to avoid distracting stimuli. Operant chambers have at least one operandum (or manipulandum ), and often two or more, that can automatically detect the occurrence of a behavioral response or action. Typical operanda for primates and rats are response levers; if the subject presses the lever, the opposite end moves and closes a switch that is monitored by a computer or other programmed device. Typical operanda for pigeons and other birds are response keys with a switch that closes if the bird pecks at the key with sufficient force. The other minimal requirement of a conditioning chamber is that it have a means of delivering a primary reinforcer or unconditioned stimulus like food (usually pellets) or water. It can also register the delivery of a conditioned reinforcer, such as an LED (see Jackson & Hackenberg 1996 in the Journal of the Experimental Analysis of Behavior for example) as a token . With such a simple configuration, one operandum and one feeder, it is possible to investigate many psychological phenomena. Modern operant conditioning chambers typically have many operanda, like many response levers, two or more feeders, and a variety of devices capable of generating many stimuli, including lights, sounds, music, figures, and drawings. Some configurations use an LCD panel for the computer generation of essentially any stimulus. Operant chambers can also have electrified nets or floors so that electrical charges can be given to the animals; or lights of different colors that give information about when the food is available. Although the use of shock is not unheard of, Institutional Research Boards (IRB) approval is needed to avoid unnecessary harmful experimentation on animals. Skinner s work did not focus on punishment, and involved a paw slap which caused him to conclude, incorrectly, that punishment was ineffective. Works by Azrin, Sidman and others in the 60s and 70s proved this was not the case. Research Impact. Skinner s operant chamber allowed him to explore the rate of response as a dependent variable, as well as develop his theory of schedules of reinforcement. The first operant chambers were attached to cumulative records on drums producing characteristic pauses, scallops, and other lines. Operant conditioning chambers have become common in a variety of research disciplines including behavioral pharmacology, and whose results inform many disciplines outside of psychology such as behavioral economics. Popular Extensions Slot machines and online games are sometimes cited as examples of human devices that use sophisticated operant schedules of reinforcement to reward repetitive actions. Skinner s analysis of contingencies can be applied to almost any activity, however, including valuable ones like scientific creativity, writing novels, and artistic exploration and creativity. Skinner Box. Skinner is noted to have said that he didn t want to be an eponym. The term Skinner Box is considered by some to be pejorative, and is probably most commonly used by those who are not in the discipline of Experimental analysis of behavior or in psychology.")
33
Zevk Alma Merkezleri (Pleasure Centers) Hoşnutsuzluk Merkezleri (Displeasure Centers) Septal alan Lateral hipotalamus Mediyal ön-beyin demeti (Diğerlerinden daha güçlü + pekiştiri sağlar) Ventral tegmental alan Dorsal pons Hipotalamusun mediyal bölgeleri Mid-brain tegmental alanının lateral kısımları
 Ventral tegmental alan. Dorsal pons. Hipotalamusun mediyal bölgeleri. Mid-brain tegmental alanının lateral kısımları.")
34
Slide 8: Reward: drug self-administration Introduce the concept of positive reinforcement or reward. Explain that rats will press a bar to get an injection of cocaine or heroin (self-administration - shown on the left). The rat keeps pressing to get more cocaine or heroin because the drugs make the rat feel so good. This is called positive reinforcement, or reward. Natural rewards include food, water and sex - each is required to maintain survival of our species. Animals and people will continue to exhibit a behavior that is rewarding - and they will cease that behavior when the reward is no longer present. Explain that there is actually a part of the brain that is activated by natural rewards and by artificial rewards such as addictive drugs. This part of the brain is called the reward system. Neuroscientists have been able to pinpoint the exact parts of the brain involved, with the help of the rats. Point to the cartoon on the right and explain that rats will also self-administer addictive drugs directly into their brains -but only into a specific area of the reward system. If the injection needle is moved less than a millimeter away from this crucial area, the rat won't press the lever for more drug. So based on information from working with the rats, scientists have drawn a map of the brain, and located the structures and pathways that are activated when an addictive drug is taken voluntarily. Tell the students that you will show them this "map".
. The rat keeps pressing to get more cocaine or heroin because the drugs make the rat feel so good. This is called positive reinforcement, or reward. Natural rewards include food, water and sex - each is required to maintain survival of our species. Animals and people will continue to exhibit a behavior that is rewarding - and they will cease that behavior when the reward is no longer present. Explain that there is actually a part of the brain that is activated by natural rewards and by artificial rewards such as addictive drugs. This part of the brain is called the reward system. Neuroscientists have been able to pinpoint the exact parts of the brain involved, with the help of the rats. Point to the cartoon on the right and explain that rats will also self-administer addictive drugs directly into their brains -but only into a specific area of the reward system. If the injection needle is moved less than a millimeter away from this crucial area, the rat won t press the lever for more drug. So based on information from working with the rats, scientists have drawn a map of the brain, and located the structures and pathways that are activated when an addictive drug is taken voluntarily. Tell the students that you will show them this map ..")
35
B F Skinner Skinner box --rewards given for response Primary reinforcer Food Drink Sex Generalised reinforcer Prestige Money Success Both Primary and generalised reinforcers can be Positive or Negative Positive reinforcement (Reward) teachers smile, praise, high grade, etc. Reinforcers may be extrinsic or intrinsic. Extrinsic is a reward given by another person, whereas intrinsic comes from within the person (e.g. satisfaction). Reinforcers may be social or material. Social reinforcers would include praise, whereas material reinforcers are concrete items such as sweets. Negative Reinforcement Taking a "bad thing" away e.g. Letting the students off their homework, because they have worked hard recently Punishment Inflicting a bad thing (punishment) because the pupil did the wrong thing Or loss of something good -- detention Punishment does not: illustrate desirable behaviours causes undesirable emotional side effects only suppresses undesirable response
 teachers smile, praise, high grade, etc. Reinforcers may be extrinsic or intrinsic. Extrinsic is a reward given by another person, whereas intrinsic comes from within the person (e.g. satisfaction). Reinforcers may be social or material. Social reinforcers would include praise, whereas material reinforcers are concrete items such as sweets. Negative Reinforcement. Taking a bad thing away e.g. Letting the students off their homework, because they have worked hard recently. Punishment. Inflicting a bad thing (punishment) because the pupil did the wrong thing Or loss of something good -- detention. Punishment does not: illustrate desirable behaviours causes undesirable emotional side effects only suppresses undesirable response.")
36
The Limbic Dopaminergic Neurons Are Involved in Behavioral ActivationThe human brain has relatively few dopaminergic neurons, and these are equally divided between the substantia nigra, which gives rise to the nigrostriatal pathway, and the ventral tegmental area, which gives rise to the mesocorticolimbic projections (Chapter 45). The neurons of the ventral tegmental area form most of the mesolimbic and mesocortical projections involved in reward. These neurons send their axons to the nucleus accumbens, the striatum, and the frontal cortex, three structures thought to be involved in motivation. When animals are trained to stimulate themselves electrically, these stimuli activate dopaminergic neurons in the ventral tegmental area, thereby increasing the output of dopamine at synapses of the mesolimbic and mesocortical projections. Pathways associated with the dopaminergic neurons are also optimal targets for electrical self-stimulation. Rats often choose self-stimulation over food and sex. Receptor blockers such as the antipsychotic drug haloperidol reduce the rewarding effect of food and intracranial self-stimulation. This action is seen as strong evidence that dopamine has a general role in reinforcement mechanisms in limbic areas. These several arguments implicate midbrain dopaminergic neurons in reward-dependent learning. However, dopamine also is essential for sensory-motor coordination. Selective depletion of dopamine from the ventrolateral sector of the striatum impairs orientation to tactile and olfactory stimuli as well as motor coordination. In experiments with drugs that block dopamine receptors in both the limbic and dorsal striatum, it is difficult to know whether the observed reduction in the hedonic value of reinforcers is due to anhedonia and lack of motivation or an inability to respond to the reinforcement. The mesolimbic dopamine system is thought to gate signals that regulate biological drives and motivation. Drugs that facilitate dopamine transmission enhance the processes by which otherwise neutral stimuli acquire incentive or reinforcing properties and facilitate further drug-seeking behavior. It is not clear, however, how the dopamine system mediates reinforcement. Brain stimulation is in a sense an unnatural reward, so we may ask: Are the dopamine neurons important for natural rewards such as food, water, and sex? Many experiments support the idea that dopamine is important not only in mediating the immediate pleasurable aspects of natural rewards, but also in mediating the arousal effects that are predictive of impending rewards. As previously discussed, lesion studies demonstrate that dopamine systems innervating the striatum contribute to feeding, drinking, and other motivated behavior in a crucial way. This view is supported by studies of intracranial self-stimulation and the demonstration that pharmacological blockade of dopamine systems impairs feeding behavior. Additional information comes from recordings by Wolfram Schultz and his colleagues of single dopaminergic neurons in alert monkeys while they receive rewards. When a monkey is presented with various appetitive stimuli (eg, fruit juice), dopaminergic neurons respond with short phasic bursts P.1010 of activity. Aversive stimuli like air puffs to the hand or drops of saline to the mouth do not cause these transient activations. Thus, dopaminergic neurons are only activated by novel stimuli that elicit reward. After repeated pairing of visual and auditory cues followed by reward, the time of phasic activation of the dopaminergic neurons changes from firing just after the reward is delivered to firing at the exact time the cue is presented. The changes in dopaminergic activity strongly resemble the transfer of an animal's appetitive behavioral reaction from the unconditional stimulus to the conditional stimulus. These arguments suggest that dopaminergic neurons encode expectations about external rewards In one experiment a naive monkey was required to touch a lever before the appearance of a light. Before training, most of the dopaminergic neurons fired a short burst of action potentials after delivery of the reward. After several days of training, the animal learned to reach for the lever as soon as the light was turned on, and this behavioral change correlated with two striking changes in the firing patterns of the dopaminergic neurons. First, the primary reward no longer elicited a phasic response. Second, the onset of the predictive light now caused a phasic activation in the dopaminergic cells' firing. Again, the changes in dopaminergic activity resemble the transfer of the animal's appetitive behavioral reaction from the unconditional to the conditional stimulus. In trials where the reward is not delivered after the light is turned on, the firing rate of dopaminergic neurons decreases below the basal rate at exactly the time the reward should have occurred. This well-timed decrease in the firing rate of dopaminergic neurons shows that the expected time of reward delivery, based on the occurrence of the light, is also encoded in the fluctuation in dopaminergic activity. In contrast, very few dopaminergic neurons respond to stimuli that predict aversive outcomes. Figure 51-9 Cocaine and nicotine affect the rate of electrical self-stimulation of the brain. As the frequency of the self-stimulation current increases, the rate at which the subject presses a self-stimulation lever increases. In the presence of the drugs animals self-stimulate with a lower-frequency current that was previously ineffective. (Adapted from Wise et al )
, dopaminergic neurons respond with short phasic bursts. P of activity. Aversive stimuli like air puffs to the hand or drops of saline to the mouth do not cause these transient activations. Thus, dopaminergic neurons are only activated by novel stimuli that elicit reward. After repeated pairing of visual and auditory cues followed by reward, the time of phasic activation of the dopaminergic neurons changes from firing just after the reward is delivered to firing at the exact time the cue is presented. The changes in dopaminergic activity strongly resemble the transfer of an animal s appetitive behavioral reaction from the unconditional stimulus to the conditional stimulus. These arguments suggest that dopaminergic neurons encode expectations about external rewards. In one experiment a naive monkey was required to touch a lever before the appearance of a light. Before training, most of the dopaminergic neurons fired a short burst of action potentials after delivery of the reward. After several days of training, the animal learned to reach for the lever as soon as the light was turned on, and this behavioral change correlated with two striking changes in the firing patterns of the dopaminergic neurons. First, the primary reward no longer elicited a phasic response. Second, the onset of the predictive light now caused a phasic activation in the dopaminergic cells firing. Again, the changes in dopaminergic activity resemble the transfer of the animal s appetitive behavioral reaction from the unconditional to the conditional stimulus. In trials where the reward is not delivered after the light is turned on, the firing rate of dopaminergic neurons decreases below the basal rate at exactly the time the reward should have occurred. This well-timed decrease in the firing rate of dopaminergic neurons shows that the expected time of reward delivery, based on the occurrence of the light, is also encoded in the fluctuation in dopaminergic activity. In contrast, very few dopaminergic neurons respond to stimuli that predict aversive outcomes. Figure 51-9 Cocaine and nicotine affect the rate of electrical self-stimulation of the brain. As the frequency of the self-stimulation current increases, the rate at which the subject presses a self-stimulation lever increases. In the presence of the drugs animals self-stimulate with a lower-frequency current that was previously ineffective. (Adapted from Wise et al )")
37
Slide 9: The reward pathway Tell students that this is a view of the brain cut down the middle. An important part of the reward system is shown and the major structures are highlighted: the ventral tegmental area (VTA), the nucleus accumbens (nuc. acc.) and the prefrontal cortex. Also, the pathway connecting these structures is highlighted. The information travels from the VTA to the nucleus accumbens and then up to the prefrontal cortex. Reiterate that this pathway is activated by a rewarding stimulus. [Note to scientists - this is not the only pathway activated by reward, other structures are involved too, but only this part of the pathway is shown for simplicity.]
, the nucleus accumbens (nuc. acc.) and the prefrontal cortex. Also, the pathway connecting these structures is highlighted. The information travels from the VTA to the nucleus accumbens and then up to the prefrontal cortex. Reiterate that this pathway is activated by a rewarding stimulus. [Note to scientists - this is not the only pathway activated by reward, other structures are involved too, but only this part of the pathway is shown for simplicity.].")
38
Slide 10: Injection of cocaine into the nucleus accumbens Demonstrate how scientists located the structures important for the addictive nature of drugs. Show that a rat will self-administer cocaine directly into the nucleus accumbens (or the VTA) to activate the pathway. Point to an area close to the nucleus accumbens or VTA and state that if the injection is placed in this other area, the rat will not press the lever to receive the drug. Indicate that scientists know a lot more than where the drug acts to produce rewarding effects - they also know how the drugs work. Show examples with cocaine, heroin and marijuana.
 to activate the pathway. Point to an area close to the nucleus accumbens or VTA and state that if the injection is placed in this other area, the rat will not press the lever to receive the drug. Indicate that scientists know a lot more than where the drug acts to produce rewarding effects - they also know how the drugs work. Show examples with cocaine, heroin and marijuana..")
39
Mutlaka bakılacak site: http://www. flyfishingdevon. co
Drugs of Abuse Increase the Level of Dopamine Released in the Brain Addictive drugs such as cocaine, amphetamine, opiates, and nicotine act like positive reinforcers. Animals will readily press a lever to give themselves an intravenous infusion of amphetamine, for example. Animals can be conditioned to self-administer addictive drugs directly to certain brain sites through a microcannula. The ability of a drug to act as a positive reinforcer that sustains behavior in experimental animals is highly correlated with the abuse potential of the drug for humans. Drugs of abuse potentiate the reinforcing effects of electrical brain stimulation, reducing the frequency of shock pulses needed to produce a given level of behavioral responses. It is as if the drugs enhance the pleasure produced by electrical brain stimulation (Figure 51-9). Psychoactive drugs that are reinforcing also increase the levels of dopamine released at terminals of the projections of the ventral tegmental area. Some drugs do so by blocking the dopamine transporter. Thus, cocaine and amphetamine both raise the level of dopamine in the nucleus accumbens by blocking the dopamine transporter (Chapter 15), thereby prolonging the time dopamine remains in the synaptic cleft. The dopamine transporter may be the site of action for both cocaine and amphetamine and as such could be a molecular target for drugs developed to control addiction. To test this idea, Marc Caron and associates disrupted the gene encoding the dopamine transporter through homologous recombination in a laboratory strain of mice. Homozygotic mice showed no behavioral activation after cocaine or amphetamine were administered systemically, consistent with the notion that the transporter is a critical participant in the mechanism of amphetamine and cocaine action. Study of slices of the striatum in vitro revealed that amphetamine releases dopamine in the wild-type mice but not in the mutant mice. Although many drugs of abuse modulate dopaminergic transmission, not all of them do so by means of the dopamine transporter. Nicotine, possibly the most addictive and most widely abused drug, increases the level of dopamine in the mesocorticolimbic pathway, as do cocaine and amphetamine. Nicotine enhances release of dopamine by acting on presynaptic cholinergic receptors. This enhancement of dopamine may serve as a constant reinforcement for cigarette smoking. By contrast, mu opioid agonists appear to be rewarding because they inhibit GABA-ergic neurons that normally suppress dopaminergic neurons in the ventral tegmental area. The nucleus accumbens, a target for the action of these drugs of addiction, has two functional sectors: the core and the shell. The shell has strong connections to the limbic system and the hypothalamus and is particularly sensitive to addictive drugs. Thus, intravenous injection of cocaine, morphine, and amphetamine results in greater release of dopamine from the shell of the nucleus. Pathways using other transmitters are also involved in regulating self-stimulation in animals and pleasure in humans. In fact, electrical stimulation of the medial forebrain bundle maintains self-stimulation by activating dopaminergic cells only indirectly. The most effective electrical stimuli activate a group of nondopaminergic neurons in the medial forebrain bundle that project to the midbrain and there activate the ascending dopaminergic neurons (Figure 51-10). Moreover, not all drugs of dependence require the dopamine system. At least some dependence on opiates, alcohol, and benzodiazepines can occur in the absence of dopaminergic mechanisms. Indeed, addiction involves more than just the positive reinforcement derived from the drug and the resulting anticipation of the euphoria it produces. Two other features characterize addiction: tolerance and dependence. Tolerance refers to progressive adaption to the dosage that produces euphoria, so that higher and higher dosages are needed to achieve the same euphoric effect. Dependence refers to the negative visceral consequences of withdrawal of the drug, such as nausea. Thus, drug abuse is driven not only by the rewarding effects of the drug but also by avoidance of the highly aversive effects of withdrawal. Tolerance may be due in part to a drug-induced desensitization of the positive reinforcing system. Likewise, some of the symptoms of withdrawal may be due to a rebound depression of the dopaminergic reinforcing system. Figure Brain-reward circuitry in the rat. Intracranial self-stimulation may act directly on descending myelinated fibers. Suspected sites of drug actions are shown in boxes. Acc = nucleus accumbens; DA = dopaminergic fibers; Enk = enkephalin and other opioid-containing neurons; GABA = GABA-ergic inhibitory interneurons; LC = locus ceruleus; NE = norepinephrine-containing fibers; THC = tetrahydrocannabinol; VTA = ventral tegmental area. (Adapted from Gardner and Lowinson 1993.)
. Psychoactive drugs that are reinforcing also increase the levels of dopamine released at terminals of the projections of the ventral tegmental area. Some drugs do so by blocking the dopamine transporter. Thus, cocaine and amphetamine both raise the level of dopamine in the nucleus accumbens by blocking the dopamine transporter (Chapter 15), thereby prolonging the time dopamine remains in the synaptic cleft. The dopamine transporter may be the site of action for both cocaine and amphetamine and as such could be a molecular target for drugs developed to control addiction. To test this idea, Marc Caron and associates disrupted the gene encoding the dopamine transporter through homologous recombination in a laboratory strain of mice. Homozygotic mice showed no behavioral activation after cocaine or amphetamine were administered systemically, consistent with the notion that the transporter is a critical participant in the mechanism of amphetamine and cocaine action. Study of slices of the striatum in vitro revealed that amphetamine releases dopamine in the wild-type mice but not in the mutant mice. Although many drugs of abuse modulate dopaminergic transmission, not all of them do so by means of the dopamine transporter. Nicotine, possibly the most addictive and most widely abused drug, increases the level of dopamine in the mesocorticolimbic pathway, as do cocaine and amphetamine. Nicotine enhances release of dopamine by acting on presynaptic cholinergic receptors. This enhancement of dopamine may serve as a constant reinforcement for cigarette smoking. By contrast, mu opioid agonists appear to be rewarding because they inhibit GABA-ergic neurons that normally suppress dopaminergic neurons in the ventral tegmental area. The nucleus accumbens, a target for the action of these drugs of addiction, has two functional sectors: the core and the shell. The shell has strong connections to the limbic system and the hypothalamus and is particularly sensitive to addictive drugs. Thus, intravenous injection of cocaine, morphine, and amphetamine results in greater release of dopamine from the shell of the nucleus. Pathways using other transmitters are also involved in regulating self-stimulation in animals and pleasure in humans. In fact, electrical stimulation of the medial forebrain bundle maintains self-stimulation by activating dopaminergic cells only indirectly. The most effective electrical stimuli activate a group of nondopaminergic neurons in the medial forebrain bundle that project to the midbrain and there activate the ascending dopaminergic neurons (Figure 51-10). Moreover, not all drugs of dependence require the dopamine system. At least some dependence on opiates, alcohol, and benzodiazepines can occur in the absence of dopaminergic mechanisms. Indeed, addiction involves more than just the positive reinforcement derived from the drug and the resulting anticipation of the euphoria it produces. Two other features characterize addiction: tolerance and dependence. Tolerance refers to progressive adaption to the dosage that produces euphoria, so that higher and higher dosages are needed to achieve the same euphoric effect. Dependence refers to the negative visceral consequences of withdrawal of the drug, such as nausea. Thus, drug abuse is driven not only by the rewarding effects of the drug but also by avoidance of the highly aversive effects of withdrawal. Tolerance may be due in part to a drug-induced desensitization of the positive reinforcing system. Likewise, some of the symptoms of withdrawal may be due to a rebound depression of the dopaminergic reinforcing system. Figure Brain-reward circuitry in the rat. Intracranial self-stimulation may act directly on descending myelinated fibers. Suspected sites of drug actions are shown in boxes. Acc = nucleus accumbens; DA = dopaminergic fibers; Enk = enkephalin and other opioid-containing neurons; GABA = GABA-ergic inhibitory interneurons; LC = locus ceruleus; NE = norepinephrine-containing fibers; THC = tetrahydrocannabinol; VTA = ventral tegmental area. (Adapted from Gardner and Lowinson 1993.)")
40
http://www. blackwell-synergy. com/doi/abs/10. 1111/j. 1527-3458. 2007
FIG. 5 Dopamine (DA) hypothesis of drug reward and NGB 2904’s actions. The mesolimbic DA system originates from the ventral tegmental area (VTA) in the midbrain and projects predominantly to the nucleus accumbens (NAc) in the forebrain. Almost all addictive drugs activate VTA DA neurons and/or increase extracellular DA levels in the NAc via distinct receptor and cellular mechanisms. DA D3 receptors are located on both presynaptic terminals and postsynaptic (GABAergic) neurons. Antagonism of postsynaptic D3 receptors by NGB 2904 may block D3 receptor-mediated addictive effects, while blockade of presynaptic D3 receptors by NGB 2904 may augment cocaine-enhanced NAc DA (via a disinhibition mechanism), which may subsequently attenuate NGB 2904’s therapeutic actions by inhibiting NGB 2904’s binding to postsynaptic D3 receptors and/or by activating other DA receptors. Zheng-Xiong Xi, Eliot L. Gardner (2007) Pharmacological Actions of NGB 2904, a Selective Dopamine D3 Receptor Antagonist, in Animal Models of Drug Addiction CNS Drug Reviews 13 (2), 240–259. doi: /j x
 hypothesis of drug reward and NGB 2904’s actions. The mesolimbic DA system originates from the ventral tegmental area (VTA) in the midbrain and projects predominantly to the nucleus accumbens (NAc) in the forebrain. Almost all addictive drugs activate VTA DA neurons and/or increase extracellular DA levels in the NAc via distinct receptor and cellular mechanisms. DA D3 receptors are located on both presynaptic terminals and postsynaptic (GABAergic) neurons. Antagonism of postsynaptic D3 receptors by NGB 2904 may block D3 receptor-mediated addictive effects, while blockade of presynaptic D3 receptors by NGB 2904 may augment cocaine-enhanced NAc DA (via a disinhibition mechanism), which may subsequently attenuate NGB 2904’s therapeutic actions by inhibiting NGB 2904’s binding to postsynaptic D3 receptors and/or by activating other DA receptors. Zheng-Xiong Xi, Eliot L. Gardner (2007) Pharmacological Actions of NGB 2904, a Selective Dopamine D3 Receptor Antagonist, in Animal Models of Drug Addiction CNS Drug Reviews 13 (2), 240–259. doi: /j x.")
44
Figure Operant intravenous self-administration apparatus used to deliver response-contingent drug infusions and collect data during i.v. self-administration sessions. Each nosepoke hole should have an aperture of 2.3 cm, and be mounted 1.5 cm above the wire mesh floor. A bulb (e.g., 5 VDC GE-47 bulb) should be mounted inside the hole to provide indirect illumination upon completion of a reinforced response. Responses consist of occlusion of an infrared photoemitter-receiver pair for at least 50 msec followed by nonocclusion; photoemitter/receivers are available at an electronics shop or from Med Associates.
 should be mounted inside the hole to provide indirect illumination upon completion of a reinforced response. Responses consist of occlusion of an infrared photoemitter-receiver pair for at least 50 msec followed by nonocclusion; photoemitter/receivers are available at an electronics shop or from Med Associates..")
45
Figure Responding for a 75 mg/kg ethanol infusion on an FR-3 schedule of reinforcement during 2-hr daily sessions. Nosepoking was reinforced on one side of the chamber ("correct nosepokes"), and nonreinforced on the other. Beta-endorphin-deficient mice acquired responding while wild-type mice did not. Open and closed squares indicate correct and incorrect nosepokes, respectively, for beta-endorphin deficient mice, while asterisks and plus signs indicate correct and incorrect nosepokes for wild-type mice. Beta-endorphin-deficient mice had both a higher rate of responding and a greater percentage of total responses on the correct side compared to wild-type mice. Bars indicate standard errors; n = Reprinted with permission from Grahame et al. (1998).
, and nonreinforced on the other. Beta-endorphin-deficient mice acquired responding while wild-type mice did not. Open and closed squares indicate correct and incorrect nosepokes, respectively, for beta-endorphin deficient mice, while asterisks and plus signs indicate correct and incorrect nosepokes for wild-type mice. Beta-endorphin-deficient mice had both a higher rate of responding and a greater percentage of total responses on the correct side compared to wild-type mice. Bars indicate standard errors; n = Reprinted with permission from Grahame et al. (1998)..")
47
Dopaminerjik Reseptörler
İkinci Ulak Selektif Agonist Selektif Antagonist D1 AS stim. cAMP ↑ Fenoldopam SKF 38393 SCH 23390 SCH 39166 SKF 83566 D2 AS inh. K+ kanal açıl. Ca2+ kanal inh. Bromokriptin Sulpirid Domperidon D3 Haloperidol, Sulpirid ve benzerleri: D3 > D2 D4 AS. İnh. Klozapin: D4 > D3 = D2 Striatumda seyrek olduğundan klozapinin ekstrapiramidal yan etkileri yoktur. D5 Fonksiyonel önemleri bilinmemektedir. Dopamin ve flufenazine karşı daha yüksek afinite gösterirler.

49
Noradrenalin

50
Noradrenerjik Uç Tirozin DOPA Dopamin (DA) NA Rezerpin DA NA
Tirozin hidroksilaz DOPA Yıkım ürünleri Aromatik amino asit dekarboksilaz KOMT Dopamin (DA) NA MAO Rezerpin DA NA DbH Bazı antidepresanlar NA
 NA. MAO. Rezerpin. DA. NA. DbH. Bazı antidepresanlar. NA.")
51
Noradrenalin (ve Adrenalin) Metabolizması
 Metabolizması")
55
Noradrenalin Locus coeruleus (LC) Lateral tegmental NAerjik sistem
Beyin noradrenalin içeriğinin yaklaşık % 70’i LC’de bulunur. LC aktivasyonu: Dikkat kesilme, korku ve alarm durumu Bu tabloya anksiyete, tremor, periferik sempatik hiperaktivasyon eşlik eder. LC tahribi Uyuşukluk, çevreye karşı ilginin azalması Lateral tegmental NAerjik sistem

56
LC’tan inen: Lateral inici yolaklar Dorsal inici yolaklar
Sempatik aktivasyon yapması olasıdır. Dorsal inici yolaklar Antinosiseptif etkiye aracılık ederler. Ventral inici yolaklar Lokomotor fonksiyonun noradrenerjik modülasyonunda rol oynarlar.

57
Noradrenerjik sistemin nöroendokrin etkileri
GnRH salgısında artma GH salgısında artma ACTH salgısında artma Vazopressin salgısında artma

58
Noradrenerjik sistemin nöroendokrin etkileri
SALGIDA ARTMA SALGIDA AZALMA GnRH Büyüme Hormonu ACTH Vazopresin

60
Serotonin (5-Hidroksitriptamin)
")
61
Serotonerjik Uç Triptofan 5-HİAA 5-HTF 5-HT 5-HT 5-HİAA MDMA
hidroksilaz 5-HTF 5-HİAA Aromatik amino asit dekarboksilaz 5-HT History of MDMA MDMA was initially developed by E. Merck Pharmaceutical company in Germany in 1914 in an attempt to produce a new appetite suppressant. It was during this same search in 1912 that the "sister drug" of MDMA, methylenedioxyamphetamine or "MDA" or "Eve", was produced. Neither MDMA nor MDA became commercially successful as appetite suppressants. In the 1950's MDMA was briefly evaluated as an adjunct to psychotherapy based on a reported ability to produce a state of consciousness that promotes willingness towards emotional self-disclosure. The drug is considered to be in a novel psychoactive class known as an "entactogen" (i.e. "to touch within oneself"). Although it never gained general acceptance, in the 1970's and early 1980's the drug was again investigated as an adjunct to psychotherapy based on its reported ability to improve interpersonal communications and enhance emotional awareness.( Shulin, Greer, Lister, Downing) During this period recreational use of MDMA was growing. (Eisner) There is evidence of wide spread abuse as a recreational drug on the college campuses. (Peroutka, Cuomo) In one study of undergraduate students at Stanford University, 39% reported that they had used MDMA at least once, with a mean number of uses of 5.4 uses and a median of 4. (Peroutka). In 1985 the Drug Enforcement Agency in the United States placed MDMA on Schedule 1 of controlled substances,citing increasing recreational use and concern over potential neurological damage. (Lawn) In 1986 it was banned from Great Britain as having no accepted medical purpose and a high potential for abuse. At the time there was evidence from rodent studies suggesting damage to and depletion of serotenergic nerves. Additionally there had been a number of reports of fatalities from overdose MAO 5-HT 5-HİAA MDMA (metilendioksimetamfetamin) Bazı antidepresanlar = ECSTASY 5-HT
. Although it never gained general acceptance, in the 1970 s and early 1980 s the drug was again investigated as an adjunct to psychotherapy based on its reported ability to improve interpersonal communications and enhance emotional awareness.( Shulin, Greer, Lister, Downing) During this period recreational use of MDMA was growing. (Eisner) There is evidence of wide spread abuse as a recreational drug on the college campuses. (Peroutka, Cuomo) In one study of undergraduate students at Stanford University, 39% reported that they had used MDMA at least once, with a mean number of uses of 5.4 uses and a median of 4. (Peroutka). In 1985 the Drug Enforcement Agency in the United States placed MDMA on Schedule 1 of controlled substances,citing increasing recreational use and concern over potential neurological damage. (Lawn) In 1986 it was banned from Great Britain as having no accepted medical purpose and a high potential for abuse. At the time there was evidence from rodent studies suggesting damage to and depletion of serotenergic nerves. Additionally there had been a number of reports of fatalities from overdose. MAO. 5-HT. 5-HİAA. MDMA. (metilendioksimetamfetamin) Bazı antidepresanlar. = ECSTASY. 5-HT.")
62
Mechanism of Action · The primary mechanism of action of MDMA is a potent release of brain serotonin as well as inhibition of serotonin reuptake. MDMA does NOT cause release of serotonin via exocitosis of serotonin containing secretory vesicles. Rather, MDMA uses a unique mechanism that causes a reverse in the direction of the normal inward bound serotonin reuptake channel. (Rudnick) So that the high concentrations of serotonin within the cell flood OUT through the transporter channel. This causes a sudden increase in serotonin in the synapse and at the same time blocks reuptake. This causes a rapid and profound acute depletion of serotonin within 3 to 6 hours. · MDMA itself is taken up into the cell via the serotonin uptake channel and/or through diffusion across the membrane. Once within the cell MDMA is known to deplete stores of tryptophan hydroxylase (TPH) via acute oxidative inactivation. The loss of this enzyme occurs primarily in the cell terminal (as opposed to the cell body with its greater size and metabolic reserves) (Schmidt) The loss of this enzyme leaves the cell terminal open to damage from oxidative stress. This is most likely the reason for serotonergic cell terminal damage seen with MDMA abuse and the most likely explanation for the long-term seroternergic damage. (Schmidt) · Pre-treatment with a serotonin reuptake inhibitor, such as paroxetine (Paxil ®), fluoxetine (Prozac ®) or sertraline (Zoloft ®), blocks the effects of MDMA. (Liechti, Stein, Schmidt) This is probably a competitive event at the serotonin reuptake transporter binding site. Additionally, treatment within one hour of MDMA ingestion with a selective serotonin reuptake inhibitor blocks the loss of tryptophan hydroxylase (TPH), and potentially reduces the nerve terminal damage. (Schmidt) · The ability of MDMA to stimulate both the sympathetic and central nervous systems ultimately results from its structural similarity to the endogenous catecholamines: epinephrine, norepinephrine, and dopamine. This is the basis for their amphetamine-like effects. The methylenedioxylation of the catechol ring is responsible for their hallucinogenic activity. This is based on their potent effects on serotonin release. · A biphasic effect on serotonergic neurons has been observed with MDMA. Acutely serotonin levels fall 3 to 6 hours after drug administration, but return to near normal by 24 hours. Levels then decrease again by 1 week. The acute depletion is secondary to serotonin secretion from the neurons, while the long-term depletion occurs because of toxic degeneration of the serotonergic nerve terminals. · The exact mechanism responsible for the hyperthermia has not been elucidated but is probably related to the serotenergic control of thermo-regulation in the hypothalmus. MDMA causes a loss of self thermo-regulation (Malberg, Drafters). Elevated ambient temperature (room/environment temperature) after MDMA ingestion has been shown to cause an increase in core body temperature, loss of thermo-regulation and an increase the damage seen in serotenergic neurons. (Malberg, Drafters) This is a potentially significant risk factor to those who use MDMA at "rave" parties where the ambient temperature is elevated from the large active crowds and inadequate ventilation facilities. · Acute toxicity from a single ingestion (whether large or small) does NOT appear to be dose related. (Waldo, Rugenthal, Mueller, Henry, Ramcharan). Serum levels have been recorded in acute overdoses as high as 7.72 mg/L and 4.3 mg/L with minimal toxicity (Henry, Regenthal, Barret) These are 20 to 30 times expected serum levels from recreational use. Conversely, fatalities that have been published with histories of small ingestions (one of two tablets) and have had serum levels that supported the history (range 0.1 to 0.4 mg/L) (Henry). Other factors such as environment (ambient temperature), over exertion with inadequate rehydration and perhaps an indiosycratic event appear to play an important role in the majority of severe acute cases. · Long term damage to Serotonergic neurons from MDMA DOES appear to be a dose-related event. However the therapeutic index (the difference between a therapeutic dose and a toxic dose) is very narrow. 1. A dose of 2.5 and 5 mg/kg produced 44 and 90% depletion of serotonin, respectively. (Ricuarte) Neither dopamine nor norepinephrine were depleted, showing MDMA's selective impact on serotonin stores. Structural changes and axonal damage to serotonin nerve fibers in the cerebral cortex was demonstrated by pathological examination at 5 mg/kg (Ricaurte). 2. Single doses of MDMA have been shown to produce long-lasting serotonin depletion (30% of neurons for up to 2 weeks) when administered orally to monkeys at near usual human doses (5 mg/kg) (usual doses in humans are 1 to 4 mg/kg) (Ricaurte). 3. MDMA appears to produce damage to and depletion of serotonergic nerve terminals. Cell body damage of serotonergic neurons in monkeys was shown by Ricaurte, etal. It is unknown at this time if there is axonal regeneration from the damaged cell bodies or if the damage is permanent. Subjective effects Effects desired by users include: increased positive emotion and decreased negative emotion increased sense of well-being increased sociability and feelings of closeness or connection with other people[16] reduced defensiveness and fear of emotional injury[17] a sense of increased insightfulness and introspective ability[16] [7] MDMA, particularly with larger doses, is sometimes reported to cause visual distortions. In a review of studies in which 1.5 to 1.7 mg/kg oral MDMA was administered in their laboratory to 74 people, Vollenweider et al. reported that scenic hallucinations were reported only once, while simple patterns, distorted objects, and flashes of light were commonly reported[17] [edit] Other short-term effects Acute physiological effects include: Pupil dilation with attendant photosensitivity and color perception Impaired vision Nystagmus, rapid involuntary rhythmic eye movement, with the eyes moving quickly in one direction (quick phase), and then slowly in the other (slow phase). General restlessness Loss of appetite Increased heart rate and blood pressure Loss of sleep/ inability to sleep Dehydration Depression after the effects have worn off, the depression can last anywhere from a few hours to several weeks. Trisma (jaw-clenching) and bruxia (grinding of the teeth) Stimulated senses. (hear, touch, smell, etc) Frothing at the corners of the mouth.
 So that the high concentrations of serotonin within the cell flood OUT through the transporter channel. This causes a sudden increase in serotonin in the synapse and at the same time blocks reuptake. This causes a rapid and profound acute depletion of serotonin within 3 to 6 hours. · MDMA itself is taken up into the cell via the serotonin uptake channel and/or through diffusion across the membrane. Once within the cell MDMA is known to deplete stores of tryptophan hydroxylase (TPH) via acute oxidative inactivation. The loss of this enzyme occurs primarily in the cell terminal (as opposed to the cell body with its greater size and metabolic reserves) (Schmidt) The loss of this enzyme leaves the cell terminal open to damage from oxidative stress. This is most likely the reason for serotonergic cell terminal damage seen with MDMA abuse and the most likely explanation for the long-term seroternergic damage. (Schmidt) · Pre-treatment with a serotonin reuptake inhibitor, such as paroxetine (Paxil ®), fluoxetine (Prozac ®) or sertraline (Zoloft ®), blocks the effects of MDMA. (Liechti, Stein, Schmidt) This is probably a competitive event at the serotonin reuptake transporter binding site. Additionally, treatment within one hour of MDMA ingestion with a selective serotonin reuptake inhibitor blocks the loss of tryptophan hydroxylase (TPH), and potentially reduces the nerve terminal damage. (Schmidt) · The ability of MDMA to stimulate both the sympathetic and central nervous systems ultimately results from its structural similarity to the endogenous catecholamines: epinephrine, norepinephrine, and dopamine. This is the basis for their amphetamine-like effects. The methylenedioxylation of the catechol ring is responsible for their hallucinogenic activity. This is based on their potent effects on serotonin release. · A biphasic effect on serotonergic neurons has been observed with MDMA. Acutely serotonin levels fall 3 to 6 hours after drug administration, but return to near normal by 24 hours. Levels then decrease again by 1 week. The acute depletion is secondary to serotonin secretion from the neurons, while the long-term depletion occurs because of toxic degeneration of the serotonergic nerve terminals. · The exact mechanism responsible for the hyperthermia has not been elucidated but is probably related to the serotenergic control of thermo-regulation in the hypothalmus. MDMA causes a loss of self thermo-regulation (Malberg, Drafters). Elevated ambient temperature (room/environment temperature) after MDMA ingestion has been shown to cause an increase in core body temperature, loss of thermo-regulation and an increase the damage seen in serotenergic neurons. (Malberg, Drafters) This is a potentially significant risk factor to those who use MDMA at rave parties where the ambient temperature is elevated from the large active crowds and inadequate ventilation facilities. · Acute toxicity from a single ingestion (whether large or small) does NOT appear to be dose related. (Waldo, Rugenthal, Mueller, Henry, Ramcharan). Serum levels have been recorded in acute overdoses as high as 7.72 mg/L and 4.3 mg/L with minimal toxicity (Henry, Regenthal, Barret) These are 20 to 30 times expected serum levels from recreational use. Conversely, fatalities that have been published with histories of small ingestions (one of two tablets) and have had serum levels that supported the history (range 0.1 to 0.4 mg/L) (Henry). Other factors such as environment (ambient temperature), over exertion with inadequate rehydration and perhaps an indiosycratic event appear to play an important role in the majority of severe acute cases. · Long term damage to Serotonergic neurons from MDMA DOES appear to be a dose-related event. However the therapeutic index (the difference between a therapeutic dose and a toxic dose) is very narrow. 1. A dose of 2.5 and 5 mg/kg produced 44 and 90% depletion of serotonin, respectively. (Ricuarte) Neither dopamine nor norepinephrine were depleted, showing MDMA s selective impact on serotonin stores. Structural changes and axonal damage to serotonin nerve fibers in the cerebral cortex was demonstrated by pathological examination at 5 mg/kg (Ricaurte). 2. Single doses of MDMA have been shown to produce long-lasting serotonin depletion (30% of neurons for up to 2 weeks) when administered orally to monkeys at near usual human doses (5 mg/kg) (usual doses in humans are 1 to 4 mg/kg) (Ricaurte). 3. MDMA appears to produce damage to and depletion of serotonergic nerve terminals. Cell body damage of serotonergic neurons in monkeys was shown by Ricaurte, etal. It is unknown at this time if there is axonal regeneration from the damaged cell bodies or if the damage is permanent. Subjective effects. Effects desired by users include: increased positive emotion and decreased negative emotion. increased sense of well-being. increased sociability and feelings of closeness or connection with other people[16] reduced defensiveness and fear of emotional injury[17] a sense of increased insightfulness and introspective ability[16] [7] MDMA, particularly with larger doses, is sometimes reported to cause visual distortions. In a review of studies in which 1.5 to 1.7 mg/kg oral MDMA was administered in their laboratory to 74 people, Vollenweider et al. reported that scenic hallucinations were reported only once, while simple patterns, distorted objects, and flashes of light were commonly reported[17] [edit] Other short-term effects. Acute physiological effects include: Pupil dilation with attendant photosensitivity and color perception. Impaired vision. Nystagmus, rapid involuntary rhythmic eye movement, with the eyes moving quickly in one direction (quick phase), and then slowly in the other (slow phase). General restlessness. Loss of appetite. Increased heart rate and blood pressure. Loss of sleep/ inability to sleep. Dehydration. Depression after the effects have worn off, the depression can last anywhere from a few hours to several weeks. Trisma (jaw-clenching) and bruxia (grinding of the teeth) Stimulated senses. (hear, touch, smell, etc) Frothing at the corners of the mouth.")
67
Serotonin Rafe Sistemi
Çıkıcı Yolaklar Uyku Korteksteki nöronların senkronizasyonu İnici Yolaklar a. Omurilik arka boynuz: Antinosiseptif b. Omurilik ön boynuz: Uyku sırasında kas tonusunun düzenlenmesi (?)
")
68
Santral Serotonerjik Sistemin Rol Oynadığı Olaylar
Majör depresyon Şizofreni Panik ve anksiyete bozuklukları Uyku İştah (azaltıcı) (Lateral hipotalamusta doygunluk merkezinin uyarılması) Ağrı Endokrin Prolaktin ACTH
 (Lateral hipotalamusta doygunluk merkezinin uyarılması) Ağrı. Endokrin. Prolaktin ACTH ")
70
Figure 9-17. The central serotonergic neuron
Figure The central serotonergic neuron. One hypothesized theory of biologic threshold variability is conceptualizing premature ejaculation (PE) as due to hypersensitivity of 5-HT1A or 5-HT2C receptors, as supported by detailed neuropharmacologic studies. The central serotonergic neuron has not been fully characterized, but 16 different 5-HT receptors (five classes with subtypes) have been identified with varying expression and activity to date. 5-HT activity autoregulation and neural signaling is regulated by multiple mechanisms and receptors. Among these, somatodendritic autoreceptors (5-HT1A receptors), presynaptic autoreceptors (5-HT1B, 5-HT1D receptors) and the 5-HT transporter reuptake system use a variety of different mechanisms to self-regulate their own activity. Each of these mechanisms is a negative feedback system that reduces synaptic cleft 5-HT and prevents overstimulation of the postsynaptic receptors. Additionally, activation of 5-HT1a receptors are attenuated or blocked by activation of 5-HT1C receptors. Increased levels of 5-HT in the synaptic cleft results in downregulation of neural signaling and appears to raise the ejaculatory threshold in response to sexual stimulation. The individual variability of the central 5-HT system may account for the sexual side effect profiles of the widely prescribed SSRI class of drugs. Other neural pathways may also be involved in ejaculatory regulation. Central dopamine (DA) has a key role in sexual behavior and appears to be regulated via D1 and D2 receptors. DA agonists, eg, apomorphine (mixed D1/D2) and quinelorane (D2) facilitate erection and ejaculation whereas dopamine antagonists inhibit sexual responses. DA synthesis and/or release is activated by testosterone []. Central activation of the nitric oxide (NO)/cGMP pathway has been shown to affect dopamine (DA)-mediated behavior []. Thus, NO could be a bridge between testosterone and central dopamine pathways in ejaculatory regulation.
 as due to hypersensitivity of 5-HT1A or 5-HT2C receptors, as supported by detailed neuropharmacologic studies. The central serotonergic neuron has not been fully characterized, but 16 different 5-HT receptors (five classes with subtypes) have been identified with varying expression and activity to date. 5-HT activity autoregulation and neural signaling is regulated by multiple mechanisms and receptors. Among these, somatodendritic autoreceptors (5-HT1A receptors), presynaptic autoreceptors (5-HT1B, 5-HT1D receptors) and the 5-HT transporter reuptake system use a variety of different mechanisms to self-regulate their own activity. Each of these mechanisms is a negative feedback system that reduces synaptic cleft 5-HT and prevents overstimulation of the postsynaptic receptors. Additionally, activation of 5-HT1a receptors are attenuated or blocked by activation of 5-HT1C receptors. Increased levels of 5-HT in the synaptic cleft results in downregulation of neural signaling and appears to raise the ejaculatory threshold in response to sexual stimulation. The individual variability of the central 5-HT system may account for the sexual side effect profiles of the widely prescribed SSRI class of drugs. Other neural pathways may also be involved in ejaculatory regulation. Central dopamine (DA) has a key role in sexual behavior and appears to be regulated via D1 and D2 receptors. DA agonists, eg, apomorphine (mixed D1/D2) and quinelorane (D2) facilitate erection and ejaculation whereas dopamine antagonists inhibit sexual responses. DA synthesis and/or release is activated by testosterone []. Central activation of the nitric oxide (NO)/cGMP pathway has been shown to affect dopamine (DA)-mediated behavior []. Thus, NO could be a bridge between testosterone and central dopamine pathways in ejaculatory regulation.")
71
Serotonerjik Reseptörler
5-HT1 5-HT1A 5-HT1B 5-HT1D 5-HT1E 5-HT1F 5-HT2 5-HT2A 5-HT2B 5-HT2C 5-HT3 5-HT4 5-HT5 5-HT6 5-HT7

72
Serotonerjik Reseptörleri Etkileyen Başlıca İlaçlar
5-HT1A Buspiron (parsiyel agonist) Anksiyolitik 5-HT1AD Sumatriptan (agonist) Anti-migren 5-HT2A, 2C LSD (agonist) Klozapin vb. (antagonist) Halusinojen Antipsikotik 5-HT3 Ondansetron vb (antagonist) Antiemetik
 Anksiyolitik. 5-HT1AD. Sumatriptan (agonist) Anti-migren. 5-HT2A, 2C. LSD (agonist) Klozapin vb. (antagonist) Halusinojen. Antipsikotik. 5-HT3. Ondansetron vb (antagonist) Antiemetik.")
73
Serotonin Receptors Receptors Gene family
Receptors linked to second-message systems 5-HT1A linked to inhibition of adenylyl cyclase 5-HT1B linked to inhibition of adenylyl cyclase 5-HT1D linked to inhibition of adenylyl cyclase 5-HT1E linked to inhibition of adenylyl cyclase 5-HT1F linked to inhibition of adenylyl cyclase 5-HT2A linked to phospholipase and PI turnover 5-HT2B linked to phospholipase and PI turnover 5-HT2C linked to phospholipase and PI turnover 5-HT4 linked to stimulation of adenylyl cyclase 5-HT5 unknown linkage 5-HT6 linked to stimulation of adenylyl cyclase 5-HT7 linked to stimulation of adenylyl cyclase Superfamily of receptors with seven transmembrane regions coupled to G proteins Receptors linked to an ion channel Superfamily of ligand-gated ion channels 5-HT3 5-HT = 5-hydroxytryptamine (serotonin); PI = Phosphatidylinositide.
; PI = Phosphatidylinositide.")
75
Histamin

76
Histamin Mast hücrelerinde Nöronlarda
Mast hücreleri kan damarlarının yakınında yerleştiğinden onların kontrolünde rol oynayabilir. Nöronlarda Hipotalamusta tuberomamiller nükleus Çıkıcı aktive edici bir yolak oluşturur; kortikal uyanca neden olur. (H1 antagonistleri sedasyon ve uyku hali yaparlar.)
")
78
Asetilkolin

81
Figure 27. 11 Important nuclei in regulation of the sleep–wake cycle
Figure Important nuclei in regulation of the sleep–wake cycle. (A) A variety of brainstem nuclei using several different neurotransmitters determines mental status on a continuum that ranges from deep sleep to a high level of alertness. These nuclei include: (left) the cholinergic nuclei of the pons–midbrain junction and the raphe nuclei; and (right) the locus coeruleus and the tuberomammillary nuclues. All have widespread ascending and descending connections to other regions (arrows), which explains their numerous effects. Curved arrows along the perimeter of the cortex indicate the innervation of lateral cortical regions not shown in this plane of section. (B) Location of hypothalamic nuclei involved in sleep. (C) Activation of VLPO induces sleep. Orexin-containing neurons project to different nuclei and produce arousal.
 A variety. of brainstem nuclei using several different neurotransmitters determines mental. status on a continuum that ranges from deep sleep to a high level of alertness. These nuclei include: (left) the cholinergic nuclei of the pons–midbrain junction and. the raphe nuclei; and (right) the locus coeruleus and the tuberomammillary. nuclues. All have widespread ascending and descending connections to other. regions (arrows), which explains their numerous effects. Curved arrows along the. perimeter of the cortex indicate the innervation of lateral cortical regions not shown. in this plane of section. (B) Location of hypothalamic nuclei involved in sleep. (C) Activation of VLPO induces sleep. Orexin-containing neurons project to different. nuclei and produce arousal.")
82
Figure 27. 11 Important nuclei in regulation of the sleep–wake cycle
Figure Important nuclei in regulation of the sleep–wake cycle. (A) A variety of brainstem nuclei using several different neurotransmitters determines mental status on a continuum that ranges from deep sleep to a high level of alertness. These nuclei include: (left) the cholinergic nuclei of the pons–midbrain junction and the raphe nuclei; and (right) the locus coeruleus and the tuberomammillary nuclues. All have widespread ascending and descending connections to other regions (arrows), which explains their numerous effects. Curved arrows along the perimeter of the cortex indicate the innervation of lateral cortical regions not shown in this plane of section. (B) Location of hypothalamic nuclei involved in sleep. (C) Activation of VLPO induces sleep. Orexin-containing neurons project to different nuclei and produce arousal.
 A variety. of brainstem nuclei using several different neurotransmitters determines mental. status on a continuum that ranges from deep sleep to a high level of alertness. These nuclei include: (left) the cholinergic nuclei of the pons–midbrain junction and. the raphe nuclei; and (right) the locus coeruleus and the tuberomammillary. nuclues. All have widespread ascending and descending connections to other. regions (arrows), which explains their numerous effects. Curved arrows along the. perimeter of the cortex indicate the innervation of lateral cortical regions not shown. in this plane of section. (B) Location of hypothalamic nuclei involved in sleep. (C) Activation of VLPO induces sleep. Orexin-containing neurons project to different. nuclei and produce arousal.")
83
Fizyolojik Önemi Öğrenme ve bellek Alzheimer hastalığında kolinerjik hipoaktivite Ekstrapiramidal sistemin dengeli çalışması Kolinerjik/Dopaminerjik oran: Parkinson hastalığı Huntington koresi Giles de la Tourette hastalığı Friedreich hastalığı

84
Asetilkolin Reseptörler
Muskarinik M1 : IP3/DAG Aktivasyonu M2 : AS inhibisyonu, K+ kanallarının açılması, Ca++ kanallarının kapanması, Pre- ve postsinaptik. M3 : IP3 /DAG Aktivasyonu M4 M5 Nikotinik Heterojendir. Pentamerik yapıda olup bu yapıya sadece ve alt birimler girer. Presinaptik yerleşimli olanlar nöromediyatör salıverilmesini artırır.

86
GABA (Gama Amino Bütirik Asit)
")
87
GABA (Gama-amino bütirik asit)
Beynin en yaygın inhibitör nöromediyatörüdür. Genellikle ara nöron şeklindedir.

88
GABA Döngüsü Glutamat GABA TCA DÖNGÜSÜ Süksinik semialdehid Vigabatrin
Gabakulin Glutamik asit dekarboksilaz Glutamat GABA GABA transaminaz TCA DÖNGÜSÜ Süksinik semialdehid

89
GABA Reseptörleri GABAA İçinde klorür kanalı bulunur. İnhibisyon yapar. GABAB İçinde iyon kanalı bulunmaz. G proteini ile kenetlidir. İnhibisyon yapar.

90
GABAA Reseptör Kompleksi
GABA bağlanma yeri (+) Agonist: GABA, musimol Antagonist: bikukulin Barbitürat bağlanma yeri (+) Agonist: barbitüratlar Benzodiazepin bağlanma yeri (+) Agonist: benzodiazepinler Ters agonist: b-CCE, b-CCM Antagonist: flumazenil Benzodiazepin bağlanma yerinin en az 2 alt tipi olduğu saptanmıştır: BZ1 (w1) ve BZ2 (w2) Pikrotoksin bağlanma yeri (-) Agonist: pikrotoksinin, pentilentetrazol Alkol bağlanma yeri (+) Agonist: alkol g-Aminobutyric acid (GABA) is perhaps the most comprehensively studied inhibitory neurotransmitter in the mammalian central nervous system (CNS). It has been estimated that about 40% of synapses in the brain are GABAergic. It is now well recognized that cellular excitability leading to convulsive seizures can be attenuated by GABAergic stimulation in the CNS. Current evidence also indicates that most anxiolytics and hypnotic-sedative drugs such as benzodiazepines and barbiturates exert their pharmacological actions via interactions with a discrete neuronal site on the GABAA Receptor-Benzodiazepine Receptor-Chloride Ion Channel Complex. Unlike other neurotransmitter receptors such as DA and 5-HT discussed earlier, the model of the GABA receptor consists of a multiple receptor complex which provides binding sites for a variety of drugs and/or endogenous biologically active compounds. Stimulation of these binding sites contributes to the responses related to the GABAergic system itself. For this reason it is referred to as a GABAA Receptor-Benzodiazepine Receptor-Chloride Ion Channel Complex. You should refer to pages 220 to 226 of the text for a discussion of the GABAergic synapse as the primary site of action of benzodiazepines and also refer to pages 194 to 197 for a brief discussion of how some anticonvulsants work via activation of GABAergic receptors. Objectives A student should be able to: 1. Reproduce the biosynthetic and metabolic pathways of GABA. 2. Identify the step at which sodium valproate exerts its anticonvulsant response in the above pathways. 3. Reproduce the model of the GABAA Receptor-Benzodiazepine Receptor-Chloride Channel Complex and explain the effect on the receptor and the biological response of each type of drug or endogenous compound, which interacts with the receptor complex. 4. Explain the location and function of GABA’s presynaptic receptor. 5. Account for the mechanism of action of baclofen and its structural relationship to GABA and its structural differences from the more classical muscle relaxants. 6. Account for the mechanism of action of irreversible GABA transaminase (GABA-T) inhibitors. 7. Identify the receptor bound conformation of GABA. 8. Explain why GABA must be present in order for barbiturates or benzodiazepines to exert their pharmacological activity.
 Agonist: GABA, musimol. Antagonist: bikukulin. Barbitürat bağlanma yeri (+) Agonist: barbitüratlar. Benzodiazepin bağlanma yeri (+) Agonist: benzodiazepinler. Ters agonist: b-CCE, b-CCM. Antagonist: flumazenil. Benzodiazepin bağlanma yerinin en az 2 alt tipi olduğu saptanmıştır: BZ1 (w1) ve BZ2 (w2) Pikrotoksin bağlanma yeri (-) Agonist: pikrotoksinin, pentilentetrazol. Alkol bağlanma yeri (+) Agonist: alkol. g-Aminobutyric acid (GABA) is perhaps the most comprehensively studied inhibitory neurotransmitter in the mammalian central nervous system (CNS). It has been estimated that about 40% of synapses in the brain are GABAergic. It is now well recognized that cellular excitability leading to convulsive seizures can be attenuated by GABAergic stimulation in the CNS. Current evidence also indicates that most anxiolytics and hypnotic-sedative drugs such as benzodiazepines and barbiturates exert their pharmacological actions via interactions with a discrete neuronal site on the GABAA Receptor-Benzodiazepine Receptor-Chloride Ion Channel Complex. Unlike other neurotransmitter receptors such as DA and 5-HT discussed earlier, the model of the GABA receptor consists of a multiple receptor complex which provides binding sites for a variety of drugs and/or endogenous biologically active compounds. Stimulation of these binding sites contributes to the responses related to the GABAergic system itself. For this reason it is referred to as a GABAA Receptor-Benzodiazepine Receptor-Chloride Ion Channel Complex. You should refer to pages 220 to 226 of the text for a discussion of the GABAergic synapse as the primary site of action of benzodiazepines and also refer to pages 194 to 197 for a brief discussion of how some anticonvulsants work via activation of GABAergic receptors. Objectives. A student should be able to: 1. Reproduce the biosynthetic and metabolic pathways of GABA. 2. Identify the step at which sodium valproate exerts its anticonvulsant response in the above pathways. 3. Reproduce the model of the GABAA Receptor-Benzodiazepine Receptor-Chloride Channel Complex and explain the effect on the receptor and the biological response of each type of drug or endogenous compound, which interacts with the receptor complex. 4. Explain the location and function of GABA’s presynaptic receptor. 5. Account for the mechanism of action of baclofen and its structural relationship to GABA and its structural differences from the more classical muscle relaxants. 6. Account for the mechanism of action of irreversible GABA transaminase (GABA-T) inhibitors. 7. Identify the receptor bound conformation of GABA. 8. Explain why GABA must be present in order for barbiturates or benzodiazepines to exert their pharmacological activity.")
91
Glisin

92
Glisin Omurilik ve beyin sapında yer alan inhibitör bir nörotransmiterdir. Reseptörü hücre membranında klorür kanalı ile kenetlenme gösterir. Striknin ile antagonize edilir. Tetanus toksini sinir ucundan glisin salıverilmesini inhibe ederek konvülsiyon oluşturur.

93
Glutamat

94
Glutamat Sentez ve yıkım
Glutamin Glutamin sentetaz Glutaminaz Glutamik asit

95
Glutamat Reseptörleri
NMDA reseptörleri (N-metil-D-aspartat) AMPA reseptörleri (kiskalat res.) (a-amino-3-hidroksi-5-metil-4-isoksazol propionat) Kainat reseptörleri Metabotropik glutamat reseptörleri mGlu1, mGlu2, mGlu3, mGlu4, mGlu5, mGlu6, mGlu7, mGlu8
 AMPA reseptörleri (kiskalat res.) (a-amino-3-hidroksi-5-metil-4-isoksazol propionat) Kainat reseptörleri. Metabotropik glutamat reseptörleri. mGlu1, mGlu2, mGlu3, mGlu4, mGlu5, mGlu6, mGlu7, mGlu8.")
96
Fizyolojik etkileri Eksitatör iletilerin taşınması Öğrenme ve bellek
Nöronlarda eksitotoksik etki Ağrı duyusunun iletilmesi

97
NMDA reseptörü Glutamat bağlanma yeri Glisin bağlanma yeri
Poliamin bağlanma yeri Mg++ bağlanma yeri Fensiklidin bağlanma yeri

99
NMDA reseptörü Ca++ Na+ l-glutamat NMDA Glisin Poliaminler İfenprodil
Eliprodil Ketamin Dizosilpin (MK 801) Fensiklidin Mg++
 Fensiklidin. Mg++")
100
Figure 24.5 Diagram of a section
through the rodent hippocampus showing the major regions, excitatory pathways, and synaptic connections. Longterm potentiation has been observed at each of the three synaptic connections shown here.

101
Figure 24.6 Long-term potentiation of
Schaffer collateral-CA1 synapses. (A) Arrangement for recording synaptic transmission; two stimulating electrodes (1 and 2) each activate separate populations of Schaffer collaterals, thus providing test and control synaptic pathways. (B) Left: Synaptic responses recorded in a CA1 neuron in response to single stimuli of synaptic pathway 1, minutes before and one hour after a high-frequency train of stimuli. The high-frequency stimulus train increases the size of the EPSP evoked by a single stimulus. Right: Responses produced by stimulating synaptic pathway 2, which did not receive high-frequency stimulation, is unchanged. (C) The time course of changes in the amplitude of EPSPs evoked by stimulation of pathways 1 and 2. High-frequency stimulation of pathway 1 causes a prolonged enhancement of the EPSPs in this pathway (purple). This potentiation of synaptic transmission in pathway 1 persists for several hours, while the amplitude of EPSPs produced by pathway 2 (orange) remains constant. (After Malinow et al., 1989.)
 Arrangement for recording synaptic. transmission; two stimulating electrodes. (1 and 2) each activate separate populations. of Schaffer collaterals, thus providing. test and control synaptic pathways. (B) Left: Synaptic responses recorded in. a CA1 neuron in response to single. stimuli of synaptic pathway 1, minutes. before and one hour after a high-frequency. train of stimuli. The high-frequency. stimulus train increases the size. of the EPSP evoked by a single stimulus. Right: Responses produced by stimulating. synaptic pathway 2, which did not. receive high-frequency stimulation, is. unchanged. (C) The time course of. changes in the amplitude of EPSPs. evoked by stimulation of pathways 1. and 2. High-frequency stimulation of. pathway 1 causes a prolonged enhancement. of the EPSPs in this pathway (purple). This potentiation of synaptic transmission. in pathway 1 persists for several. hours, while the amplitude of EPSPs. produced by pathway 2 (orange) remains constant. (After Malinow et al., 1989.)")
103
Tim Bliss ve Terje Lømo 1973 Oslo - Norveç
Figure 24.7 Pairing presynaptic and postsynaptic activity causes LTP. Single stimuli applied to a Schaffer collateral synaptic input evokes EPSPs in the postsynaptic CA1 neuron. These stimuli alone do not elicit any change in synaptic strength. However, when the CA1 neuron’s membrane potential is briefly depolarized (by applying current pulses through the recording electrode) in conjunction with the Schaffer collateral stimuli, there is a persistent increase in the EPSPs. (After Gustafsson et al., 1987.)
 in conjunction. with the Schaffer collateral. stimuli, there is a persistent increase in. the EPSPs. (After Gustafsson et al., 1987.)")
104
In cultured cells, applying a series of short, high-frequency electric stimuli to a synapse can strengthen, or potentiate, the synapse for minutes to hours. In living cells, LTP occurs naturally and can last from hours to days, months, and years. Neurons connected by a synapse that has undergone LTP have a tendency to be active simultaneously: after a synapse has undergone LTP, subsequent stimuli applied to the presynaptic cell are more likely to elicit action potentials in the postsynaptic cell.

105
Dinlenme durumunda NMDA reseptörünün kanalı magnezyum tarafından tıkanmıştır ve oradan uzaklaştırılması depolarizasyon ile sağlanır. Bu ya presinaptik uçtan gelen çok güçlü bir uyarıyla (fazla glutamat salıvererek) ya da presinaptik nöronun tekrarlanan (repetitive) uyarılarıyla sağlanabilir. İkinci mekanizmada, postsinaptik membran depolarize hale getirilerek magnezyum uzaklaştırılır; ardından yeterli miktarda kalsiyum hücre içine girebilir. Bu bulgu Hebb’in 1949 yılında öne sürdüğü öngörüyü destekleyen ilk kanıttır: “bir A nöronunun aksonu B nöronunu sürekli ve tekrarlayan bir şekilde uyardığında, B hücre uyarılmasının sürdürülmesini sağlamak için, bu hücrelerden birinde ya da her ikisinde bazı büyüme süreci veya metabolik değişiklikler meydana gelir.” Hücreye giren kalsiyum postsinaptik uçta en az 3 protein kinazı aktive eder: Kalsiyum-kalmodulin-bağımlı protein kinaz II (CaM kinaz II olarak da bilinir) Protein kinaz C Tirozin kinaz, fyn Buradaki kinazlar daha önce anlatılan (3. konu) sAMP bağımlı kinazlardan farklı olmakla birlikte ona eşdeğer işler yapmaktadırlar. Örneğin CaM kinaz II, nonNMDA reseptörlerinin fosforilasyonunu sağlayarak presinaptik uçtan salıverilen glutamata duyarlılığını artırmaktadır. Ayrıca CaM kinaz II’nin intraselüler AMPA reseptörlerinin membrana translokasyonunu sağladığı gösterilmiştir. Ekstrem bir örnek olarak, bazı sinapslarda sadece NMDA reseptörleri bulunur ve bu reseptörler normal sinaptik aşırımı sağlayamazlar; yani bu sinapslar kullanılamaz durumdadırlar. Ancak LTP’den sonra postsinaptik membranda AMPA reseptörlerin ortaya çıkması ile bu sinapslar rutin sinaptik iletide rol almaya başlarlar. Normal koşullar altında tek bir CA3 nöronu sadece tek bir CA1 nöronu ile sadece tek bir sinaptik bağlantı yapar. Bu tek sinaptik bağlantıda da sadece 1 tane aktif zon yer alır ve buradan sadece tek 1 vezikül salıverilir. Bu vezikülün içinde 5000 adet glutamat molekülü bulunmaktadır. LTP’nin oluşmasından önce bu sinaptik bağlantılar etkisiz durumdadır. Presinaptik nöronda oluşan çoğu aksiyon potansiyeli bu 1 adet vezikülü salmakta bile başarısız olmaktadır; postsinaptik membranda sinaptik bir potansiyel oluşturamamaktadır. Ancak, LTP’den sonra aksiyon potansiyellerinin çoğu, bu 1 tek vezikülü salıverebilmekte ve sinaptik potansiyel oluşturabilmektedir. Böylece LTP, aktivitenin bir sinapsın gücünü nasıl artırdığına canlı bir örnektir. Burada, postsinaptik membranda üretilen bir maddenin presinaptik uca difüze olarak orayı etkilediği anlaşılmaktadır. Bu maddenin NO olabileceğine ilişkin bilgilerimiz klasik kitaplarda bulunmaktadır (HK).
 Protein kinaz C. Tirozin kinaz, fyn. Buradaki kinazlar daha önce anlatılan (3. konu) sAMP bağımlı kinazlardan farklı olmakla birlikte ona eşdeğer işler yapmaktadırlar. Örneğin CaM kinaz II, nonNMDA reseptörlerinin fosforilasyonunu sağlayarak presinaptik uçtan salıverilen glutamata duyarlılığını artırmaktadır. Ayrıca CaM kinaz II’nin intraselüler AMPA reseptörlerinin membrana translokasyonunu sağladığı gösterilmiştir. Ekstrem bir örnek olarak, bazı sinapslarda sadece NMDA reseptörleri bulunur ve bu reseptörler normal sinaptik aşırımı sağlayamazlar; yani bu sinapslar kullanılamaz durumdadırlar. Ancak LTP’den sonra postsinaptik membranda AMPA reseptörlerin ortaya çıkması ile bu sinapslar rutin sinaptik iletide rol almaya başlarlar. Normal koşullar altında tek bir CA3 nöronu sadece tek bir CA1 nöronu ile sadece tek bir sinaptik bağlantı yapar. Bu tek sinaptik bağlantıda da sadece 1 tane aktif zon yer alır ve buradan sadece tek 1 vezikül salıverilir. Bu vezikülün içinde 5000 adet glutamat molekülü bulunmaktadır. LTP’nin oluşmasından önce bu sinaptik bağlantılar etkisiz durumdadır. Presinaptik nöronda oluşan çoğu aksiyon potansiyeli bu 1 adet vezikülü salmakta bile başarısız olmaktadır; postsinaptik membranda sinaptik bir potansiyel oluşturamamaktadır. Ancak, LTP’den sonra aksiyon potansiyellerinin çoğu, bu 1 tek vezikülü salıverebilmekte ve sinaptik potansiyel oluşturabilmektedir. Böylece LTP, aktivitenin bir sinapsın gücünü nasıl artırdığına canlı bir örnektir. Burada, postsinaptik membranda üretilen bir maddenin presinaptik uca difüze olarak orayı etkilediği anlaşılmaktadır. Bu maddenin NO olabileceğine ilişkin bilgilerimiz klasik kitaplarda bulunmaktadır (HK).")
106
Uzun Süreli Potansiyalizasyon (Long Term Potentiation, LTP)
Dinlenme durumunda NMDA reseptörünün kanalı magnezyum tarafından tıkanmıştır ve oradan uzaklaştırılması depolarizasyon ile sağlanır. Bu ya presinaptik uçtan gelen çok güçlü bir uyarıyla (fazla glutamat salıvererek) ya da presinaptik nöronun tekrarlanan (repetitive) uyarılarıyla sağlanabilir. İkinci mekanizmada, postsinaptik membran depolarize hale getirilerek magnezyum uzaklaştırılır; ardından yeterli miktarda kalsiyum hücre içine girebilir. Bu bulgu Hebb’in 1949 yılında öne sürdüğü öngörüyü destekleyen ilk kanıttır: “bir A nöronunun aksonu B nöronunu sürekli ve tekrarlayan bir şekilde uyardığında, B hücre uyarılmasının sürdürülmesini sağlamak için, bu hücrelerden birinde ya da her ikisinde bazı büyüme süreci veya metabolik değişiklikler meydana gelir.” Hücreye giren kalsiyum postsinaptik uçta en az 3 protein kinazı aktive eder: Kalsiyum-kalmodulin-bağımlı protein kinaz II (CaM kinaz II olarak da bilinir) Protein kinaz C Tirozin kinaz, fyn Buradaki kinazlar daha önce anlatılan (3. konu) sAMP bağımlı kinazlardan farklı olmakla birlikte ona eşdeğer işler yapmaktadırlar. Örneğin CaM kinaz II, nonNMDA reseptörlerinin fosforilasyonunu sağlayarak presinaptik uçtan salıverilen glutamata duyarlılığını artırmaktadır. Ayrıca CaM kinaz II’nin intraselüler AMPA reseptörlerinin membrana translokasyonunu sağladığı gösterilmiştir. Ekstrem bir örnek olarak, bazı sinapslarda sadece NMDA reseptörleri bulunur ve bu reseptörler normal sinaptik aşırımı sağlayamazlar; yani bu sinapslar kullanılamaz durumdadırlar. Ancak LTP’den sonra postsinaptik membranda AMPA reseptörlerin ortaya çıkması ile bu sinapslar rutin sinaptik iletide rol almaya başlarlar. Normal koşullar altında tek bir CA3 nöronu sadece tek bir CA1 nöronu ile sadece tek bir sinaptik bağlantı yapar. Bu tek sinaptik bağlantıda da sadece 1 tane aktif zon yer alır ve buradan sadece tek 1 vezikül salıverilir. Bu vezikülün içinde 5000 adet glutamat molekülü bulunmaktadır. LTP’nin oluşmasından önce bu sinaptik bağlantılar etkisiz durumdadır. Presinaptik nöronda oluşan çoğu aksiyon potansiyeli bu 1 adet vezikülü salmakta bile başarısız olmaktadır; postsinaptik membranda sinaptik bir potansiyel oluşturamamaktadır. Ancak, LTP’den sonra aksiyon potansiyellerinin çoğu, bu 1 tek vezikülü salıverebilmekte ve sinaptik potansiyel oluşturabilmektedir. Böylece LTP, aktivitenin bir sinapsın gücünü nasıl artırdığına canlı bir örnektir. Burada, postsinaptik membranda üretilen bir maddenin presinaptik uca difüze olarak orayı etkilediği anlaşılmaktadır. Bu maddenin NO olabileceğine ilişkin bilgilerimiz klasik kitaplarda bulunmaktadır (HK). İlk başlarda bu maddenin NO olabileceği düşünülmekte iken şimdilerde hücre adezyon molekülleri olabileceği iddia ediliyor.
 ya da presinaptik nöronun tekrarlanan (repetitive) uyarılarıyla sağlanabilir. İkinci mekanizmada, postsinaptik membran depolarize hale getirilerek magnezyum uzaklaştırılır; ardından yeterli miktarda kalsiyum hücre içine girebilir. Bu bulgu Hebb’in 1949 yılında öne sürdüğü öngörüyü destekleyen ilk kanıttır: bir A nöronunun aksonu B nöronunu sürekli ve tekrarlayan bir şekilde uyardığında, B hücre uyarılmasının sürdürülmesini sağlamak için, bu hücrelerden birinde ya da her ikisinde bazı büyüme süreci veya metabolik değişiklikler meydana gelir. Hücreye giren kalsiyum postsinaptik uçta en az 3 protein kinazı aktive eder: Kalsiyum-kalmodulin-bağımlı protein kinaz II (CaM kinaz II olarak da bilinir) Protein kinaz C. Tirozin kinaz, fyn. Buradaki kinazlar daha önce anlatılan (3. konu) sAMP bağımlı kinazlardan farklı olmakla birlikte ona eşdeğer işler yapmaktadırlar. Örneğin CaM kinaz II, nonNMDA reseptörlerinin fosforilasyonunu sağlayarak presinaptik uçtan salıverilen glutamata duyarlılığını artırmaktadır. Ayrıca CaM kinaz II’nin intraselüler AMPA reseptörlerinin membrana translokasyonunu sağladığı gösterilmiştir. Ekstrem bir örnek olarak, bazı sinapslarda sadece NMDA reseptörleri bulunur ve bu reseptörler normal sinaptik aşırımı sağlayamazlar; yani bu sinapslar kullanılamaz durumdadırlar. Ancak LTP’den sonra postsinaptik membranda AMPA reseptörlerin ortaya çıkması ile bu sinapslar rutin sinaptik iletide rol almaya başlarlar. Normal koşullar altında tek bir CA3 nöronu sadece tek bir CA1 nöronu ile sadece tek bir sinaptik bağlantı yapar. Bu tek sinaptik bağlantıda da sadece 1 tane aktif zon yer alır ve buradan sadece tek 1 vezikül salıverilir. Bu vezikülün içinde 5000 adet glutamat molekülü bulunmaktadır. LTP’nin oluşmasından önce bu sinaptik bağlantılar etkisiz durumdadır. Presinaptik nöronda oluşan çoğu aksiyon potansiyeli bu 1 adet vezikülü salmakta bile başarısız olmaktadır; postsinaptik membranda sinaptik bir potansiyel oluşturamamaktadır. Ancak, LTP’den sonra aksiyon potansiyellerinin çoğu, bu 1 tek vezikülü salıverebilmekte ve sinaptik potansiyel oluşturabilmektedir. Böylece LTP, aktivitenin bir sinapsın gücünü nasıl artırdığına canlı bir örnektir. Burada, postsinaptik membranda üretilen bir maddenin presinaptik uca difüze olarak orayı etkilediği anlaşılmaktadır. Bu maddenin NO olabileceğine ilişkin bilgilerimiz klasik kitaplarda bulunmaktadır (HK). İlk başlarda bu maddenin NO olabileceği düşünülmekte iken şimdilerde hücre adezyon molekülleri olabileceği iddia ediliyor.")
107
A model for the induction of the early phase of long-term potentiation
A model for the induction of the early phase of long-term potentiation. According to this model NMDA and non-NMDA receptor-channels are located near each other in dendritic spines. A. During normal, low-frequency synaptic transmission glutamate (Glu) is released from the presynaptic terminal and acts on both the NMDA and non-NMDA receptors. The non-NMDA receptors here are the AMPA type. Na+ and K+ flow through the non-NMDA channels but not through the NMDA channels, owing to Mg2+ blockage of this channel at the resting level of membrane potential. B. When the postsynaptic membrane is depolarized by the actions of the non-NMDA receptor-channels, as occurs during a high-frequency tetanus that induces LTP, the depolarization relieves the Mg2+ blockage of the NMDA channel. This allows Ca2+ to flow through the NMDA channel. The resulting rise in Ca2+ in the dendritic spine triggers calcium-dependent kinases (Ca2+/calmodulin kinase and protein kinase C) and the tyrosine kinase Fyn that together induce LTP. The Ca2+/calmodulin kinase phosphorylates non-NMDA receptor-channels and increases their sensitivity to glutamate thereby also activating some otherwise silent receptor channels. These changes give rise to a postsynaptic contribution for the maintenance of LTP. In addition, once LTP is induced, the postsynaptic cell is thought to release (in ways that are still not understood) a set of retrograde messengers, one of which is thought to be nitric oxide, that act on protein kinases in the presynaptic terminal to initiate an enhancement of transmitter release that contributes to LTP.
 is released from the presynaptic terminal and acts on both the NMDA and non-NMDA receptors. The non-NMDA receptors here are the AMPA type. Na+ and K+ flow through the non-NMDA channels but not through the NMDA channels, owing to Mg2+ blockage of this channel at the resting level of membrane potential. B. When the postsynaptic membrane is depolarized by the actions of the non-NMDA receptor-channels, as occurs during a high-frequency tetanus that induces LTP, the depolarization relieves the Mg2+ blockage of the NMDA channel. This allows Ca2+ to flow through the NMDA channel. The resulting rise in Ca2+ in the dendritic spine triggers calcium-dependent kinases (Ca2+/calmodulin kinase and protein kinase C) and the tyrosine kinase Fyn that together induce LTP. The Ca2+/calmodulin kinase phosphorylates non-NMDA receptor-channels and increases their sensitivity to glutamate thereby also activating some otherwise silent receptor channels. These changes give rise to a postsynaptic contribution for the maintenance of LTP. In addition, once LTP is induced, the postsynaptic cell is thought to release (in ways that are still not understood) a set of retrograde messengers, one of which is thought to be nitric oxide, that act on protein kinases in the presynaptic terminal to initiate an enhancement of transmitter release that contributes to LTP.")
108
A model for the early and late phase of LTP
A model for the early and late phase of LTP. A single train of action potentials leads to early LTP by activating NMDA receptors, Ca2+ influx into the postsynaptic cell, and a set of second messengers. With repeated trains the Ca2+ influx also recruits an adenylyl cyclase, which activates the cAMP-dependent protein kinase (cAMP kinase) leading to its translocation to the nucleus, where it phosphorylates the CREB protein. CREB in turn activates targets that are thought to lead to structural changes. Mutations in mice that block PKA or CREB reduce or eliminate the late phase of LTP. The adenylyl cyclase can also be modulated by dopaminergic and perhaps other modulatory inputs. BDNF = brain-derived neurotrophic factor; C/EBPβ = transcription factor; P = phosphate; R(AB) dominant negative PKA; tPA tissue plasminogen activator. A model for the early and late phase of LTP. A single train of action potentials leads to early LTP by activating NMDA receptors, Ca2+ influx into the postsynaptic cell, and a set of second messengers. With repeated trains the Ca2+ influx also recruits an adenylyl cyclase, which activates the cAMP-dependent protein kinase (cAMP kinase) leading to its translocation to the nucleus, where it phosphorylates the CREB protein. CREB in turn activates targets that are thought to lead to structural changes. Mutations in mice that block PKA or CREB reduce or eliminate the late phase of LTP. The adenylyl cyclase can also be modulated by dopaminergic and perhaps other modulatory inputs. BDNF = brain-derived neurotrophic factor; C/EBPβ = transcription factor; P = phosphate; R(AB) dominant negative PKA; tPA tissue plasminogen activator.
 leading to its translocation to the nucleus, where it phosphorylates the CREB protein. CREB in turn activates targets that are thought to lead to structural changes. Mutations in mice that block PKA or CREB reduce or eliminate the late phase of LTP. The adenylyl cyclase can also be modulated by dopaminergic and perhaps other modulatory inputs. BDNF = brain-derived neurotrophic factor; C/EBPβ = transcription factor; P = phosphate; R(AB) dominant negative PKA; tPA tissue plasminogen activator. A model for the early and late phase of LTP. A single train of action potentials leads to early LTP by activating NMDA receptors, Ca2+ influx into the postsynaptic cell, and a set of second messengers. With repeated trains the Ca2+ influx also recruits an adenylyl cyclase, which activates the cAMP-dependent protein kinase (cAMP kinase) leading to its translocation to the nucleus, where it phosphorylates the CREB protein. CREB in turn activates targets that are thought to lead to structural changes. Mutations in mice that block PKA or CREB reduce or eliminate the late phase of LTP. The adenylyl cyclase can also be modulated by dopaminergic and perhaps other modulatory inputs. BDNF = brain-derived neurotrophic factor; C/EBPβ = transcription factor; P = phosphate; R(AB) dominant negative PKA; tPA tissue plasminogen activator.")
109
Mechanisms of brain plasticity
Mechanisms of brain plasticity. Collateral sprouting refers to the development of new connections from remaining nerves to the denervated region. Unmasking occurs when anatomically established but previously latent pathways and synapses are called upon to substitute for lost inputs.

110
P Maddesi

111
P Maddesi Primer duyusal nöronların (C ve Ad liflerin) akson uçlarında glutamat ile birlikte bulunur. Periferden gelen nosiseptif uyarıları SSS’ye taşır.
 akson uçlarında glutamat ile birlikte bulunur. Periferden gelen nosiseptif uyarıları SSS’ye taşır.")
112
Opioid Sistem

113
Endojen opioid peptidler(Opiopeptinler)
Prekürsör polipeptidler Pro-opiomelano-kortin (POMK) Pro-enkefalin A Prodinorfin (Pro-enkefalin B) β-lipotropin β -endorfin (-endorfin, -endorfin, ACTH ve MSH da oluşur) Kısıtlı dağılım (hipofiz + hipotalamusta nükleus arcuatus çevresilimbik sistem ve periakuaduktal gri madde) reseptörlerine afinite enkefalinler (met-enkefalin, leu-enkefalin) Yaygın dağılım (ağrı iletimi ile ilgili bölgeler, amigdala, hipokampus, LC, medulla oblongata, median eminens, adrenal medulla, pankreas, GIK sinir pleksusları) reseptörlerine afinite dinorfin-A, dinorfin-B, -neoendorfin, β -neoendorfin Yaygın dağılım (Pro-enkefalin A sistemine benzerlik gösterir) reseptörlerine afinite
 Pro-enkefalin A. Prodinorfin. (Pro-enkefalin B) β-lipotropin. β -endorfin. (-endorfin, -endorfin, ACTH ve MSH da oluşur) Kısıtlı dağılım (hipofiz + hipotalamusta nükleus arcuatus çevresilimbik sistem ve periakuaduktal gri madde) reseptörlerine afinite. enkefalinler. (met-enkefalin, leu-enkefalin) Yaygın dağılım (ağrı iletimi ile ilgili bölgeler, amigdala, hipokampus, LC, medulla oblongata, median eminens, adrenal medulla, pankreas, GIK sinir pleksusları) reseptörlerine afinite. dinorfin-A, dinorfin-B, -neoendorfin, β -neoendorfin. Yaygın dağılım (Pro-enkefalin A sistemine benzerlik gösterir) reseptörlerine afinite.")
114
Endojen opioid peptidler (Opiopeptinler)
Pro-opiomelano-kortin (POMK) Pro-enkefalin A Prodinorfin (Pro-enkefalin B) β-lipotropin β –endorfin enkefalinler (met-enkefalin, leu-enkefalin) dinorfin-A, dinorfin-B, -neoendorfin, β –neoendorfin Kısıtlı dağılım (hipofiz + hipotalamusta nükleus arcuatus çevresi limbik sistem ve periakuaduktal gri madde) Yaygın dağılım (ağrı iletimi ile ilgili bölgeler, amigdala, hipokampus, LC, medulla oblongata, median eminens, adrenal medulla, pankreas, GIK sinir pleksusları) Yaygın dağılım (Pro-enkefalin A sistemine benzerlik gösterir) reseptörlerine afinite reseptörlerine afinite reseptörlerine afinite
 Pro-enkefalin A. Prodinorfin. (Pro-enkefalin B) β-lipotropin. β –endorfin. enkefalinler. (met-enkefalin, leu-enkefalin) dinorfin-A, dinorfin-B, -neoendorfin, β –neoendorfin. Kısıtlı dağılım (hipofiz + hipotalamusta nükleus arcuatus çevresi limbik sistem ve periakuaduktal gri madde) Yaygın dağılım (ağrı iletimi ile ilgili bölgeler, amigdala, hipokampus, LC, medulla oblongata, median eminens, adrenal medulla, pankreas, GIK sinir pleksusları) Yaygın dağılım (Pro-enkefalin A sistemine benzerlik gösterir) reseptörlerine afinite. reseptörlerine afinite. reseptörlerine afinite.")
117
Figure The three known families of endogenous opioid peptides arise from three large precursor polyproteins. Each of the precursor molecules gives rise to a variety of biologically active peptide fragments, about half of which are shown in this diagram. (From Fields 1987.) A. Proopiomelanocortin (POMC) is so named because it gives rise to β-endorphin, melanocyte-stimulating hormone (MSH), adrenocorticotropic hormone (ACTH), and corticotropin-like intermediate lobe peptide (CLIP). B. Proenkephalin gives rise to multiple copies of meten-kephalin (M), a leucine-enkephalin (L), and several extended enkephalins including ME-Arg-Gly-Leu, ME-Arg-Phe, and peptides E, F, and B. Peptide E is further broken down into a family of large enkephalins that appear to be the most potent analgesic fragments derived from proenkephalin. C. Prodynorphin gives rise to dynorphin (D8), which contains the LE sequence, and neoendorphin (N).
 is so named because it gives rise to β-endorphin, melanocyte-stimulating hormone (MSH), adrenocorticotropic hormone (ACTH), and corticotropin-like intermediate lobe peptide (CLIP). B. Proenkephalin gives rise to multiple copies of meten-kephalin (M), a leucine-enkephalin (L), and several extended enkephalins including ME-Arg-Gly-Leu, ME-Arg-Phe, and peptides E, F, and B. Peptide E is further broken down into a family of large enkephalins that appear to be the most potent analgesic fragments derived from proenkephalin. C. Prodynorphin gives rise to dynorphin (D8), which contains the LE sequence, and neoendorphin (N).")
119
Endojen opioid peptidler (Opiopeptinler)
Pro-opiomelano-kortin (POMK) Pro-enkefalin A Prodinorfin (Pro-enkefalin B) β-lipotropin β –endorfin enkefalinler (met-enkefalin, leu-enkefalin) dinorfin-A, dinorfin-B, -neoendorfin, β –neoendorfin Kısıtlı dağılım (hipofiz + hipotalamusta nükleus arcuatus çevresi limbik sistem ve periakuaduktal gri madde) Yaygın dağılım (ağrı iletimi ile ilgili bölgeler, amigdala, hipokampus, LC, medulla oblongata, median eminens, adrenal medulla, pankreas, GIK sinir pleksusları) Yaygın dağılım (Pro-enkefalin A sistemine benzerlik gösterir) reseptörlerine afinite reseptörlerine afinite reseptörlerine afinite
 Pro-enkefalin A. Prodinorfin. (Pro-enkefalin B) β-lipotropin. β –endorfin. enkefalinler. (met-enkefalin, leu-enkefalin) dinorfin-A, dinorfin-B, -neoendorfin, β –neoendorfin. Kısıtlı dağılım (hipofiz + hipotalamusta nükleus arcuatus çevresi limbik sistem ve periakuaduktal gri madde) Yaygın dağılım (ağrı iletimi ile ilgili bölgeler, amigdala, hipokampus, LC, medulla oblongata, median eminens, adrenal medulla, pankreas, GIK sinir pleksusları) Yaygın dağılım (Pro-enkefalin A sistemine benzerlik gösterir) reseptörlerine afinite. reseptörlerine afinite. reseptörlerine afinite.")
120
Memelilerde Endojen Opioid Ligandlar
Prekürsör Endojen peptid Amino asid sırası Pro-opiomelanokortin b-Endorfin YGGFMTSEKSQTPLVTL- FKNAIIKNAYKKGE Pro-enkefalin [Met]enkefalin [Leu]enkefalin Metorfamide YGGFM YGGFL YGGFMRF YGGFMRGL YGGFMRRV-NH2 Pro-dinorfin Dinorfin A Dinorfin A(1-8) Dinorfin B a-neoendorfin b-neoendorfin YGGFLRRIRPKLKWDNQ YGGFLRRI YGGFLRRQFKVVT YGGFLRKYPK YGGFLRKYP Pro-nosiseptin / OFQ Nosiseptin FGGFTGARKSARKLANQ Pro-endomorfin* Endomorfin-1 Endomorfin-2 YPWF-NH2 YPFF-NH2
 Dinorfin B a-neoendorfin b-neoendorfin. YGGFLRRIRPKLKWDNQ YGGFLRRI YGGFLRRQFKVVT YGGFLRKYPK YGGFLRKYP. Pro-nosiseptin / OFQ. Nosiseptin. FGGFTGARKSARKLANQ. Pro-endomorfin* Endomorfin-1 Endomorfin-2. YPWF-NH2 YPFF-NH2.")
121
Endojen Opioid Peptid Afinitesi
Opioid reseptörler üzerine opiopeptinlerin etkileri Opioid reseptörler Endojen Opioid Peptid Afinitesi Mü (µ) OP3 Endorfin enkefalin dinorfin Endomorfin I, II Delta (δ) OP1 Enkefalin endorfin ve dinorfin Kappa (κ) OP2 Dinorfin endorfin ve enkefalin ORL-1 NOP Nosiseptin (orphanin FQ) (ORL-1 µ κ δ) NOMENCLATURE OF OPIOID RECEPTORS The nomenclature for the opioid receptors remains controversial. A 1996 review and proposal for a novel nomenclature[20] based on guidelines from NC-IUPHAR has not been widely accepted by the research community. The 1996 proposal recommended replacement of the terms, μ, δ, and κ with the terms OP3, OP1, and OP2, respectively. However, in the three years or more since the publication of this recommendation, almost all papers referring to opioid receptors have continued to use the well-established Greek symbol nomenclature. Many in the field have voiced their concerns that the original Greek symbol nomenclature is now so established that introduction of an alternative nomenclature is both inappropriate and confusing. It is argued that elimination of this well-established terminology will lead to impaired access to, and reduced citation of, the large body of research literature already published on the structure and properties of opioid receptors. NC-IUPHAR reconvened its opioid receptor subcommittee in late 1999 and charged it with developing revised recommendations for the nomenclature for opioid receptors consistent with the overall guidelines of NC-IUPHAR. This chapter is a preliminary report from the reconstituted subcommittee; all recommendations in this chapter should be considered as interim while the complex issue of nomenclature for this group of receptors is discussed further at national and international meetings. Since discussion continues on a number of complex issues, our recommendations are conservative, emphasising retention of Greek symbol nomenclature. Readers are invited to send their comments to the Chairman for review by the subcommittee. Issues relating to the nomenclature for the receptor for N/OFQ are discussed below.
 OP3. Endorfin enkefalin dinorfin. Endomorfin I, II. Delta (δ) OP1. Enkefalin endorfin ve dinorfin. Kappa (κ) OP2. Dinorfin endorfin ve enkefalin. ORL-1. NOP. Nosiseptin (orphanin FQ) (ORL-1 µ κ δ) NOMENCLATURE OF OPIOID RECEPTORS The nomenclature for the opioid receptors remains controversial. A 1996 review and proposal for a novel nomenclature[20] based on guidelines from NC-IUPHAR has not been widely accepted by the research community. The 1996 proposal recommended replacement of the terms, μ, δ, and κ with the terms OP3, OP1, and OP2, respectively. However, in the three years or more since the publication of this recommendation, almost all papers referring to opioid receptors have continued to use the well-established Greek symbol nomenclature. Many in the field have voiced their concerns that the original Greek symbol nomenclature is now so established that introduction of an alternative nomenclature is both inappropriate and confusing. It is argued that elimination of this well-established terminology will lead to impaired access to, and reduced citation of, the large body of research literature already published on the structure and properties of opioid receptors. NC-IUPHAR reconvened its opioid receptor subcommittee in late 1999 and charged it with developing revised recommendations for the nomenclature for opioid receptors consistent with the overall guidelines of NC-IUPHAR. This chapter is a preliminary report from the reconstituted subcommittee; all recommendations in this chapter should be considered as interim while the complex issue of nomenclature for this group of receptors is discussed further at national and international meetings. Since discussion continues on a number of complex issues, our recommendations are conservative, emphasising retention of Greek symbol nomenclature. Readers are invited to send their comments to the Chairman for review by the subcommittee. Issues relating to the nomenclature for the receptor for N/OFQ are discussed below.")
124
Amino Asit 3 harf 1 Harf Alanine Ala A Arginine Arg R Asparagine Asn N Aspartic acid Asp D Cysteine Cys C Glutamic Acid Glu E Glutamine Gln Q Glycine Gly G Histidine His H Isoleucine Ile I Leucine Leu L Lysine Lys K Methionine Met M Phenylalanine Phe F Proline Pro P Serine Ser S Threonine Thr T Tryptophan Trp W Tyrosine Tyr Y Valine Val V Unknonwn or "other" X

126
ORL-1 (orphan) nociceptin (orphanin FQ) *spinal analjezi
OP1 Delta (δ) OP2 Kappa (κ) OP3 Mü (µ) ORL-1 (orphan) nociceptin (orphanin FQ) *spinal analjezi - nalokson tarafından bloke edilmez - pro-orphan’dan kaynaklanır Endomorfin I, II (Mü selektif) Sigma (σ): Disfori Halusinasyon Vazomotor stimulasyon Solunum stimulasyonu ***Opioid reseptörü olarak kabul edilmez
 OP2 Kappa (κ) OP3 Mü (µ) ORL-1 (orphan) nociceptin (orphanin FQ) *spinal analjezi. - nalokson tarafından bloke edilmez. - pro-orphan’dan kaynaklanır. Endomorfin I, II (Mü selektif) Sigma (σ): Disfori. Halusinasyon. Vazomotor stimulasyon. Solunum stimulasyonu. ***Opioid reseptörü olarak kabul edilmez.")
127
Opioidlerin etkilerinde reseptör sonrası mekanizmalar
Adenilil siklaz inhibisyonu: , , Ca++ girişinin azaltılması: , , K+ konduktansının artırılması: ,

128
Bazı opioid etkilerine aracılık eden reseptör tipleri
Reseptör tipi Supraspinal analjezi µ1, δ, κ Spinal analjezi µ2, δ, κ Öfori µ1 Disfori κ Sedasyon µ, κ Miyozis µ1, κ Solunum depresyonu µ2 Gastrointestinal transitin inhibisyonu µ2, κ Diürez Prolaktin salınımında artış Büyüme hormonu salınımında artış µ2, δ Asetilkolin salınımı inhibisyonu Dopamin salınımı inhibisyonu Beslenme µ, δ, κ

130
Figure 24-8 Three of the major ascending pathways that transmit nociceptive information from the spinal cord to higher centers. The spinothalamic tract is the most prominent ascending nociceptive pathway in the spinal cord. (Adapted from Willis 1985.)
.")
131
Figure The localized modulation of pain can be explained by the gate control hypothesis. This hypothesis focuses on the interaction of four classes of neurons in the dorsal horn of the spinal cord: (1) nonmyelinated nociceptive afferents (C fibers), (2) myelinated nonnociceptive afferents (Aβ fibers), (3) projection neurons, and (4) inhibitory interneurons. The projection neuron is excited by both nocicep-tive and nonnociceptive neurons and the balance of these inputs determines the intensity of pain. The inhibitory interneuron is spontaneously active and normally inhibits the projection neuron, thus reducing the intensity of pain. It is excited by the myelinated nonnociceptive afferent but inhibited albeit not directly by the nonmyelinated nociceptor. The nociceptor thus has both direct and indirect effects on the projection neuron.
 nonmyelinated nociceptive afferents (C fibers), (2) myelinated nonnociceptive afferents (Aβ fibers), (3) projection neurons, and (4) inhibitory interneurons. The projection neuron is excited by both nocicep-tive and nonnociceptive neurons and the balance of these inputs determines the intensity of pain. The inhibitory interneuron is spontaneously active and normally inhibits the projection neuron, thus reducing the intensity of pain. It is excited by the myelinated nonnociceptive afferent but inhibited albeit not directly by the nonmyelinated nociceptor. The nociceptor thus has both direct and indirect effects on the projection neuron..")
132
Figure A descending pathway regulates nociceptive relay neurons in the spinal cord. The pathway arises in the midbrain periaqueductal gray region and projects to the nucleus raphe magnus and other serotonergic nuclei (not shown), then via the dorsolateral funiculus to the dorsal horn of the spinal cord. Additional spinal projections arise from the noradrenergic cell groups in the pons and medulla and from the nucleus paragigantocellularis, which also receives input from the periaqueductal gray region. In the spinal cord these descending pathways inhibit nociceptive projection neurons through direct connections as well as through inter-neurons in the superficial layers of the dorsal horn (see Figure 24-10).
, then via the dorsolateral funiculus to the dorsal horn of the spinal cord. Additional spinal projections arise from the noradrenergic cell groups in the pons and medulla and from the nucleus paragigantocellularis, which also receives input from the periaqueductal gray region. In the spinal cord these descending pathways inhibit nociceptive projection neurons through direct connections as well as through inter-neurons in the superficial layers of the dorsal horn (see Figure 24-10)..")
133
Figure Local-circuit interneurons in the superficial dorsal horn of the spinal cord integrate descending and afferent pathways. A. Possible interactions between nociceptor afferent fibers, local interneurons, and descending fibers in the dorsal horn of the spinal cord. Nociceptive fibers terminate on second-order spinothalamic projection neurons. Local enkephalin-containing interneurons (ENK) exert both presynaptic and postsynaptic inhibitory actions at these synapses. Serotonergic and noradrenergic neurons in the brain stem activate the local opioid interneurons and also suppress the activity of spinothalamic projection neurons. B. 1. Activation of nociceptors leads to the release of glutamate and neuropeptides from sensory terminals in the superficial dorsal horn, thus depolarizing and activating projection neurons. 2. Opiates decrease the duration of the nociceptor's action potential, probably by decreased Ca2+ influx, and thus decrease the release of transmitter from primary afferent terminals. In addition, opiates hyperpolarize the membrane of the dorsal horn neurons by activating a K+ conductance. Stimulation of the nociceptor normally produces a fast excitatory postsynaptic potential in the dorsal horn neuron; opiates decrease the amplitude of the postsynaptic potential.
 exert both presynaptic and postsynaptic inhibitory actions at these synapses. Serotonergic and noradrenergic neurons in the brain stem activate the local opioid interneurons and also suppress the activity of spinothalamic projection neurons. B. 1. Activation of nociceptors leads to the release of glutamate and neuropeptides from sensory terminals in the superficial dorsal horn, thus depolarizing and activating projection neurons. 2. Opiates decrease the duration of the nociceptor s action potential, probably by decreased Ca2+ influx, and thus decrease the release of transmitter from primary afferent terminals. In addition, opiates hyperpolarize the membrane of the dorsal horn neurons by activating a K+ conductance. Stimulation of the nociceptor normally produces a fast excitatory postsynaptic potential in the dorsal horn neuron; opiates decrease the amplitude of the postsynaptic potential.")
134
Bazı opioid ilaçların opioid reseptörler üzerine etkileri
Mü (µ) Delta (δ) Kappa (κ) Morfin +++ - + Sufentanil Fentanil Levorfanol ? Buprenorfin PAg Ant Pentazosin ++ Butorfanol Nalbufin Nalorfin Nalokson Naltrekson
 Delta (δ) Kappa (κ) Morfin Sufentanil. Fentanil. Levorfanol. Buprenorfin. PAg. Ant. Pentazosin. ++ Butorfanol. Nalbufin. Nalorfin. Nalokson. Naltrekson.")
135
A. Spinal cord in transverse section, illustrating the course of the afferent fibers and the major ascending pathways. Fast-conducting pain fibers are not confined to the spinothalamic tract but are scattered diffusely in the anterolateral funiculus (see also Fig. 8-3). B. Transverse section through a cervical segment of the spinal cord, illustrating the subdivision of the gray matter into laminae according to Rexed and the entry and termination of the main sensory fibers.
. B. Transverse section through a cervical segment of the spinal cord, illustrating the subdivision of the gray matter into laminae according to Rexed and the entry and termination of the main sensory fibers..")
136
The main somatosensory pathways
The main somatosensory pathways. The spinothalamic tract (pain, thermal sense) and the posterior column–lemniscal system (touch, pressure, joint position) are shown. Offshoots from the ascending anterolateral fasciculus (spinothalamic tract) to nuclei in the medulla, pons, and mesencephalon and nuclear terminations of the tract are indicated. The cortical representation of sensation is shown grossly; it is shown more explicitly in Fig. 9-4 and discussed in Chap. 9.
 and the posterior column–lemniscal system (touch, pressure, joint position) are shown. Offshoots from the ascending anterolateral fasciculus (spinothalamic tract) to nuclei in the medulla, pons, and mesencephalon and nuclear terminations of the tract are indicated. The cortical representation of sensation is shown grossly; it is shown more explicitly in Fig. 9-4 and discussed in Chap. 9.")
137
Spinal cord showing the segmental arrangement of nerve fibers within major tracts. On the left side are indicated the "sensory modalities" that appear to be mediated by the two main ascending pathways. C, cervical; L, lumbar; S, sacral; Th, thoracic.

138
Mechanism of action of enkephalin (endorphin) and morphine in the transmission of pain impulses from the periphery to the CNS. Spinal interneurons containing enkephalin synapse with the terminals of pain fibers and inhibit the release of the presumptive transmitter, substance P. As a result, the receptor neuron in the dorsal horn receives less excitatory (pain) impulses and transmits fewer pain impulses to the brain. Morphine binds to unoccupied enkephalin receptors, mimicking the pain-suppressing effects of the endogenous opiate enkephalin.
 impulses and transmits fewer pain impulses to the brain. Morphine binds to unoccupied enkephalin receptors, mimicking the pain-suppressing effects of the endogenous opiate enkephalin..")
139
Opioids Cortex + Opioids +/- Thalamus Hypothalamus + +/- PAG + + NRM
LC + NRPG + DLF Noradrenaline 5-HT - - - Enkephalin Dorsal horn Nociceptive afferent neuron Opioids - Periphery

140
Segmental Akupunkturun Mekanizması
Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors. Medical Acupuncture. Edinburgh: Churchill Livingston, 1998: Segmental Akupunkturun Mekanizması Figure 6.1 Mechanism of segmental acupuncture. The C primary afferent polymodal nociceptor projects to substantia gelatinosa (SG) cells in the superficial dorsal horn; these generate further impulses that pass to, or perhaps disinhibit, wide dynamic range (WDR) (or convergent) cells whose axons pass up to the brain in the spinoreticular tract where they are eventually interpreted as painful. The Aδ primary afferent pinprick receptors project both to marginal cells (M), which project up to the brain in the spinothalamic tract carrying information about pinprick that will become conscious, and to enkephalinergic stalked cells (St), which can release enkephalins (ENK) that inhibit SG cells, thus preventing information generated by noxious stimulation being transmitted further. (After Thompson & Filshie 1993, derived from Bowsher 1992.) WDR: Wide Dynamic range (convergent) hücre St: Stellar (Sepet) hücre, M: Marginal hücre; SG: Substansiya jelatinosa hücresi
 cells in the superficial dorsal horn; these generate further impulses that pass to, or perhaps disinhibit, wide dynamic range (WDR) (or convergent) cells whose axons pass up to the brain in the spinoreticular tract where they are eventually interpreted as painful. The Aδ primary afferent pinprick receptors project both to marginal cells (M), which project up to the brain in the spinothalamic tract carrying information about pinprick that will become conscious, and to enkephalinergic stalked cells (St), which can release enkephalins (ENK) that inhibit SG cells, thus preventing information generated by noxious stimulation being transmitted further. (After Thompson & Filshie 1993, derived from Bowsher 1992.) WDR: Wide Dynamic range (convergent) hücre. St: Stellar (Sepet) hücre, M: Marginal hücre; SG: Substansiya jelatinosa hücresi.")
141
Akupunkturda Serotonerjik Mekanizma
WDR: Wide Dynamic Range (convergent) hücre St: Stellar hücre (sepet hücre) M: Marginal hücre SG: Substansiya jelatinosa hücresi RF: Retiküler formasyon nRM: n. rafe magnus PAG: periakuaduktal gri madde Figure 6.2 Serotonergic mechanism of acupuncture. Pinprick information is carried up from marginal cells (M) (see also Fig. 6.1) to the ventroposterior lateral thalamic nucleus, whence it is projected to the cortex and becomes conscious; but in the midbrain these axons give off a collateral branch to the periaqueductal grey matter (PAG). The PAG projects down to the nucleus raphe magnus (NRM) in the midline of the medulla oblongata, and this in turn sends serotonergic (5-HT) fibres to the stalked cells (St). The latter inhibit substantia gelatinosa cells (SG) by an enkephalinergic mechanism (ENK), and so prevent noxious information arriving in C primary afferent nociceptors from being transmitted to wide dynamic range (WDR) cells deep in the spinal grey matter, which send their axons up to the brain (reticular formation, RF). OP = opioid peptides. The PAG is also influenced by opioid endorphinergic fibres descending from the arcuate nucleus in the hypothalamus, and the hypothalamus in turn receives projections from the prefrontal cortex. (After Thompson & Filshie 1993; derived from Bowsher 1992 see Fig p 188.) Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors. Medical Acupuncture. Edinburgh: Churchill Livingston, 1998:
 hücre. St: Stellar hücre (sepet hücre) M: Marginal hücre. SG: Substansiya jelatinosa hücresi. RF: Retiküler formasyon. nRM: n. rafe magnus. PAG: periakuaduktal gri madde. Figure 6.2 Serotonergic mechanism of acupuncture. Pinprick information is carried up from marginal cells (M) (see also Fig. 6.1) to the ventroposterior lateral thalamic nucleus, whence it is projected to the cortex and becomes conscious; but in the midbrain these axons give off a collateral branch to the periaqueductal grey matter (PAG). The PAG projects down to the nucleus raphe magnus (NRM) in the midline of the medulla oblongata, and this in turn sends serotonergic (5-HT) fibres to the stalked cells (St). The latter inhibit substantia gelatinosa cells (SG) by an enkephalinergic mechanism (ENK), and so prevent noxious information arriving in C primary afferent nociceptors from being transmitted to wide dynamic range (WDR) cells deep in the spinal grey matter, which send their axons up to the brain (reticular formation, RF). OP = opioid peptides. The PAG is also influenced by opioid endorphinergic fibres descending from the arcuate nucleus in the hypothalamus, and the hypothalamus in turn receives projections from the prefrontal cortex. (After Thompson & Filshie 1993; derived from Bowsher 1992 see Fig p 188.) Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors. Medical Acupuncture. Edinburgh: Churchill Livingston, 1998:")
142
Akupunkturda Adrenerjik Mekanizma
DNIC: Diffuse noxious inhibitory controls WDR: Wide Dynamic Range (convergent) hücre TENS: Transcutaneous electrical nerve stimulation St: Stellar hücre (sepet hücre) M: Marginal hücre SG: Substansiya jelatinosa hücresi PAG: periakuaduktal gri madde RF: Retiküler formasyon nRM: n. rafe magnus nPGC: n. paragigantaoselülaris lateralis nRG: n. retikülaris gigantoselülaris R: n. retikülaris dorsalis Figure 6.3 (facing) Adrenergic mechanism of acupuncture. Marginal cells (M), activated by Aδ pinprick receptors, in addition to their projections to the ventral posterior lateral nucleus and the PAG (through the nucleus raphe magnus (NRM) and nucleus raphe gigantocellularis (NRG) cells) also send axon branches to the following: (a) Subnucleus reticularis dorsalis (R) in the caudal medulla oblongata. Descending projections from this structure bring about inhibition of noxiously generated information arriving at the spinal cord (SG) in C nociceptors. This is the DNIC mechanism (see text). (b) Nucleus paragigantocellularis lateralis (PGC), which indirectly (? via the locus coeruleus LC, see (c) below) brings about noradrenergically mediated inhibition at spinal cord level. (c) The locus coeruleus at the junction of medulla oblongata and pons. Its noradrenergic axons (NAD) are directly inhibitory to those spinal neurons with which they enter into synaptic contact. OP = opioid peptides, DCS = dorsal column stimulation. Note: The figure also includes the Aβ primary afferent tactile receptor, which projects to the dorsal column and in addition, via an interneuron to the SG cells. Thus activation of the tactile receptor sends impulses to the dorsal column and also, via the interneurone, leads to an inhibition of the SG cells, probably through the release of γ-aminobutyric acid (GABA). The latter action will prevent information generated by noxious stimulation being transmitted further; this is believed to be the principal mechanism of transcutaneous electrical nerve stimulation (TENS) (see Ch. 11). (After Thompson & Filshie 1993, derived from Bowsher 1992) DNIC Desendan ağrı inhibisyon sisteminin özel bir formu ağrı ağrıyı inhibe eder prensibine dayanır. Bu fizyolojik sistem diffuz noxious inhibitör kontrol (DNIC) olarak adlandırılır. DNIC50-51in somatotopik olarak organize edilmesi mümkün görülmüyor, çünkü ağrı bölgesinin dışında herhangi bir vücut parçasından birçok tür noxıous stimulus tarafından tetiklenebilir. Bu fenemon hayvanların yanı sıra insanlarda da araştırılmıştır İnhibisyonun derecesi ağrı oluşturan stimulus ile doğru orantılıdır. Fakat stimülasyon kesildikten sonra sadece birkaç dakika süren mekanizma endorfinerjik ve seratoninerjik nöronları kapsayan supraspin lupla ilgilidir . DNIC ile SIA arasındaki ilişki açık değildir fakat SIA da nabız ve tansiyon yükselmeleri vardır, bu değişikler DNIC aktivasyonu için gerekli değildir. Mekanizma olarak nosiseptif sistemin bir türü olarak kabul edilmiştir.Diğer tüm ağrı inputlarının blokajı ile en yoğun ağrı oluşturan bölge meydana çıkarılır. Klinik uygulamada ağrılı bölgeye bazen gerçkten ağrılı bir stimulus verilir. Örneğin boynu ağrıyan bir hastanın boynunda adele içine serum fizyolojik zerk edilmesi gibi. Bu klinik metod muhtemelen DNIC ile çok ilgili değildir, çünkü DNIC çok kısa bir süre aktivedir,ve stimulus ağrı bölgesinin dışına verilir. Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors. Medical Acupuncture. Edinburgh: Churchill Livingston, 1998:
 hücre. TENS: Transcutaneous electrical nerve stimulation. St: Stellar hücre (sepet hücre) M: Marginal hücre. SG: Substansiya jelatinosa hücresi. PAG: periakuaduktal gri madde. RF: Retiküler formasyon. nRM: n. rafe magnus. nPGC: n. paragigantaoselülaris lateralis. nRG: n. retikülaris gigantoselülaris. R: n. retikülaris dorsalis. Figure 6.3 (facing) Adrenergic mechanism of acupuncture. Marginal cells (M), activated by Aδ pinprick receptors, in addition to their projections to the ventral posterior lateral nucleus and the PAG (through the nucleus raphe magnus (NRM) and nucleus raphe gigantocellularis (NRG) cells) also send axon branches to the following: (a) Subnucleus reticularis dorsalis (R) in the caudal medulla oblongata. Descending projections from this structure bring about inhibition of noxiously generated information arriving at the spinal cord (SG) in C nociceptors. This is the DNIC mechanism (see text). (b) Nucleus paragigantocellularis lateralis (PGC), which indirectly ( via the locus coeruleus LC, see (c) below) brings about noradrenergically mediated inhibition at spinal cord level. (c) The locus coeruleus at the junction of medulla oblongata and pons. Its noradrenergic axons (NAD) are directly inhibitory to those spinal neurons with which they enter into synaptic contact. OP = opioid peptides, DCS = dorsal column stimulation. Note: The figure also includes the Aβ primary afferent tactile receptor, which projects to the dorsal column and in addition, via an interneuron to the SG cells. Thus activation of the tactile receptor sends impulses to the dorsal column and also, via the interneurone, leads to an inhibition of the SG cells, probably through the release of γ-aminobutyric acid (GABA). The latter action will prevent information generated by noxious stimulation being transmitted further; this is believed to be the principal mechanism of transcutaneous electrical nerve stimulation (TENS) (see Ch. 11). (After Thompson & Filshie 1993, derived from Bowsher 1992) DNIC Desendan ağrı inhibisyon sisteminin özel bir formu ağrı ağrıyı inhibe eder prensibine dayanır. Bu fizyolojik sistem diffuz noxious inhibitör kontrol (DNIC) olarak adlandırılır. DNIC50-51in somatotopik olarak organize edilmesi mümkün görülmüyor, çünkü ağrı bölgesinin dışında herhangi bir vücut parçasından birçok tür noxıous stimulus tarafından tetiklenebilir. Bu fenemon hayvanların yanı sıra insanlarda da araştırılmıştır İnhibisyonun derecesi ağrı oluşturan stimulus ile doğru orantılıdır. Fakat stimülasyon kesildikten sonra sadece birkaç dakika süren mekanizma endorfinerjik ve seratoninerjik nöronları kapsayan supraspin lupla ilgilidir . DNIC ile SIA arasındaki ilişki açık değildir fakat SIA da nabız ve tansiyon yükselmeleri vardır, bu değişikler DNIC aktivasyonu için gerekli değildir. Mekanizma olarak nosiseptif sistemin bir türü olarak kabul edilmiştir.Diğer tüm ağrı inputlarının blokajı ile en yoğun ağrı oluşturan bölge meydana çıkarılır. Klinik uygulamada ağrılı bölgeye bazen gerçkten ağrılı bir stimulus verilir. Örneğin boynu ağrıyan bir hastanın boynunda adele içine serum fizyolojik zerk edilmesi gibi. Bu klinik metod muhtemelen DNIC ile çok ilgili değildir, çünkü DNIC çok kısa bir süre aktivedir,ve stimulus ağrı bölgesinin dışına verilir. Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors. Medical Acupuncture. Edinburgh: Churchill Livingston, 1998:")
143
Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors
Bowsher D. Mechanisms of acupuncture. In: Filshie J, White A, editors. Medical Acupuncture. Edinburgh: Churchill Livingston, 1998:

Benzer bir sunumlar