Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Membran ve Kas Fizyolojisi

2
Hücre membranından transport ister çift katlı lipid tabakadan direkt, isterse proteinler üzerinden olsun, iki temel olaydan biriyle gerçekleşir. Difüzyon (pasif transport) Aktif transport
 Aktif transport.")
4
Membrandan Geçiş Pasif taşınım
a. Basit Difüzyon (por, kanal, yağda eriyen) b. Kolaylaştırılmış difüzyon (glikoz) Aktif taşınım a. Doğrudan enerji kaynaklı (Na-K pompası) b. İndirek enerji kaynaklı-sekonder aktif (glikoz-Na, Na-Ca değişim) Osmoz Endositoz - Eksositoz
 b. Kolaylaştırılmış difüzyon (glikoz) Aktif taşınım. a. Doğrudan enerji kaynaklı (Na-K pompası) b. İndirek enerji kaynaklı-sekonder aktif. (glikoz-Na, Na-Ca değişim) Osmoz. Endositoz - Eksositoz.")
5
PASİF DİFÜZYON Lipidde Eriyen Maddelerin Çift Katlı Lipid Tabakasından Difüzyonu: Bir maddenin çift katlı lipid tabakadaki hareket hızını belirleyen en önemli faktörlerden birisi o maddenin lipiddeki eriyebilirliğidir. Örneğin oksijen, karbon dioksid, azot ve alkol Su ve Diğer Lipidde Erimeyen Moleküllerin Protein Kanallarından Difüzyonu: Su membran lipidlerinde hemen hiç erimediği halde membrandan özellikle protein kanalları aracılığı ile rahatlıkla geçer. Molekül büyüklüğü arttıkça penetrasyonları hızla azalır.

6
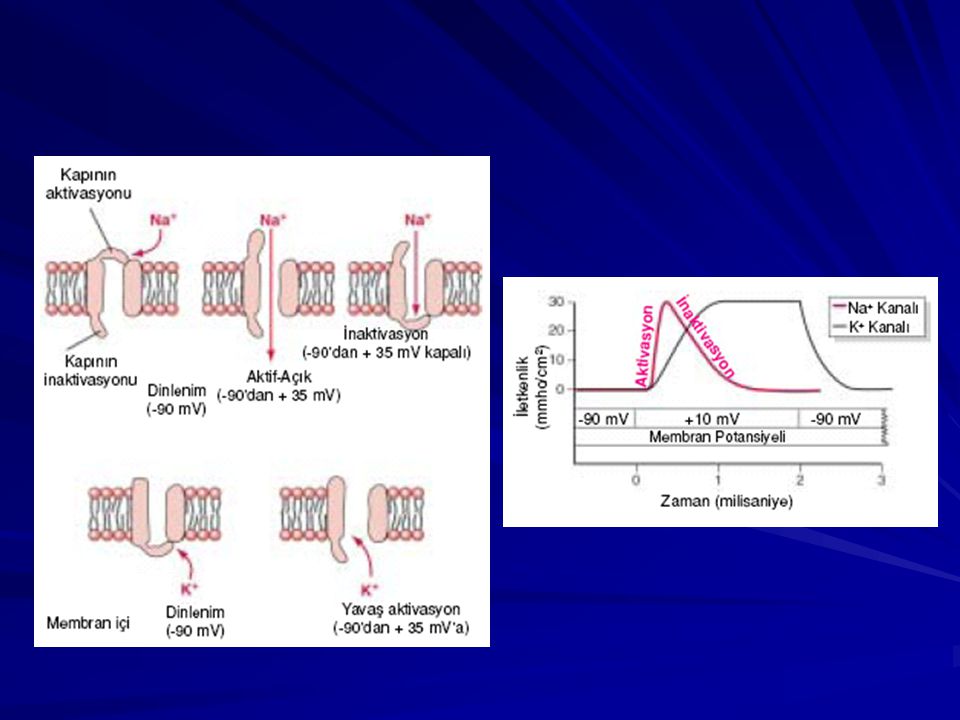
Protein Kanallardan Difüzyon ve Bu Kanalların “Kapıları”
Kapıların açılıp kapanması iki temel yolla kontrol edilir: Voltaj kapısı; Kapının moleküler konumu hücre membranının elektriksel potansiyeline bağlıdır. 2. Kimyasal kapı; Bazı protein kanal kapıları, diğer bir molekülün protein ile bağlanmasıyla açılır (asetilkolin)
")
7
KOLAYLAŞTlRILMIŞ TRANSPORT
Kolaylaştırılmış difüzyona aynı zamanda taşıyıcı aracılığı ile difüzyon da denir. Glikoz ve aminoasitlerin çoğu membrandan kolaylaştırılmış difüzyonla geçer.

8
Fark ???

9
Net difüzyon hızını etkileyen faktörler
A- Difüzyon Üzerine Konsantrasyon Farkının Etkisi. B- İyonların Difüzyonu Üzerine Elektriksel Potansiyelin Etkisi – “Nernst Eşitliği”. C- Basınç Farkının Etkisi.

10
AKTİF TRANSPORT Bir hücre membranında molekül ve iyonların konsantrasyon gradyanına (ya da elektriksel veya basınç gradyanına karşı) “yokuş yukarı” hareketine aktif transport denir. Örn sodyum, potasyum, kalsiyum, demir, hidrojen, klorür, iyodür, bazı şekerler ve aminoasitlerin çoğu
 yokuş yukarı hareketine aktif transport denir. Örn sodyum, potasyum, kalsiyum, demir, hidrojen, klorür, iyodür, bazı şekerler ve aminoasitlerin çoğu.")
11
Primer ve Sekonder Aktif Transport
Taşımada kullanılan enerjinin kaynağına göre Primer aktif transportta enerji doğrudan adenosin trifosfat (ATP) veya diğer yüksek enerji fosfat bileşiklerinin yıkılmasından üretilir. Sekonder aktif transportta enerji, birinci planda primer aktif transportla membranın iki tarafında iyonik konsantrasyon farkı şeklinde depolanmış kaynaklardan ikincil olarak üretilir
 veya diğer yüksek enerji fosfat bileşiklerinin yıkılmasından üretilir. Sekonder aktif transportta enerji, birinci planda primer aktif transportla membranın iki tarafında iyonik konsantrasyon farkı şeklinde depolanmış kaynaklardan ikincil olarak üretilir.")
12
Primer Aktif Transport
Na+-K+ Pompası Ca+2’un Primer Aktif Transportu Hidrojen İyonlarının Primer Aktif Transportu iki yerde önemlidir: 1. Midenin gastrik bezlerinde 2. Böbreklerde distal tubulusların son bölümü ve kortikal toplama kanalları pariyetal hücreler İnterkale hücrelerde

13
Sekonder Aktif Transport Birlikte Taşınma ve Zıt Taşınma
Glikoz ve Aminoasitlerin Sodyumla Birlikte Taşınması Kalsiyum ve Hidrojen İyonlarının Sodyumla Zıt Taşınması

14
OSMOZ Seçici Geçirgen Membranlarda Osmoz Suyun Net Difüzyonu
“Osmolalite” - Osmol. Bir eriyikte parçacıkların sayısal konsantrasyonlarını açıklamak için gram yerine osmol birimi kullanılır. Eğer bir kilogram suda bir osmol madde erimişse eriyiğin osmolalitesi kilogramda bir osmoldur. Ekstrasellüler ve intrasellüler sıvıların normal osmolalitesi yaklaşık olarak kilogramda 300 miliosmoldur.

15
Klatrin: - fibriler protein - aktin – miyozin
ENDOSİTOZ Klatrin: - fibriler protein - aktin – miyozin

16
Vücut sıvılarındaki iyonlar

17
Membran Potansiyelleri ve Aksiyon Potansiyelleri
SİNİRLERDE MEMBRAN DİNLENİM (İSTİRAHAT) POTANSİYELİ - Potasyum Difüzyon Potansiyelinin Katkısı - Sinir Membranından Sodyum Difüzyonunun Katkısı - Na+-K+ Pompasının Katkısı
 POTANSİYELİ. - Potasyum Difüzyon Potansiyelinin Katkısı. - Sinir Membranından Sodyum Difüzyonunun Katkısı. - Na+-K+ Pompasının Katkısı.")
18
AKSİYON POTANSiYELİ İstirahat Dönemi. Aksiyon potansiyeli oluşmadan önceki membran sükun potansiyeli. Bu dönemde büyük bir negatif (-90 mV) membran potansiyelinin bulunması nedeniyle membran “polarize” durumdadır.
 membran potansiyelinin bulunması nedeniyle membran polarize durumdadır.")
19
Depolarizasyon Dönemi
Depolarizasyon Dönemi. Bu sırada membran aniden sodyuma karşı geçirgen hale gelerek, çok büyük miktarda pozitif yüklü sodyum iyonunun aksonun içine alınmasına yol açar. Repolarizasyon Dönemi. Membranın sodyuma geçirgenliği çok arttıktan sonra, saniyenin onbinde biri gibi kısa bir sürede sodyum kanalları kapanmaya başlar ve potasyum kanalları normaldeki haline göre daha da açılır. Potasyum iyonlarının dışa doğru hızlı difüzyonu normal negatif membran dinlenim potansiyelinin yeniden oluşmasını sağlar. Buna membranın “repolarizasyonu” adı verilir. Geçirgenlik: K > Na (100 kat)
")
22
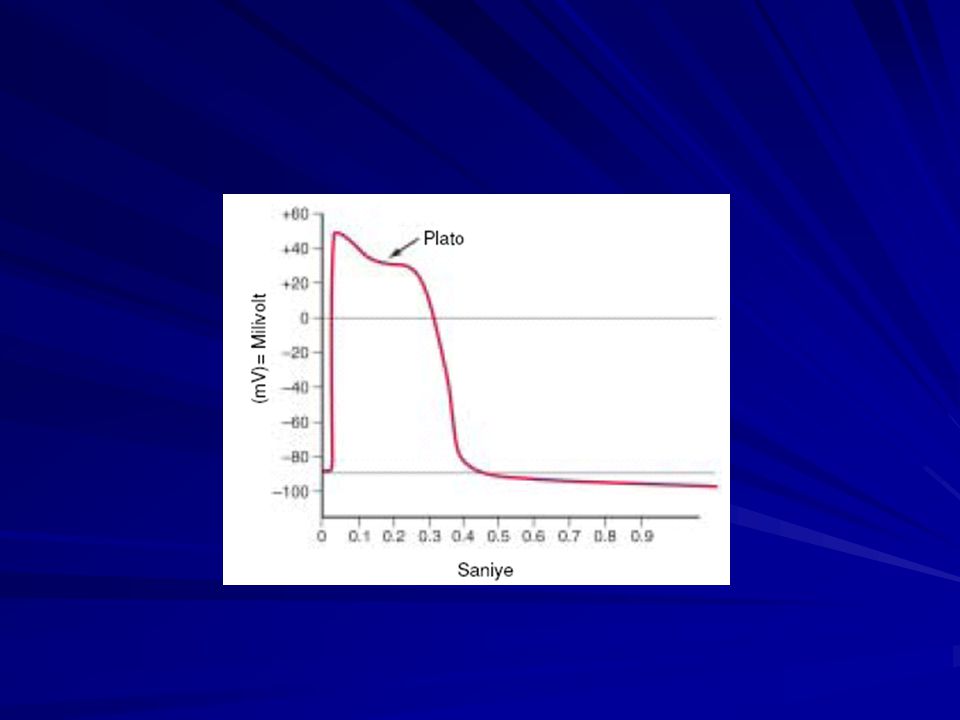
Plato Oluşumu Kalp kasında iki tip kanal depolarizasyon olayına katkıda bulunur. 1) Sodyum kanalları, hızlı kanallar, 2) Kalsiyum kanalları, yavaş kanallar. Bu kanallar, başlıca kalsiyum iyonlarıyla birlikte bir miktar sodyum iyonunun da difüzyonunu sağlar. Hızlı kanalların açılması aksiyon potansiyelinin sivri bölümünü oluşturur; ancak yavaş kalsiyum sodyum kanallarının, yavaş fakat uzun süreli açılması başlıca aksiyon potansiyelinin plato bölümünden sorumludur. Platodan kısmen sorumlu ikinci faktör ise, voltaj kapılı potasyum kanallarının çok yavaş aktive olması ve sıklıkla platonun sonuna kadar açılmamasıdır. Bu membran potansiyelinin dinlenim değerine dönmesini geciktirir. Fakat daha sonra, potasyum kanalları açılırken yavaş kanalların kapanmaya başlaması; platonun sonunda, aksiyon potansiyelinin hızla düşerek istirahat durumuna inmesini sağlar.
 Kalsiyum kanalları, yavaş kanallar. Bu kanallar, başlıca kalsiyum iyonlarıyla birlikte bir miktar sodyum iyonunun da difüzyonunu sağlar. Hızlı kanalların açılması aksiyon potansiyelinin sivri bölümünü oluşturur; ancak yavaş kalsiyum sodyum kanallarının, yavaş fakat uzun süreli açılması başlıca aksiyon potansiyelinin plato bölümünden sorumludur. Platodan kısmen sorumlu ikinci faktör ise, voltaj kapılı potasyum kanallarının çok yavaş aktive olması ve sıklıkla platonun sonuna kadar açılmamasıdır. Bu membran potansiyelinin dinlenim değerine dönmesini geciktirir. Fakat daha sonra, potasyum kanalları açılırken yavaş kanalların kapanmaya başlaması; platonun sonunda, aksiyon potansiyelinin hızla düşerek istirahat durumuna inmesini sağlar.")
23
SİNİRDE SİNYAL İLETİMİNİN ÖZEL DURUMLARI
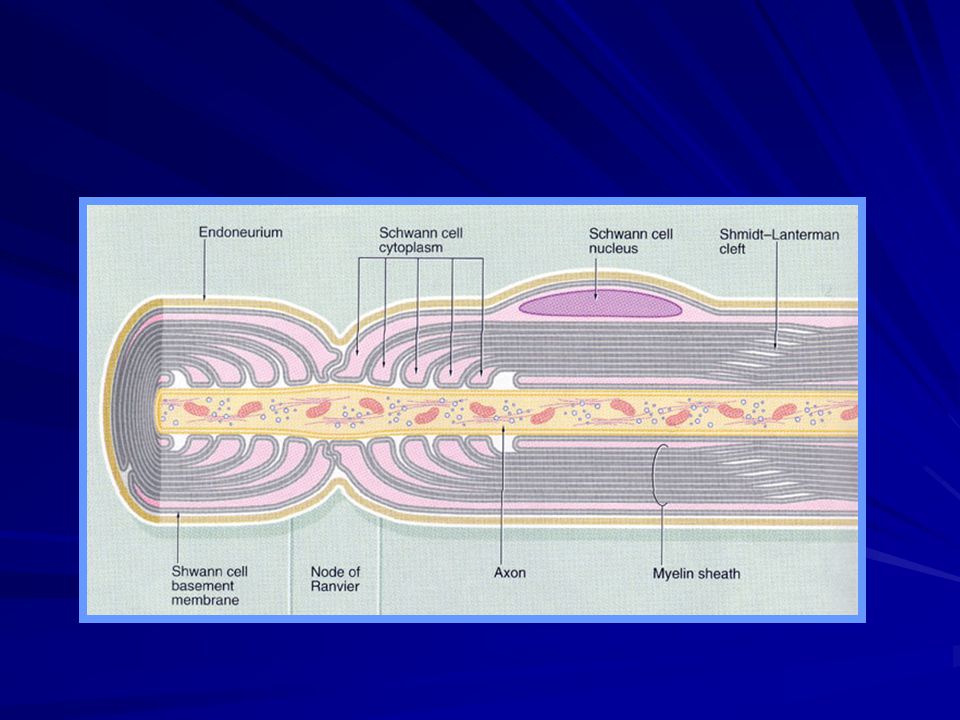
Lifin merkezinde akson bulunur ve aksonun membranı aksiyon potansiyelini ileten gerçek iletken membranıdır. Akson, viskozitesi yüksek intrasellüler bir sıvı aksoplazma ile doludur. Aksonu çeviren miyelin kılıfı çok defa aksonun kendisinden daha kalındır ve miyelin kılıfı, aksonun uzunluğu boyunca her 1-3 milimetrede bir Ranvier boğumu ile kesilmiştir.

25
Miyelinli Sİnir Liflerinde Bir Boğumdan Ötekine “Sıçrayıcı” İleti
Saltatorik ileti

26
İkinci bir aksiyon potansiyelinin çok kuvvetli bir uyaranla bile yaratılamadığı süreye mutlak refrakter periyod denir. Mutlak refrakter periyod sonrası onun dörtte biri veya yarısı kadar uzunlukta rölatif refrakter periyod gelir. Bu sürede normalden daha kuvvetli uyaran lifi uyarabilir.

27
Rölatif refrakterliğin iki nedeni vardır:
(1) Bu sırada sodyum kanallarının bazısı hala inaktivasyon durumundadır, (2) Potasyum kanallarının genellikle tam açık olması nedeniyle, pozitif yüklü potasyum iyonlarının dışarı aşırı akması lifin uyarılmasını güçleştirir.
 Bu sırada sodyum kanallarının bazısı hala inaktivasyon durumundadır, (2) Potasyum kanallarının genellikle tam açık olması nedeniyle, pozitif yüklü potasyum iyonlarının dışarı aşırı akması lifin uyarılmasını güçleştirir.")
28
“Stabilize Ediciler” ve Lokal Anestetikler
Ekstrasellüler sıvıda yüksek kalsiyum iyon konsantrasyonu, lokal anestetik olarak kullanılan prokain, tetrakain gibi maddeler direkt olarak sodyum kanallarının aktivasyon kapılarının açılmasını zorlaştırarak membranın eksitabilitesini azaltırlar.

29
İskelet Kasında Kasılma
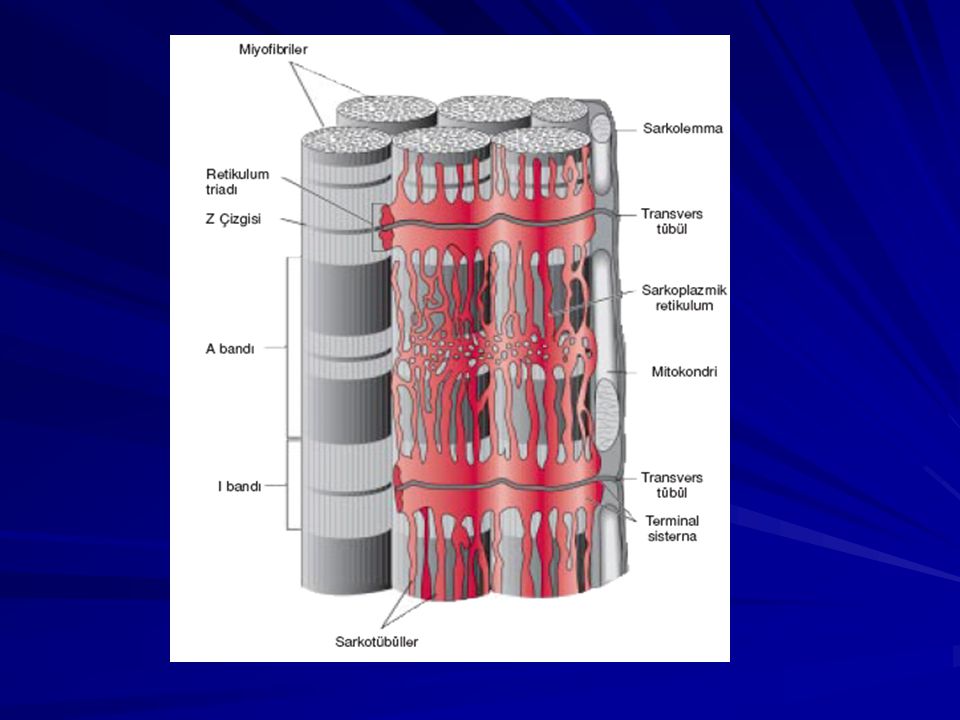
Vücudun yaklaşık %40’ı iskelet kası, %10’u düz kas ve kalp kasıdır. Sarkolemma: Kas lifinin hücre membranıdır. Sarkolemma, plazma membranı denilen gerçek hücre membranı ile birçok ince kollajen fibril içeren bir polisakkarid tabakasından meydana gelen dış kılıftan ibarettir. Miyofibriller; aktin ve miyozin filamentleri: Her kas lifi birkaç yüz ile birkaç bin arasında miyofibril içerir. Her miyofibrilde yan yana uzanan yaklaşık 1500 miyozin filamenti ve 3000 aktin filamenti vardır.

30
Kalın filamentler miyozin, ince filamentler aktin’dir.
Açık bantlar sadece aktin filamentlerini içerir ve I bandı adını alır. Koyu bantlar miyozin filamentleri ile, aralarına giren aktin filamentlerinin uçlarını içerir. Koyu bantlara A bandı denir. Miyozin filamentlerinin yan taraflarından çıkan küçük uzantılar çapraz köprülerdir. Aktin filamentlerinin ucunun tutunduğu Z diskleri Z diski, miyofibriller arasında çapraz uzanır ve kas lifi boyunca ilerleyerek bir miyofibril diğerine bağlar. (çizgili görünüm) İki Z çizgisi arasında kalan miyofibril bölümüne sarkomer denir.
 İki Z çizgisi arasında kalan miyofibril bölümüne sarkomer denir.")
31
Miyozin ve aktin moleküllerini bir arada tutan filamentöz titin molekülleridir. Miyozin ve aktin filamentleri arasındaki yan yana ilişkiyi sürdürmek zordur. Bunu, titin adı verilen çok sayıda filamentöz molekül sağlar.

32
Sarkoplazma: - Miyofibriller kas lifinde sarkoplazma denilen intraselüler maddelerden oluşan bir matriks içinde asılıdır. - Sarkoplazma sıvısı potasyum, magnezyum, fosfat ve protein yapıda enzimler içerir. - Çok sayıda mitokondri içerir (ATP)
")
33
Sarkoplazmik Retikulum:
Sarkoplazma içinde bulunan zengin endoplazmik retikuluma kas lifinde sarkoplazmik retikulum denir.

34
KAS KASILMASININ GENEL MEKANİZMASI
1. Aksiyon potansiyeli motor sinir boyunca kas lifindeki sonlanmasına kadar yayılır. 2. Her sinir ucundan nörotransmiter olarak az miktarda asetilkolin salgılanır. 3. Kas lifi membranında lokal bir alanda etki gösteren asetilkolin, membrandaki asetilkolin kapılı kanalları açar. 4. Asetilkolin kanallarının açılması, kas lifi membranından çok miktarda sodyum iyonunun içeri girmesini sağlar. Bu olay kas lifinde aksiyon potansiyelini başlatır.

35
5. Aksiyon potansiyeli sinir membranında olduğu gibi kas lifi membranı boyunca da yayılır.
6. Aksiyon potansiyeli kas lifi membranını depolarize eder ve kas lifi içine doğru yayılarak, sarkoplazmik retikulumda depolanmış olan kalsiyum iyonlarının büyük miktarlarda miyofibrile serbestlemesine neden olur. 7. Kalsiyum iyonları, filamentlerin kaymasını sağlayan, aktin ile miyozin filamentleri arasındaki çekici güçleri başlatır. 8. Sonra kalsiyum iyonları sarkoplazmik retikuluma geri pompalanır. Yeni bir kas aksiyon potansiyeli gelinceye kadar burada depolanır: kalsiyum iyonlarının uzaklaştırılması kasılmanın sona ermesine neden olur.

36
KAS KASILMASININ MOLEKÜLER MEKANİZMASI
Gevşek durumda, iki ardışık Z diskinden çıkan aktin filamentlerinin uçları, ancak birbiri üstüne binmeye başlarken, aynı zamanda miyozin filamentlerine komşu olarak uzanmaktadırlar. Kasılı durumda ise aktin filamentleri miyozin filamentleri arasında ortaya doğru çekilmiş, dolayısıyla büyük oranda üstüste binmiştir. Z diskleri de, aktin filamentleri tarafından miyozin filamentlerinin uçlarına kadar çekilmiştir.

37
Kayan filament mekanizması
Miyozin filamentlerinin çapraz köprüleri ile aktin filamenlerinin etkileşimi sonucunda mekanik bir güç oluşur. İstirahat koşullarında bu güçler inhibe edilmiştir, ancak bir aksiyon potansiyelinin kas lifi membranında yayılması sarkoplazmik retikulumdan kalsiyumun hızla serbestlemesine neden olur. Kalsiyum iyonları miyozinle aktin filamentleri arasındaki güçleri aktive eder ve kasılma başlar.

38
Kasılabilir Filamentlerin Moleküler Özellikleri
Miyozin Miyozin molekülü iki ağır zincir dört hafif zincir olmak üzere altı polipeptid zincirinden olmuştur. İki ağır zincir bir çift sarmal oluşturmak üzere birbiri etrafına spiral olarak sarılır. Miyozin molekülünün bu sarmalına kuyruk adı verilir. Bu zincirlerden herbirinin bir ucu kıvrılarak miyozin başı denen globuler polipeptid yapıyı meydana getirir.

39
Miyozin filamenti Miyozin molekülünün kuyrukları demet halinde toplanarak filamentin gövdesini oluşturmaktadır; birçok baş ise gövdeden dışarı doğru sarkmıştır. Ayrıca, her miyozin molekülünün sarmal kısmı başla beraber yana doğru uzanır ve şekilde görüldüğü gibi, başı vücuttan uzatan bir kol oluşturur. Dışarı doğru uzanan kollar ve başlara birlikte çapraz-köprü denir.

40
Miyozin Başının ATPaz Aktivitesi
Miyozin başının kas kasılması için temel olan diğer bir özelliği ATPaz enzimi olarak fonksiyon görmesidir. ATP’ı yıkmasını ve ATP’ın yüksek enerjili fosfat bağlarından elde edilen enerjiyi kasılma işlemini enerjilendirmek için kullanmasını sağlar.

41
Aktin Filamenti 3 protein bileşeninden oluşmuş bir komplekstir: aktin
tropomiyozin troponin Tropomiyozin molekülünün bir ucuna tutunmuş troponin denen bir başka protein daha vardır. Zayıf bağlı üç protein altbiriminden oluşmuş bir komplekstir. - troponin I aktin için - troponin T tropomiyozin için, - troponin C kalsiyum iyonları için kuvvetli affiniteye sahiptir. Bu kompleksin tropomiyozini aktine bağladığı düşünülür. Aktin filamenti çift-sarmal F-aktin protein molekülünden oluşur. Tropomiyozin denen farklı bir proteini de içerir. Bu moleküller F-aktin iplikleri ile zayıf bir şekilde birleşmiş ve F-aktin sarmalının kenarları etrafına spiral olarak sarılmıştır.

42
Kas kasılmasında “Boyunca yürüme” mekanizması - (I)
Kalsiyum iyonları, her biri 4 kalsiyum iyonuna kuvvetle bağlanabilen troponin C molekülleri ile birleştiğinde, troponin kompleksi biçim değişikliğine uğrar ve tropomiyozin molekülüne uyguladığı kuvvetle onu iki aktin ipliği arasındaki oluğa çeker.

43
Kas kasılmasında “Boyunca yürüme” mekanizması - (II)
Aktin filamenti kalsiyum iyonları ile aktive olur olmaz, miyozin filamentinin çapraz köprü başları aktin filamentinin aktif bölgelerine çekilir ve kasılmaya neden olur. Kasılma işlemi sırasında büyük miktarda ATP, ADP’a yıkılır. Kas tarafından yapılan iş ne kadar büyükse, yıkılan ATP miktarı da o kadar fazladır, buna Fenn etkisi denir.

45
Enerji Kaynakları yüksek enerjili fosfat bağı taşıyan fosfokreatin + ATP (5-8 saniye) kas hücrelerinde depolanmış olan glikojen (glikoliz) oksidatif metabolizma
 oksidatif metabolizma.")
46
İzotonik ve İzometrik Kasılma
Kasın kısalmadan kasılmasına izometrik, Kasın gerimi sabit kalıp kısalarak kasılmasına izotonik kasılma denir.

47
Yavaş lifler (Kırmızı)
Hızlı ve Yavaş Lifler Yavaş lifler (Kırmızı) Daha küçük lifler. Daha küçük sinir lifleri ile inervasyon. Daha fazla oksijen sağlamak için daha gelişkin kan damarı sistemi ve kapillerler. (4) Yüksek düzeydeki oksidatif metabolizmayı desteklemek için çok sayıda mitokondri. (5) Lifler bol miktarda, eritrositlerdeki hemoglobine benzer şekilde demir içeren bir protein olan miyoglobin içerir. Hızlı lifler (Beyaz) Daha büyük kasılma gücü için daha büyük lifler. Kasılmayı başlatmak üzere hızlı kalsiyum serbestlemesi için geniş bir sarkoplazmik retikulum. Glikolitik işlemle hızlı enerji sağlamak için çok miktarda glikolitik enzimler. (4) Oksidatif metabolizma ikincil önemde olduğu için daha az kan akımı. (5) Yine oksidatif metabolizma ikincil olduğundan daha az sayıda mitokondri.
 Daha küçük lifler. Daha küçük sinir lifleri ile inervasyon. Daha fazla oksijen sağlamak için daha gelişkin kan damarı sistemi ve kapillerler. (4) Yüksek düzeydeki oksidatif metabolizmayı desteklemek için çok sayıda mitokondri. (5) Lifler bol miktarda, eritrositlerdeki hemoglobine benzer şekilde demir içeren bir protein olan miyoglobin içerir. Hızlı lifler (Beyaz) Daha büyük kasılma gücü için daha büyük lifler. Kasılmayı başlatmak üzere hızlı kalsiyum serbestlemesi için geniş bir sarkoplazmik retikulum. Glikolitik işlemle hızlı enerji sağlamak için çok miktarda glikolitik enzimler. (4) Oksidatif metabolizma ikincil önemde olduğu için daha az kan akımı. (5) Yine oksidatif metabolizma ikincil olduğundan daha az sayıda mitokondri.")
48
Sumasyon, tek tek sarsıların birleşerek kasın kasılma şiddetini artırması demektir.
İki yolla meydana gelir: 1) eşzamanlı kasılan motor ünitelerin sayısını artırarak (multipl lif sumasyonu), 2) kasılma frekansını artırarak (frekans sumasyonu); bu tetanizasyona neden olabilir. Frekans belli bir kritik değere ulaştığında, ardıl kasılmalar o kadar hızlıdır ki, gerçekten birbiriyle kaynaşır ve şekilde görüldüğü gibi kasılma tamamen düzgün ve devamlı olur. Buna tetanizasyon denir.
 eşzamanlı kasılan motor ünitelerin sayısını artırarak (multipl lif sumasyonu), 2) kasılma frekansını artırarak (frekans sumasyonu); bu tetanizasyona neden olabilir. Frekans belli bir kritik değere ulaştığında, ardıl kasılmalar o kadar hızlıdır ki, gerçekten birbiriyle kaynaşır ve şekilde görüldüğü gibi kasılma tamamen düzgün ve devamlı olur. Buna tetanizasyon denir.")
49
Kas Hipertrofisi ve Atrofisi
Kas Hipertrofisi; Hemen hemen bütün kas hipertrofileri kas liflerindeki aktin ve miyozin filamentlerinin ve enerji sağlayan enzim sistemlerinin artışından kaynaklanır, buna bağlı olarak kas lifi genişler ki buna lif hipertrofisi denir. Kas uzun süre kullanılmadığı zaman kontraktil proteinlerin ve miyofibrillerin yıkılma hızı, yenilenme hızından daha fazladır. Kas atrofisi meydana gelir.

50
Rigor Mortis Ölümden birkaç saat sonra bütün vücut kasları “rigor mortis” (ölüm katılığı) denen bir kontraktür durumuna girer; bunda aksiyon potansiyeli olmadığı halde kas kasılır ve katılaşır. Bu katılık, gevşeme sırasında çapraz köprülerin aktin filamentlerinden ayrılması için gerekli olan ATP’nin tamamen kaybedilmesinden kaynaklanır.
 denen bir kontraktür durumuna girer; bunda aksiyon potansiyeli olmadığı halde kas kasılır ve katılaşır. Bu katılık, gevşeme sırasında çapraz köprülerin aktin filamentlerinden ayrılması için gerekli olan ATP’nin tamamen kaybedilmesinden kaynaklanır.")
51
UYARILARIN SİNİRDEN İSKELET KAS LİFLERİNE İLETİMİ:
SİNİR-KAS (NÖROMUSKÜLER) KAVŞAĞI
 KAVŞAĞI.")
52
Sinir lifi uç kısmında terminal dallara ayrılır ve kas lifinin içine doğru girer, fakat lifin plazma membranının dışında kalırlar. Bu yapının tamamına motor son plak denir. tek-akson terminali ile kas lifi membranı arasındaki kavşak: Membranın invajinasyonuna sinaptik oluk veya sinaptik çukur denir ve terminalle lif membranı arasındaki boşluğa sinaptik yarık denir. Oluğun tabanında subnöral yarık denen kas membranının yaptığı çok sayıda küçük kıvrım vardır.

53
Akson terminalinde bulunan çok sayıda mitokondri başlıca eksitatör bir transmiter olan asetilkolinin sentezi için gerekli enerjiyi sağlar. Asetilkolin kas lifini uyarır. Asetilkolin sinir terminalinin sitoplazmasında sentezlenir fakat hızla birçok küçük sinaptik veziküle absorbe edilir Sinaptik aralıkta bulunan çok miktarda asetilkolinesteraz enzimi asetilkolini sinaptik veziküllerden salındıktan sonra yıkar.

54
Kas membranında çok sayıda asetilkolin reseptörü görülmektedir;
bunlar gerçekte asetilkolin kapılı iyon kanallarıdır ve asetilkolin veziküllerinin sinaptik yarığa boşaltıldığı yoğun çubuk alanlarının hemen altında uzanan subnöral kıvrımların ağızlarında yerleşmişlerdir. (1) Asetilkolinin çoğu, presinaptik uç ile postsinaptik kas membranı arasındaki sinaptik boşluğu dolduran ince süngerimsi bağ dokusu tabakası olan bazal laminaya tutunmuş bulunan asetilkolinesteraz enzimi ile yıkılır. (2) Küçük bir miktar ise sinaptik alanın dışına difüze olur ve artık kas lifi membranını etkileyemez.
 Asetilkolinin çoğu, presinaptik uç ile postsinaptik kas membranı arasındaki sinaptik boşluğu dolduran ince süngerimsi bağ dokusu tabakası olan bazal. laminaya tutunmuş bulunan asetilkolinesteraz enzimi ile yıkılır. (2) Küçük bir miktar ise sinaptik alanın dışına difüze olur ve artık kas lifi membranını etkileyemez.")
55
Asetikolin kanalı A) Kapalı durumda B) Asetilkolin bağlandıktan
ve biçimsel değişiklik kanalı açıldıktan sonra, çok miktarda sodyumun (K+, Ca++) kas lifine girmesine ve kasılmayı uyarmasına izin verir. Negatif yüklü iyonların girişini engelleyen kanal ağzında negatif yükler bulunur.
 kas lifine girmesine ve kasılmayı uyarmasına izin verir. Negatif yüklü iyonların girişini engelleyen kanal ağzında negatif yükler bulunur.")
56
Sinir-Kas Kavşağında İletimi Etkileyen İlaçlar
Asetilkolin-Benzeri Etki İle Kas Lifini Uyaran İlaçlar: - Metakolin, karbakol ve nikotin gibi birçok bileşik kas lifinde asetilkoline benzer etkiye sahiptir. Bu ilaçlarla asetilkolin arasındaki fark, - kolinesteraz ile yıkılmamaları veya yavaş yıkılmaları, - etkilerinin birkaç dakikadan birkaç saate kadar sürmesidir.

57
Sinir-Kas Kavşağında İletiyi Bloke eden İlaçlar:
Kürariform ilaçlar olarak bilinen bir grup ilaç, uyarıların son plaktan kasa geçmesini engelleyebilir. D-tubokürarin kas lifi membranını, asetil kolinin etkisini asetil kolin reseptörü bölgesinde bloke ederek etkiler.

58
Asetilkolinesterazı İnaktive Ederek Sinir-Kas Kavşağını Uyaran İlaçlar
Özellikle iyi bilinen üç ilaç; neostigmin, fizostigmin ve diizopropil florofosfat, asetilkolinesterazı inaktive ederler. Sinaptik aralıkta giderek artan miktarda asetilkolin birikir. Bu durum, kasa birkaç sinir uyarısı ulaşması durumunda bile kas spazmlarına yol açar.

59
Miyastenia Gravis Sinir-kas kavşağında, sinyallerin sinir lifinden kasa iletilememesi nedeniyle paraliziye sebep olur. Miyastenia gravisli kişilerin çoğunun kanında asetilkolin kapılı taşıyıcı proteinlere karşı otoantikorlar gösterilmiştir.

60
Transvers Tübül-Sarkoplazmik
UYARILMA - KASILMA BAĞLANTISI Transvers Tübül-Sarkoplazmik Retikulum Sistemi Aksiyon potansiyellerinin kas lifi boyunca bir yandan diğer yana uzanan transvers tübüllerle (T tübülleri) taşınması sayesinde gerçekleşir. T tübülü aksiyon potansiyeli, Ca++ un sarkoplazmik retikulumdan miyofibrillerin hemen yanına serbestlemesine neden olur ve Ca++ kasılmayı sağlar. Bütün bu olaylara uyarılma-kasılma bağlantısı denir. T tübül / Z çizgisi
 taşınması sayesinde gerçekleşir. T tübülü aksiyon potansiyeli, Ca++ un sarkoplazmik retikulumdan miyofibrillerin hemen yanına serbestlemesine neden olur ve Ca++ kasılmayı sağlar. Bütün bu olaylara uyarılma-kasılma bağlantısı denir. T tübül / Z çizgisi.")
61
Sarkoplazmik Retikulumdan Kalsiyum İyonlarının Serbestlemesi
T tübülünde aksiyon potansiyeli Sinyal sisternaların membranında ve sarkoplazmik retikulumun longitudinal tübüllerinde çok sayıda kalsiyum kanalının hızla açılmasına Serbestleyen kalsiyum iyonları Troponin C’ye kuvvetle bağlanır ve kas kasılması

62
Kasta uyarılma kasılma bağlantısı, aksiyon potansiyeli ile sarkoplazmik retikulumdan kalsiyum iyonlarının serbestlemesi ve sonra kalsiyum pompası ile geri alınması.

63
Uyarılması ve Kasılması
Düz Kasın Uyarılması ve Kasılması

64
Düz Kas Tipleri Düz kaslar genelde iki büyük gruba ayrılabilir:
- Çok birimli düz kas - Üniter (tek birimli) düz kas
 düz kas.")
65
Çok Birimli Düz Kaslar Gözün silyer kasının düz kas lifleri
Çok birimli düz kas liflerinin en önemli özelliği, her lifin diğerlerinden bağımsız kasılabilmesi ve temel olarak sinir sinyalleri ile kontrol edilmeleridir. Gözün silyer kasının düz kas lifleri Gözün irisi Piloerektör kaslar

66
Üniter Düz Kaslar Sinsisyal düz kas, barsak, safra kanalları, üreter, uterus ve kan damarları birçok iç organın duvarında, bunlara viseral düz kas da denir. Lifler genellikle demet veya katlar halinde düzenlenmiştir ve hücre membranları birçok noktada birbirine bitişiktir. Hücre membranlarını birleştiren yarık bağlantılar, iyonların hücreler arasında serbestçe hareketini sağlar.

67
Düz Kas Kasılmasının Kimyasal Temeli
Düz kas, iskelet kasındaki aktin ve miyozin filamentlerine benzer kimyasal özelliklere sahip olan aktin ve miyozin filamentlerini içerir. İskelet kasındaki troponin kompleksini içermez, dolayısıyla kasılmanın kontrol mekanizması farklıdır. Kontraktil süreç kalsiyum iyonları ile aktive edilir

68
yoğun cisimleri iskelet kasındaki Z diskleri ile aynı

69
Düz Kas ile İskelet Kası Kasılmasının Karşılaştırılması
Miyozin Çapraz Köprülerinin Yavaş Döngüsü - İskelet kaslarının hızlı kasılıp gevşemesine karşın, çoğu düz kas kasılması bazen saatler veya günler süren uzun süreli tonik kasılmadır. Kasılma gücünü belirleyen temel faktör olan çapraz köprülerin aktin filamentlerine bağlı kalma süresinin, düz kasta oldukça uzadığı düşünülür. Olası bir neden çapraz köprü başlarının iskelet kasındakinden daha az ATPaz aktivitesine sahip olmasıdır. Başların hareketi için enerji sağlayan ATP yıkımı büyük miktarda azalır ve döngünün hızını da yavaşlatır. Düz Kas Kasılmasını Devam Ettirmek İçin Gerekli Enerji - Düz kasta aynı kasılma gerimini devam ettirmek için gerekli olan enerji, iskelet kasının sadece 1/10 - 1/300’üdür. - Nedeni çapraz köprü döngüsünün yavaş olması, her döngü için sadece bir ATP molekülüne ihtiyaç olunmasıdır. (enerji kullanımındaki tutumluluk)
")
70
Düz Kasta Kasılma ve Gevşeme Başlangıcının Yavaşlığı
Süre olarak iskelet kasının bir tek kasılmasının yaklaşık 30 katıdır - Kasılmanın yavaş başlaması ve uzun sürmesi çapraz köprülerin kurulması ve ayrılmasındaki yavaşlıktan kaynaklanır. Kas Kasılmasının Gücü Düz kasta miyozin filamentlerinin az olması ve çapraz köprü döngüsünün yavaş olmasına rağmen, düz kasın maksimum kasılma gücü iskelet kasınınkinden bile büyüktür.

71
“Mandal” Mekanizması Düz kas bir kez tam kasıldığında, kasın aktivasyon derecesi genellikle başlangıç seviyesinin çok altına düşürülebilir, ancak kas yine de tam kasılma gücünü sürdürebilir. Mandal mekanizmasının önemi, düz kasta az miktarda enerji kullanarak saatlerce tonik kasılmanın sürdürülebilmesini sağlamasıdır. Bunun için az miktarda sinirsel ya da hormonal uyarıya gereksinim vardır.

72
Düz Kasın Stres – Gevşemesi
Özellikle boşluklu organlarda viseral tip düz kasların uzatılma veya kısaltılmalarından sonraki saniyeler içinde hemen hemen orijinal kasılma gücüne dönebilmeleridir. Örneğin, mesanedeki sıvı hacminin aniden artmasına bağlı olarak mesane duvarı düz kasının gerilmesi, kesede aniden basınç artışına neden olur. Bununla birlikte, sonraki 15 saniye - dakika sırasında mesane duvarında gerim devam etse de, basınç hemen hemen önceki düzeyine döner. Hacim aniden düştüğü zaman basınç başlangıçta çok düşer, ancak birkaç saniye veya dakikada ilk düzeyine döner. Bu fenomene stres-gevşemesi ve ters stres gevşemesi denir.

73
Kasılma Mekaniği Kasılma intraselüler kalsiyum iyonlarındaki artış ile başlar Kalsiyum artışı düz kas lifinin sinirsel ya da hormonal yolla uyarılması, lifin gerilmesi veya lifin kimyasal çevresindeki değişikliklerden kaynaklanabilir.

74
Kalsiyum İyonlarının Kalmodulinle Birleşmesi-Miyozin Kinaz Aktivasyonu ve Miyozin Başının Fosforilasyonu Düz kas hücreleri, troponin yerine, çok miktarda kalmodulin denen bir düzenleyici protein içerirler. Bu protein dört kalsiyum iyonu ile reaksiyona girer Kalmodulin miyozin çapraz köprülerini aktive ederek kasılmayı başlatır

75
Kalsiyum iyonları kalmoduline bağlanır.
2. Kalmodulin-kalsiyum kombinasyonu, fosforile edici bir enzim olan miyozin kinazla birleşerek onu aktive eder. 3. Miyozin başının hafif zincirlerinden biri miyozin kinaza yanıt olarak fosforile olur. Bu zincir fosforile değilken, miyozin başın aktinle tutunma-ayrılma döngüsü oluşmaz. 4. Fakat regülatör zincir fosforile olduğunda, iskelet kasındaki gibi, baş aktin filamenti ile bağlanma ve kas kasılmasını sağlama kapasitesine sahiptir.

76
Kasılmanın Durması - Miyozin Fosfatazın Rolü
Kalsiyum iyon konsantrasyonu kritik bir değerin altına düştüğünde düz kas hücresi sıvısında bulunan miyozin fosfataz enzimi hafif zincirden fosfatı ayırır döngü durur ve kasılma kesilir

77
Düz kas kasılmasının sinirsel kontrolü
- Düzenli sinir-kas kavşağı yok Onun yerine, düz kası inerve eden otonom sinir lifleri genellikle bir kas lifi tabakasının üstünde diffüz olarak dallanır. İnce terminal aksonlar eksenleri boyunca dağılan birçok genişlemeler (varikozite) bulunur.
 bulunur.")
78
Lifler düz kas lifleri ile direkt temas etmezler, onun yerine yaygın kavşaklar (diffuse junctions) transmiter maddeleri birkaç nanometreden birkaç mikrometreye varan uzaklıktan düz kası çevreleyen matrikse salgılarlar Sadece asetilkolin içeren iskelet kas kavşağındaki veziküllerin tersine, otonom sinir lifi sonlanmalarındaki veziküller bazı liflerde asetilkolin, bazılarında norepinefrin ve nadiren başka maddeler içerir. Bazı reseptör proteinler eksitatörken diğerleri inhibitördür.

79
Bir dış uyaranla meydana getirilmiş tipik düz kas aksiyon potansiyeli
Düz Kasta Membran Potansiyelleri ve Aksiyon Potansiyelleri Normal istirahat halinde membran potansiyeli genellikle milivolt kadar olup, iskelet kasından yaklaşık 30 milivolt daha az negatiftir. Sivri Potansiyeller Bir dış uyaranla meydana getirilmiş tipik düz kas aksiyon potansiyeli

80
Platolu Aksiyon Potansiyelleri
Üreter, bazen uterus ve bazı damar düz kasları gibi düz kas lifi tiplerinde meydana gelen uzun süreli kasılmadan sorumlu

81
Düz kas hücre zarı, iskelet kasından çok daha fazla voltaj kapılı kalsiyum kanalına, fakat daha az voltaj kapılı sodyum kanalına sahiptir. Aksiyon potansiyeli oluşumunda sodyumun katkısı azdır. Kalsiyum kanalları sodyum kanallarından çok daha yavaş açılır, fakat ondan daha uzun süre açık kalır. Düz kas liflerinin yavaş aksiyon potansiyellerinden büyük ölçüde bu olay sorumludur.

82
Yavaş dalgaların önemi, aksiyon potansiyellerini başlatabilmeleridir
Yavaş dalgaların önemi, aksiyon potansiyellerini başlatabilmeleridir. Yavaş dalgaların kendisi kas kasılmasına neden olamaz, fakat yavaş dalganın potansiyeli yaklaşık -35 milivoltun üzerine (çoğu viseral düz kasta aksiyon potansiyeli oluşturabilecek yaklaşık eşik) çıktığı zaman aksiyon potansiyeli doğar, kas kitlesinde yayılır ve kasılma meydana gelir.
 çıktığı zaman aksiyon potansiyeli doğar, kas kitlesinde yayılır ve kasılma meydana. gelir.")
83
Viseral (üniter) düz kas yeterince gerildiğinde, genellikle spontan aksiyon potansiyelleri meydana gelir. 1) normal yavaş dalga potansiyellerinin etkilerinin, 2) gerilmeye bağlı olarak membran potansiyelinin negatifliğinde meydana gelen azalma ile birleşmesinden kaynaklanır.
 normal yavaş dalga potansiyellerinin etkilerinin, 2) gerilmeye bağlı olarak membran potansiyelinin negatifliğinde meydana gelen azalma ile birleşmesinden kaynaklanır.")
84
Lokal Doku Faktörlerine Yanıt Olarak Düz Kas Kasılması
Dokuda lokal oksijen eksikliği düz kasta gevşemeye ve dolayısıyla vazodilatasyona neden olur. 2. Aşırı karbon dioksit vazodilatasyona sebep olur 3. Hidrojen iyon konsantrasyonunun artması da vazodilatasyonu arttırır.

85
Adenozin, laktik asit, potasyum iyonlarının
Kalsiyum iyon konsantrasyonunun lokal vazodilatasyona neden olurlar

86
Kasılmayı etkileyen en önemli kan kaynaklı hormonlar
norepinefrin, epinefrin asetilkolin anjiotensin vazopresin oksitosin serotonin ve histamindir Çoğunlukla aksiyon potansiyeli olmadan kasılmayı meydana getiren Ca++ girişi ile birlikte depolarizasyon meydana gelir, İkinci haberciler siklik adenozin monofosfat (cAMP) veya siklik guanozin monofosfat (cGMP)
 veya siklik guanozin monofosfat (cGMP)")
87
Kasılmaya Neden Olan Kalsiyum İyonlarının Kaynakları:
1) Hücre membranı 2) Sarkoplazmik retikulum (iskelet kasından daha az)
 Hücre membranı. 2) Sarkoplazmik retikulum (iskelet kasından daha az)")
88
Membranın çukurcuk denen küçük
girintileri bu tübüllerin yüzeyine bitişiktir. Çukurcuklarda bir aksiyon potansiyeli yayıldığı zaman, iskelet kasında transvers tübüllerin longitudinal sarkoplazmik retikulumdan kalsiyumun serbestlemesine neden olması gibi, komşu sarkoplazmik tübüllerden kalsiyum iyonlarının serbestlemesini uyarmakta Kalsiyum iyonlarını düz kas lifinden ekstraselüler sıvıya veya sarkoplazmik retikuluma pompalayan kalsiyum pompaları (düz kas gevşemesi)
")
Benzer bir sunumlar