Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Öğr.Gör. Emine KILIÇ TOPRAK
MEMBRAN TRANSPORTU Öğr.Gör. Emine KILIÇ TOPRAK

3
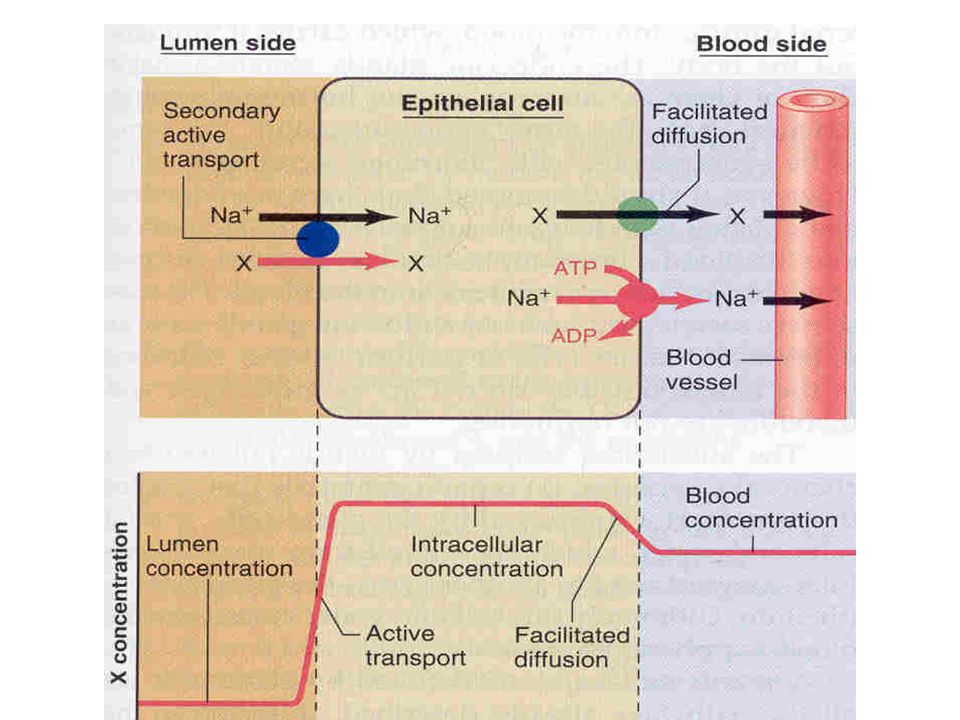
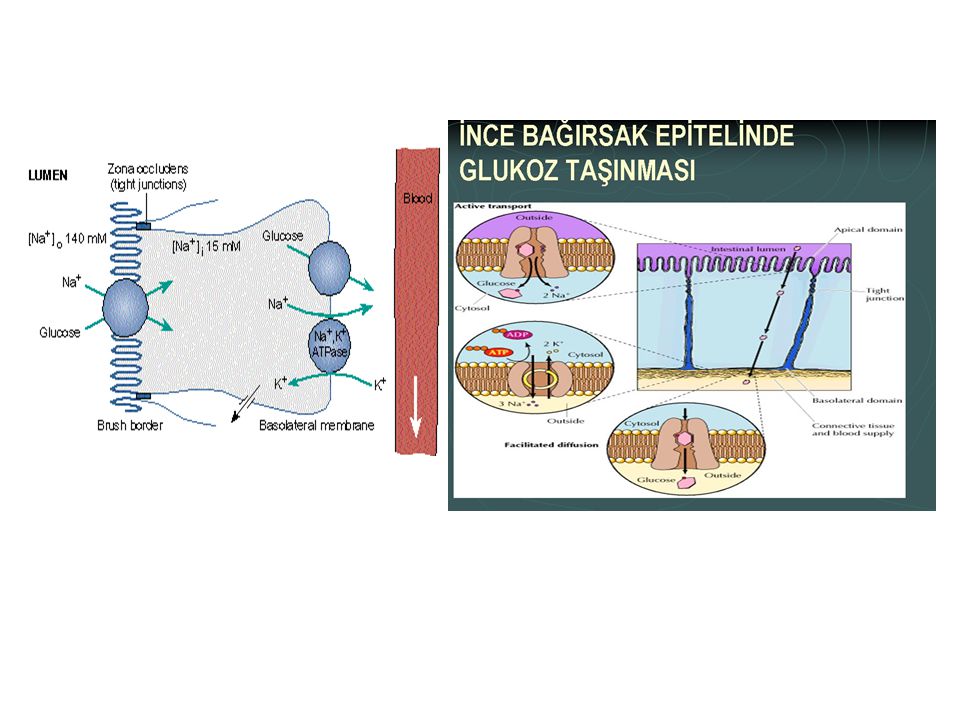
Sekonder aktif taşınmada, 2 veya daha fazla madde özel bir membran prt ile taşınır
Glukoz kimyasal gradyantla zıt yönde taşınır, doğrudan ATP kullanmasa da glikozun emilimi, bazolat membrandaki aktif Na-K-ATP-az pompasının harcadığı enerjiye bağlıdır

4
Membranın lüminal tarafında, Na iyonlarını lüminal taraftan bağlayan ve hücre içine bırakarak Na’un membrandan kolaylaştırılmış difüzyonla hücreye taşınmasını sağlayan Na taşıyıcı prtler vardır

6
PLASMA OSMOLARİTESİ HESAPLANMASI
Plasma osm. = 1.86 [Na]+[glukoz]+[BUN] 275 – 295 mOsm/kg’ dır.

7
Ozmolarite ( mOsm/L) Ozmolarite sodyum+potasyum / su ile orantılı Plazmada su miktarı fazla olursa Plazmada sodyum fazla olursa (Hipotalamusta osmoreseptörler)
")
8
HİDROSTATİK VE OSMOTİK BASINÇ

9
Arteriyel kandaki basınçlar

10
Sodyum - potasyum dengesi
Neden önemli ? Ozmolarite Uyarılabilirlik İstirahat membran potansiyeli Aksiyon potansiyeli

11
MEMBRAN POTANSİYELLERİ

12
İstirahat Membran Potansiyeli
“Yarı geçirgen bir membranın iki tarafındaki iyonların konsantrasyonu arasındaki fark, uygun koşullar altında bir membran potansiyeli yaratır. İstirahat halindeki hücrede gözlenen bu potansiyele istirahat membran potansiyeli denir.”

13
Membran Potansiyeli

14
İstirahat Membran Potansiyeli
Normalde bütün hücrelerin içi ile dışı arasında elektriksel bir potansiyel fark vardır. Sinir hücresinde -70 mv Kas hücresinde -90 mv Bu potansiyel fark hücre fonksiyonlarının başlatılmasında-tetiklenmesinde kullanılır.

15
Potansiyel fark; Na, K, Cl gibi elektrolitlerle, Hücresel proteinler,
Fosfat grupları ve Diğer nükleotidlerin hücre zarının iki tarafındaki (iç/dış) eşit olmayan dağılımından kaynaklanır.
 eşit olmayan dağılımından kaynaklanır.")
16
Hücre dışı Na mEq/lt K meq/lt Ca mEq/lt Cl mEq/lt glukoz mg/dl aminoasit 30 mg/dl Hücre içi 14 mEq/lt 140 mEq/lt <1 mEq/lt 4 mEq/lt 5 mg/dl 200 mg/dl Hücre içinde başlıca K+, Amino asitler, Mg, Fosfatlar ve proteinler fazla Hücre dışında ise; Na+, Ca+ , Cl-, Glikoz daha fazladır.

18
Membran potansiyeli Nernst potansiyeli: Membranın iki tarafı arasındaki potansiyel farkı bir iyonun membrandan net difüzyonunu önleyecek düzeydeyse o iyon için bu potansiyele verilen isimdir

19
Nernst denklemi kons iç EMF (mV)= 61 log kons dış Denge potansiyeli
Klor için: -70 mV Sodyum için: +60 mV Potasyum için: -90 mV EMF: Elektromotor Kuvvet Nernst formülünde genellikle zarın dışındaki ekstrasellüler sıvının potansiyeli sıfır olarak kabul edilir, hesaplanan Nernst potansiyeli zarın içindeki potansiyeldir.

20
K+ (potasyum) K+ hücre içinde dışına göre normalden daha fazladır.
Hücre içinden dışına çıkmak ister. Zar, sadece K+’a karşı geçirgen olsaydı, hücrenin içinden dışına doğru büyük bir K+ difüzyonu olacaktır. Böylece K+ pozitif yükleri dışarı taşıyarak zarın dış yüzünü elektropozitif yaparken, geride kalan negatif anyonlar zarın iç yüzünü negatif yapar. Difüzyonu durdurmak için gereken potansiyel fark, zarın iç yüzü negatif olmak üzere yaklaşık -90mv’tur.

21
Na+ (sodyum) Na+ hücre dışında içine göre daha fazladır.
Na+ hücre içine girme eğilimindedir. Na+ ’un hücre dışında yüksek tutulması +60 mv luk bir potansiyel fark oluşturur.

22
Bu –70 mv luk bir potansiyel fark oluşturur.
Cl– (Klor) Klor iyonlarının da hücre dışındaki miktarı hücre içinden fazladır. Ve o da hücre içine girmek ister. Bu –70 mv luk bir potansiyel fark oluşturur.
 Klor iyonlarının da hücre dışındaki miktarı hücre içinden fazladır. Ve o da hücre içine girmek ister. Bu –70 mv luk bir potansiyel fark oluşturur.")
24
Goldman-Hodgkin-Katz Denklemi
Membran birden fazla iyona geçirgen ise denge potansiyeli 3 faktöre bağlıdır: Her bir iyonun elektriksel yükünün çeşidi Membranın her iyona geçirgenliği Her bir iyonun membranın iki tarafındaki konsantrasyonu Bu gibi durumlarda membran potansiyel-denge potansiyeli Goldman-Hodgkin-Katz denklemi ile hesaplanır (-86 mv)
")
26
Goldman-Hodgkin-Katz denklemi
CNa+ iPNa+ + CK+iPK+ +CCl-dPCl- EMF = 61 log CNa+dPNa+ + CK+dPK+ +CCl-iPCl- EMF = 61 log =(-86) mv
 mv.")
27
Goldman-Hodgkin-Katz Denkleminin Önemi
1* Na, K, Cl iyonları MSS’ndeki sinir hücrelerinde olduğu gibi, sinir ve kas liflerinde zar potansiyeli gelişimine katkıda bulunan en önemli iyonlardır. 2* Voltajın hesaplanmasında herbir iyonun önem derecesi, o iyona karşı zar geçirgenliğiyle doğru orantılıdır. 3* Pozitif bir iyonun zarın içinden dışına doğru olan konsantrasyon farkı, zarın içinde elektronegatiflik yaratır. 4* Sodyum ve potasyum kanallarının geçirgenliği, sinir uyarılarının iletisi sırasında hızla değişirken, klor kanallarının geçirgenliğinde büyük bir değişim olmaz.

28
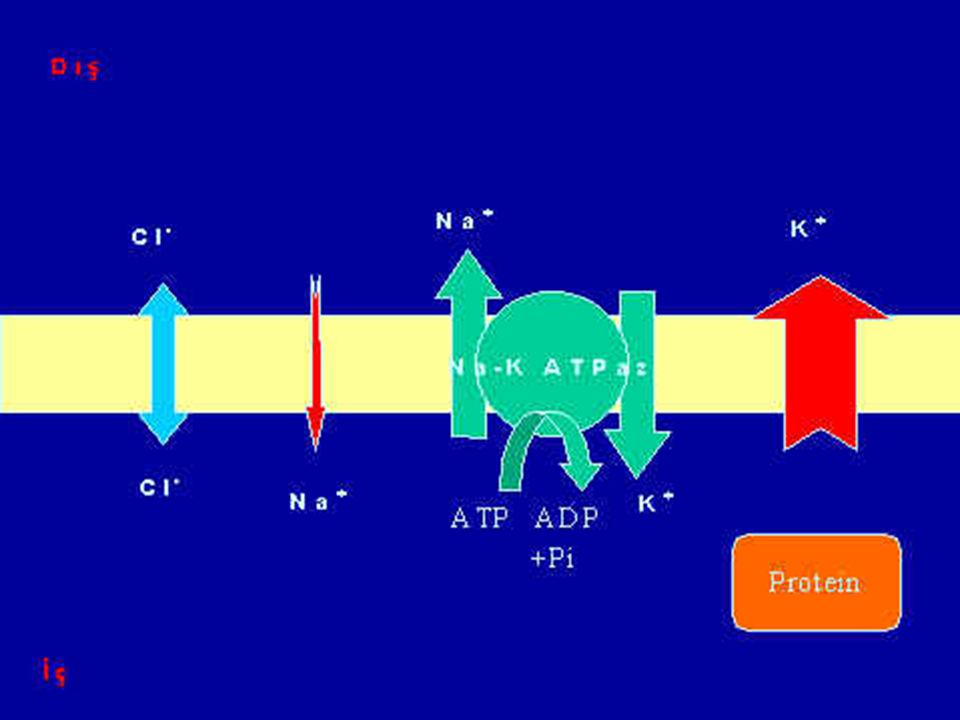
Membran dinlenim potansiyeli -90 milivolt
Na-K ATPazın etkisi (Na-K pompası) Na ve K difüzyon kanallarının etkisi (Membranın potasyuma geçirgenliğinin daha fazla olması) Hücre içi anyonların etkisi (Donnan etkisi)
 Na ve K difüzyon kanallarının etkisi (Membranın potasyuma geçirgenliğinin daha fazla olması) Hücre içi anyonların etkisi (Donnan etkisi)")
29
Potasyum sızma kanallarından K+ sızması
Zar sadece K+’a geçirgen olsaydı Nernst denklemince belirlendiği gibi potansiyel fark – 90 mv olurdu. Sodyum sızma kanallarından Na+ sızması Zar sadece Na+’a geçirgen olsaydı yine Nernst denklemine göre fark +60mv olurdu. Fakat zar Na+’a, K+ kadar geçirgen değildir (K+ 100 kat daha geçirgen) Na ve K iyonları ikisi birlikte sızdırma kanallarından oluşan geçişle –86 mv luk bir potansiyel fark oluşturur.
 Na ve K iyonları ikisi birlikte sızdırma kanallarından oluşan geçişle –86 mv luk bir potansiyel fark oluşturur.")
30
Na+ - K+ Pompası Vücuttaki bütün hücrelerin zarında bulunan bir pompadır. Görevi Na+ u hücre dışına, K+ u hücre içine pompalamaktır. Bu pompa içeriye pompaladığı her 2K+ için 3Na+ u dışarıya pompalamaktadır. Böylece devamlı olarak hücre içinde pozitif yük kaybı oluşur. (2/3) Bu durum sızma kanalları aracılığıyla oluşan difüzyonun oluşturduğu potansiyel farka ilave olarak -4 mv luk bir potansiyel fark oluşturur.
 Bu durum sızma kanalları aracılığıyla oluşan difüzyonun oluşturduğu potansiyel farka ilave olarak -4 mv luk bir potansiyel fark oluşturur.")
31
Na+ - K+ Pompası Bu pompa hücre içinde negatif bir elektriksel potansiyel oluşumunu sağlar Zarın 2 yüzü arasında Na ve K konsantrasyon farklarının korunmasına yardımcı olur Sinirsel sinyalleri ileten sinir fonksiyonlarının temelini oluşturur Hücre hacminin kontrolünü sağlar

32
İyon Geçişinde Etkili Güçler Donnan etkisinin sonuçları
Proteinler nedeniyle hücre içinde daha fazla ozmotik aktif partikül var- su alabilir Dengede iken, iyonlar asimetrik dağılımdadır; membranda elektriksel fark oluşur Donnan etkisi X Y m K+ K + Cl- Cl - Prot – K+X Cl - Y K+Y Cl - X

33
Aksiyon Potansiyeli Membran potansiyelindeki hızlı değişikliktir. Hücreler özel görevlerini aksiyon potansiyelleri olarak başlayan uyarılar ile gerçekleştirirler. Sinir sinyalleri aksiyon potansiyelleri ile iletilir. Kas kasılmaları aksiyon potansiyelleri ile başlatılır.

34
Aksiyon potansiyeli Hep yada hiç yasası…
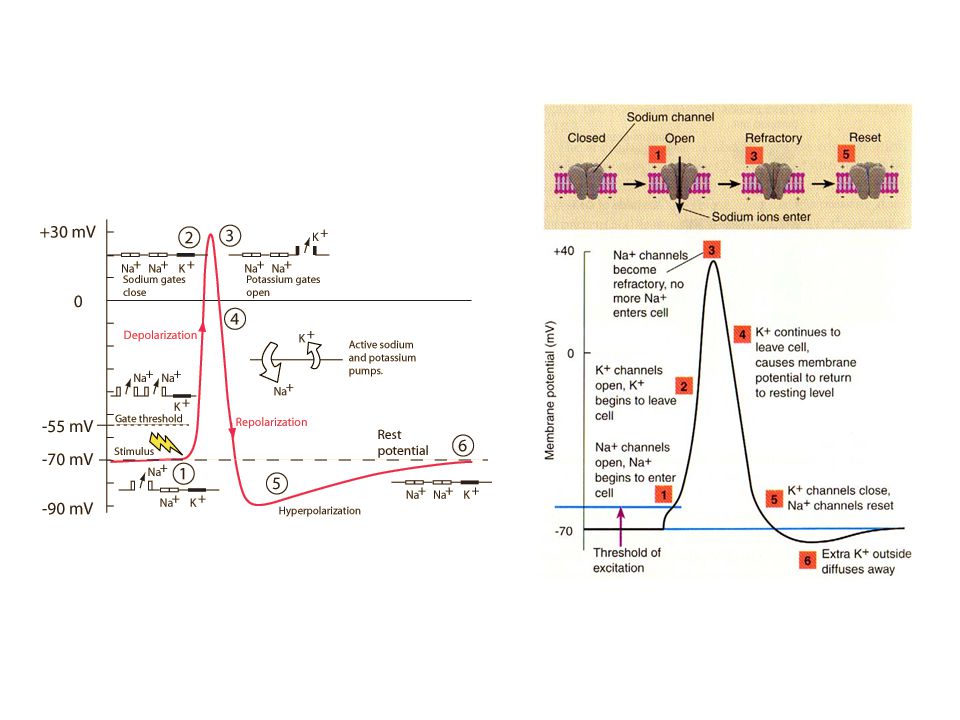
Aksiyon potansiyeli hep ya da hiç yasasına göre oluşur. Bu yasaya göre aksiyon potansiyeli ya başlar ya da başlamaz, başlarsa tüm membrana yayılır. Buna hep ya da hiç yasası denir. Bu yasa tüm uyarılabilir dokular için geçerlidir. Her aksiyon potansiyeli normal dinlenim negatif potansiyelden pozitif membran potansiyeline ani bir değişme ile başlar ve hemen hemen aynı hızla tekrar negatif potansiyele döner.

35
Aksiyon Potansiyeli Dinlenim durumu [Polarizasyon (-70,-90 mv)]
Depolarizasyon Repolarizasyon Hiperpolarizasyon
![Aksiyon Potansiyeli Dinlenim durumu [Polarizasyon (-70,-90 mv)]](http://slideplayer.biz.tr/slide/2719742/10/images/35/Aksiyon+Potansiyeli+Dinlenim+durumu+%5BPolarizasyon+%28-70%2C-90+mv%29%5D.jpg "Depolarizasyon. Repolarizasyon. Hiperpolarizasyon.")
36
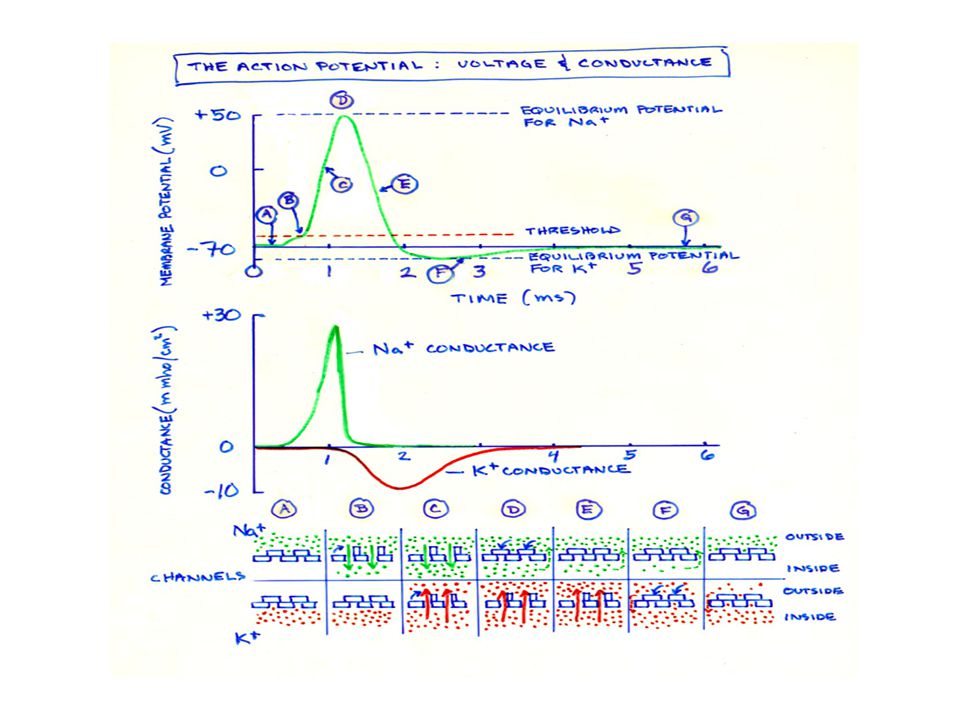
Depolarizasyon Na+ kanallarının açılıp hücre içine hızla Na+ girişi ile hücre içinin pozitifleşmesi. Depolarizasyon döneminde potansiyel fark +35 mv’a kadar çıkar.

37
Repolarizasyon Depolarizasyonda açılan Na + kanallarının kapanarak, K kanalllarının gittikçe artan hızda açılmasıyla hücre dışına K+ çıkışının olduğu dönemdir. Hücre içinden + yük (K+) kaybı ile hücre içi yeniden negatifleşerek dinlenim potansiyeli oluşur.
 kaybı ile hücre içi yeniden negatifleşerek dinlenim potansiyeli oluşur.")
38
Voltaj kapılı Na kanalları
Dinlenimde kapalı Zar potansiyeli istirahat durumuna göre daha az negatif olursa, aktivasyon kapısı aniden açılır. -90mv’dan +35mv’a aktif Voltaj daha da artarsa inaktivasyon kapısı HIZLI kapanır Dinlenim potansiyeline yakın bir potansiyelden önce kanal HIZLI açılır Voltaj kapılı K kanalı K kanal kapısı sadece zarın iç yüzeyinde yeralır ve yavaş açılır-kapanır -90 milivoltun üzerinde açılır,

40
İstirahat durumunda zarın K iletkenliği, Na iletkenliğinden 100 kat fazladır. Bunun nedeni sızma kanallarından K sızıntısının Na’dan fazla olmasıdır Aksiyon potansiyeli başladığında Na kanalları ani aktifleşir ve Na geçirgenliği 5000 kat artar

42
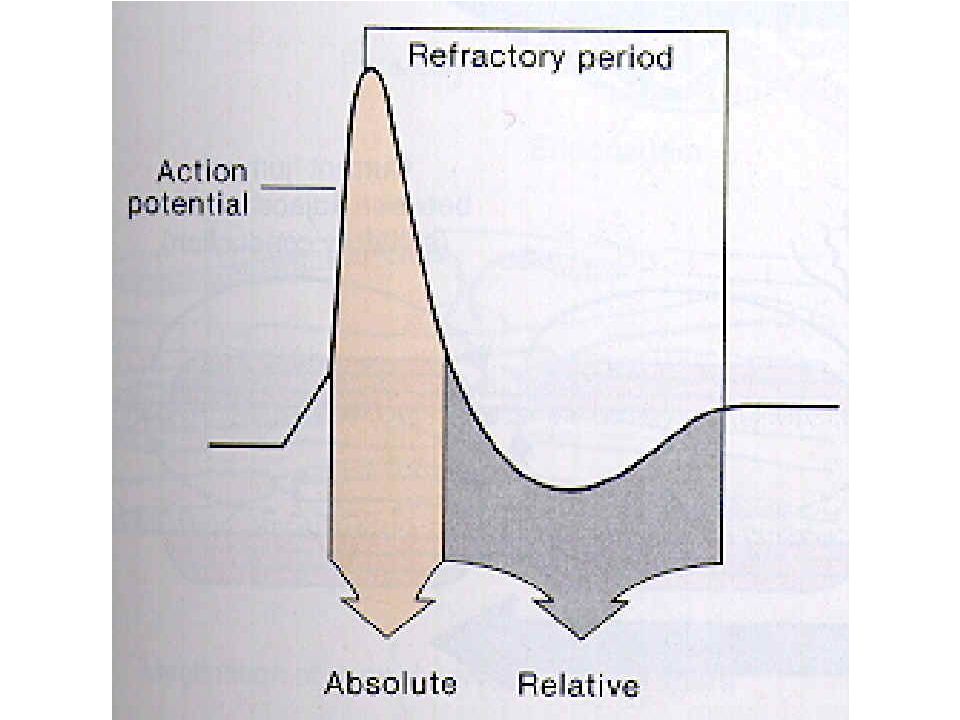
Refrakter periyod Absolute: kaslar, yüksek şiddette uyarılara bile duyarsızdır, hiçbir şekilde yanıt vermez.. Na kapıları İNAKTİF, Relative: Kas lifleri yüksek şiddetli uyarılara karşı cevap verebilir Herbir Na kanalı farklı voltajlarda AÇILIR Uzun refraktör periyod tetanik kontraksiyonların oluşmasını engeller.

44
Reobaz Kronaksi

45
Hiperpolarizasyon (ard potansiyel)
K kanal kapıları yavaş kapanır Repolarizasyon uzar

46
Aksiyon potansiyelinin iyonik temeli

47
Aksiyon potansiyelinin iyonik temeli
Bir uyarı ile voltaj değişikliği meydana gelir

48
Aksiyon potansiyelinin iyonik temeli
Voltaj bağımlı Na+ kanalları hızla, K+ kanalları yavaşça açılır

49
Aksiyon potansiyelinin iyonik temeli
Na + kanalları kapanır

50
Aksiyon potansiyelinin iyonik temeli

51
Aksiyon Potansiyelinin Yayılması
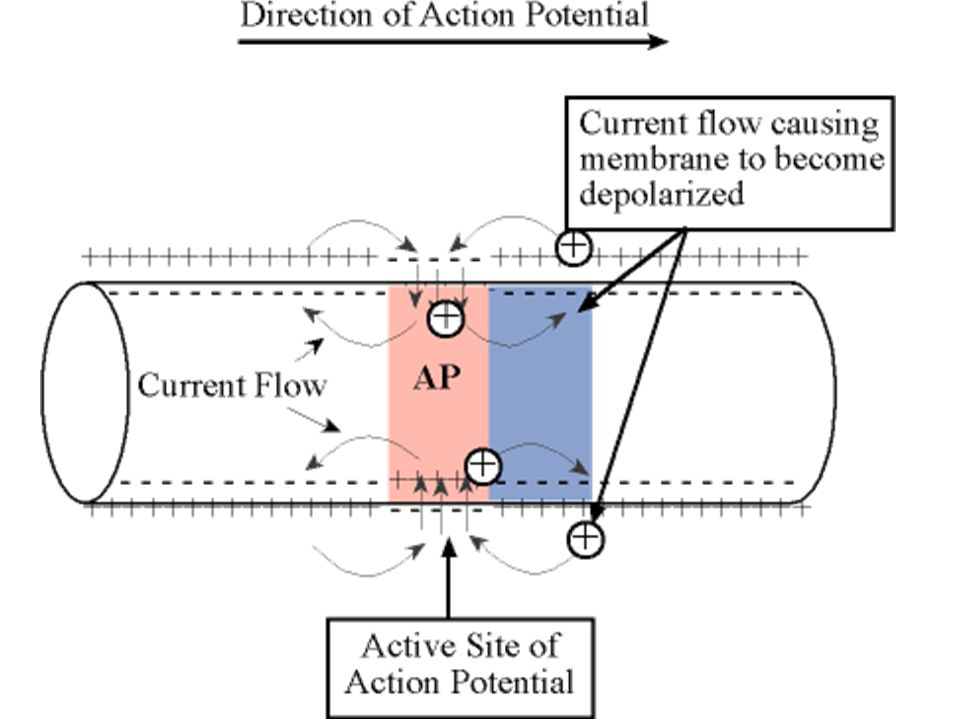
Uyarılabilen bir hücrenin (kas, sinir gibi) herhangi bir noktasında oluşan bir aksiyon potansiyeli membranın komşu bölgelerini de uyarır ve böylece aksiyon potansiyeli yayılır. Yayılma her iki yönde de oluşur.
 herhangi bir noktasında oluşan bir aksiyon potansiyeli membranın komşu bölgelerini de uyarır ve böylece aksiyon potansiyeli yayılır. Yayılma her iki yönde de oluşur.")
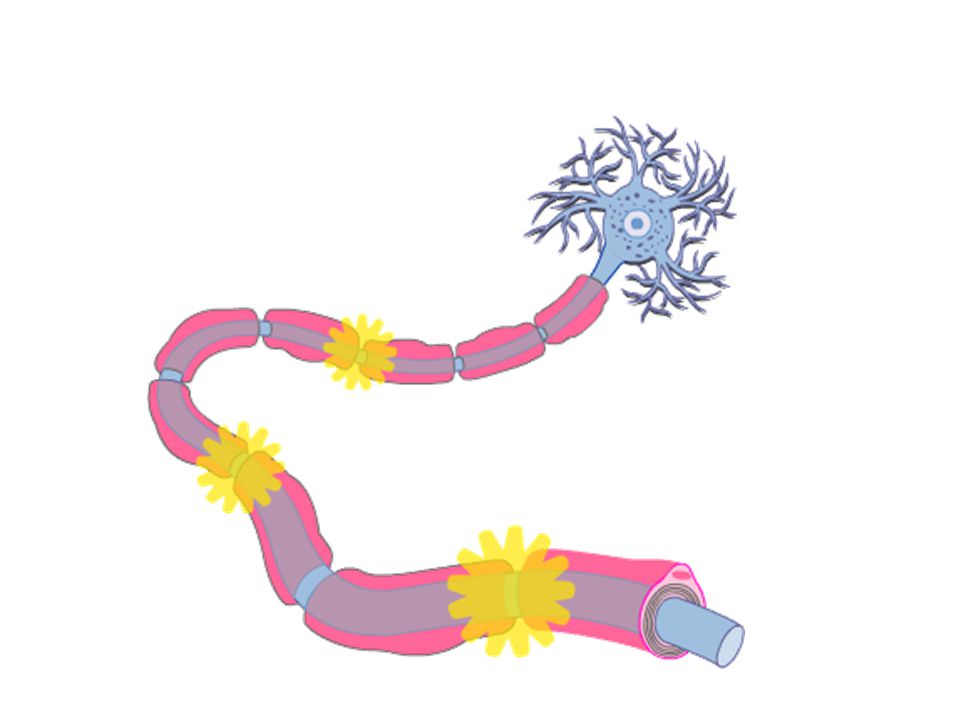
53
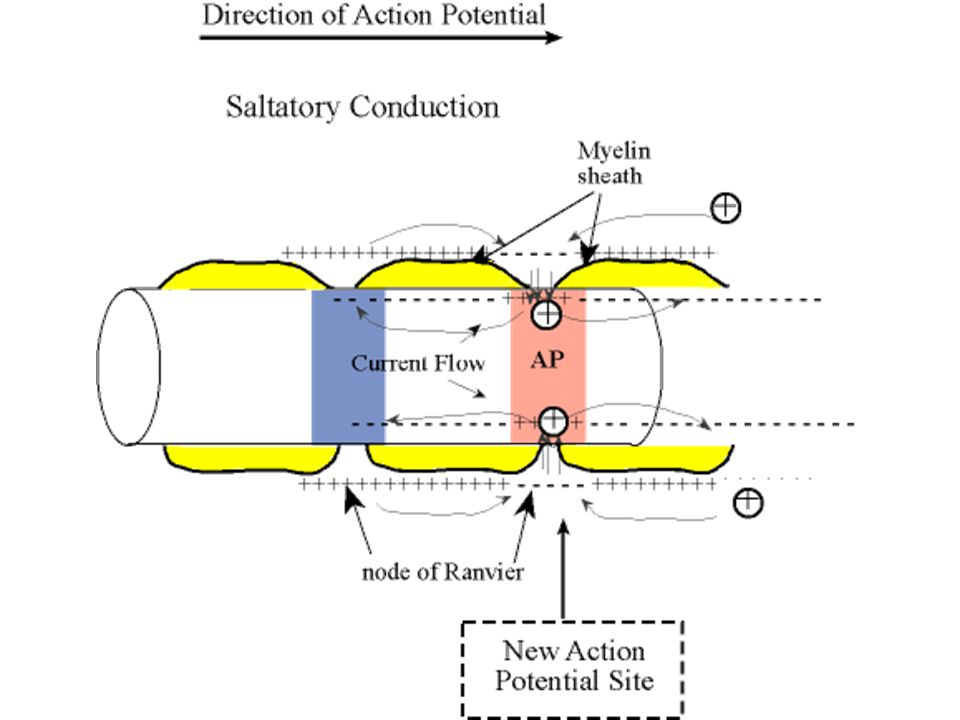
SALTATORİK İLETİ Miyelinli sinirlerin kalın miyelin kılıfından neredeyse hiçbir iyon geçmez ancak Ranvier boğumlardan kolayca geçer. Aksiyon potansiyeli sadece boğumlarda oluşabilir, sıçrayıcı şekilde sinir sinyalleri iletilir 1- Miyelinli sinirlerde ileti hızı 5-50 kat arttırılır 2- Enerji korunur, çünkü sadece boğumlarda depolarizasyon olur ve iyon kaybı 100 kat azaltılır. İleti hızı Çok küçük myelinsiz 0.25 m/sn Çok kalın myelinli 100 m/sn

54
Myelinli liflerde ileti hızı 5-50 kat fazla Enerji tasarrufu

55
İleti hızı Çok küçük myelinsiz 0.25 m/sn Çok kalın myelinli 100 m/sn

58
Fig. 8-19

59
Motor sinir ile uyarılır Uyarı organda başlar, kendiliğinden doğar.
İskelet kası Kalp kası Motor sinir ile uyarılır Uyarı organda başlar, kendiliğinden doğar. (pacemaker hücreler) Motor üniteler şeklinde kasılır Motor ünite yoktur; bütün hücreler birlikte kasılır (sinsisyum) Aksiyon potansiyeli birkaç msn sürer Aksiyon potansiyeli msn sürer Otonom sinir yok Sempatik, parasempatik sinirler
 Motor üniteler şeklinde kasılır. Motor ünite yoktur; bütün hücreler birlikte kasılır (sinsisyum) Aksiyon potansiyeli birkaç msn sürer. Aksiyon potansiyeli msn sürer. Otonom sinir yok. Sempatik, parasempatik sinirler.")
60
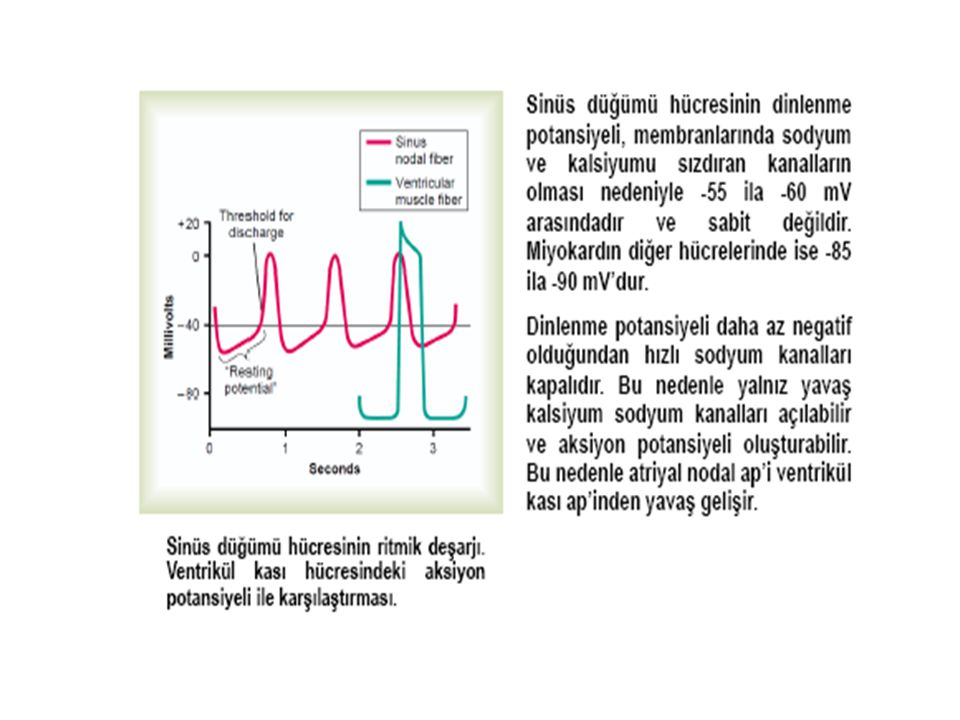
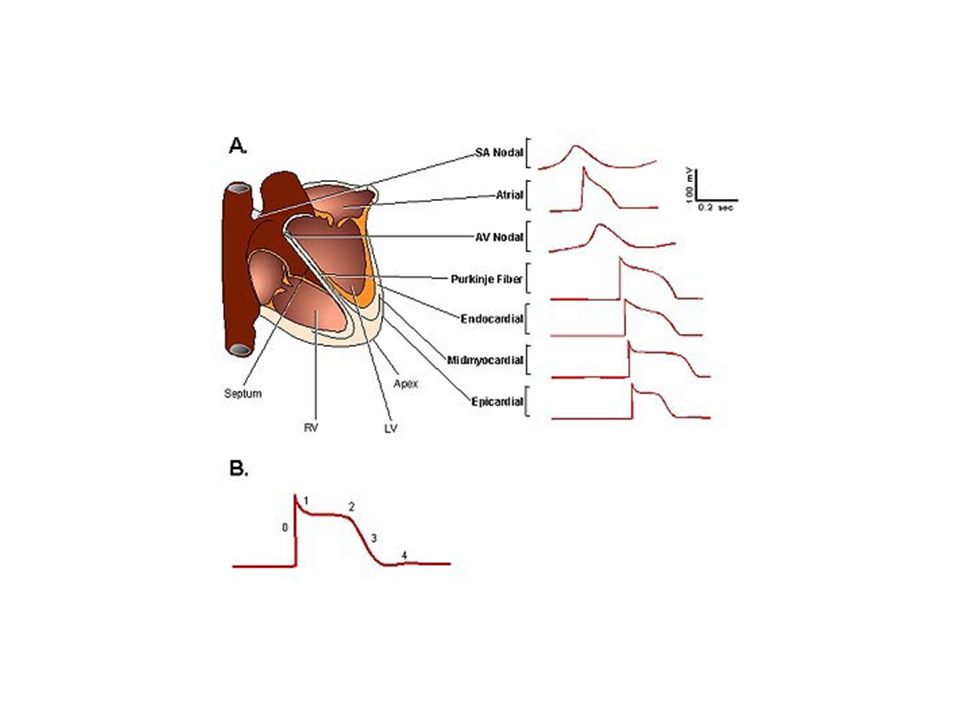
Kalpte Oluşan Aksiyon Potansiyeli 2 Tiptedir;
1- Hızlı yanıt tipi; normal atriyal ve ventriküler myositlerde ve özelleşmiş ileti liflerinde – PURKİNJE hücrelerinde- Aksiyon potansiyeli 4 esas bölümden (faz ) oluşur 2- Yavaş yanıt tipi; SA düğüm ve AV düğümde oluşur Yavaş yanıtta faz 0 çok daha az diktir faz 1 yoktur plato daha kısa sürelidir, düz değildir platodan final repolarizasyona geçiş daha az belirgindir
 oluşur. 2- Yavaş yanıt tipi; SA düğüm ve AV düğümde oluşur. Yavaş yanıtta faz 0 çok daha az diktir. faz 1 yoktur. plato daha kısa sürelidir, düz değildir. platodan final repolarizasyona geçiş daha az belirgindir.")
61
Kalp Kasında Hızlı yanıt tipiAksiyon Potansiyeli:
Faz 0: Voltaj-kapılı Na+ kanalları açılır Na+ girişi artar ve depolarizasyon gerçekleşir. Faz 1: AP pik değere ulaştığında (+20 mV), voltage-gated Na+ kanalları kapanır, K+ kanallarının açılması ile K+ çıkışı artar. Faz 2: Plato Fazı. Bu fazda hızlı K+ kanalları kapanır ve K+ çıkışı azalır, Ca2+ permeabilitesi artar. (Ca2+ kanallarının açılması ile) Faz 3: Plato fazı, Ca2+ kanallarının kapanması ve yavaş K+ kanallarının açılması ile sona erer. Faz 4: K+’un dışarı çıkması ile membran potansiyeli -90 mV düzeyine geri döner.
, voltage-gated Na+ kanalları kapanır, K+ kanallarının açılması ile K+ çıkışı artar. Faz 2: Plato Fazı. Bu fazda hızlı K+ kanalları kapanır ve K+ çıkışı azalır, Ca2+ permeabilitesi artar. (Ca2+ kanallarının açılması ile) Faz 3: Plato fazı, Ca2+ kanallarının kapanması ve yavaş K+ kanallarının açılması ile sona erer. Faz 4: K+’un dışarı çıkması ile membran potansiyeli -90 mV düzeyine geri döner.")
62
Kalp kası lifinde aksiyon potansiyeli

63
PLATOLU AKSİYON POTANSİYELİ
Kalpte, MSS’de ve düz kasların çoğunda Uyarılmış zar, depolarizasyondan sonra hemen repolarize olmaz, potansiyel birkaç sn süreyle sivri potansiyele yakın bir düzlük-plato- çizer ancak ondan sonra repolarizasyon başlar. Normal kalp kasında istirahat zar potansiyeli (–85mV),(–95mV) arasındadır. Platonun varlığı, kalp kasındaki kasılmanın, iskelet kasındakine oranla 3-15 kez daha uzun sürmesine neden olur. Kalp kası liflerinde ( sn) plato oluşumunda etkili faktörler; 1- Kalp kasında depolarizasyon olayına iki tip kanal katkıda bulunur a- voltaj bağımlı Na kanalları (hızlı), aksiyon potansiyelinin sivri bölümünü oluşturur b- voltaj bağımlı Ca-Na kanalları (yavaş), plato bölümünden sorumlu 2- Voltaj bağımlı K kanalları çok yavaş aktive olur ve platonun sonuna kadar pek fazla açılmaz.
,(–95mV) arasındadır. Platonun varlığı, kalp kasındaki kasılmanın, iskelet kasındakine oranla 3-15 kez daha uzun sürmesine neden olur. Kalp kası liflerinde ( sn) plato oluşumunda etkili faktörler; 1- Kalp kasında depolarizasyon olayına iki tip kanal katkıda bulunur. a- voltaj bağımlı Na kanalları (hızlı), aksiyon potansiyelinin sivri bölümünü oluşturur. b- voltaj bağımlı Ca-Na kanalları (yavaş), plato bölümünden sorumlu. 2- Voltaj bağımlı K kanalları çok yavaş aktive olur ve platonun sonuna kadar pek fazla açılmaz.")
64
Plato dönemi Aksiyon potansiyelinde plato varlığı, ventrikül kasılmasının kalp kasında iskelet kasındakine oranla 15 kez daha uzun sürmesine neden olur Plato dönemi, kas kontraksiyonunun ventriküllerden kanı fırlatmaya yetecek kadar uzun sürmesini sağlar. Fırlatma tamamlanmadan kas hücrelerinin tekrar uyarılmalarına izin vermez. Plato döneminde kalsiyumun hücre içine girmesi kontraksiyon sürecini başlatır.

65
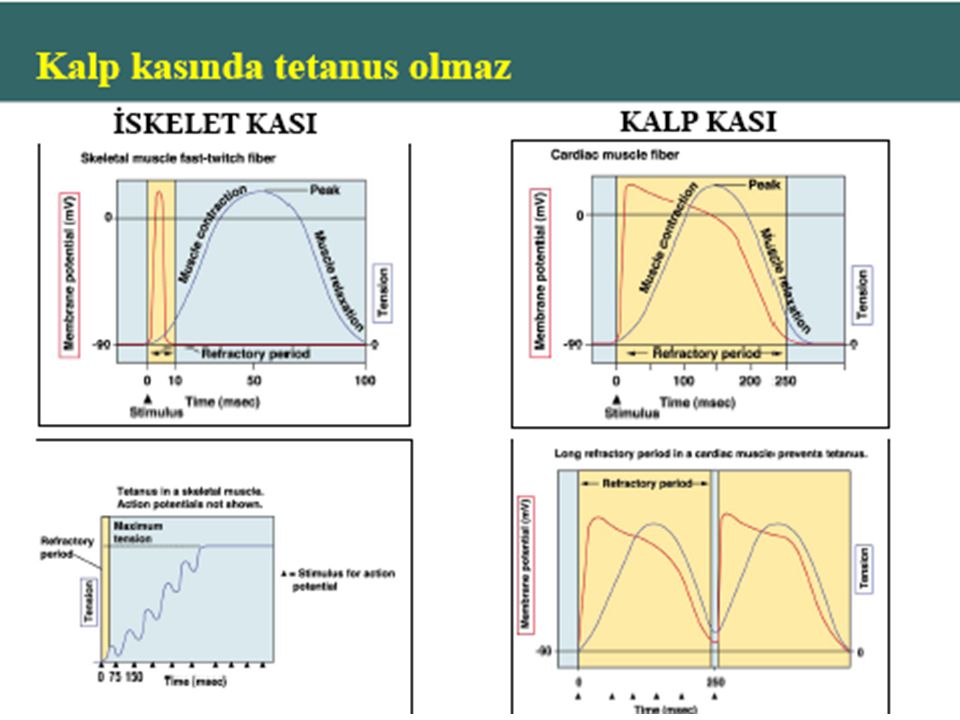
Kaslar uyarılmaya karşı (AP) kasılma yanıtı verirler.
AP tamamlanmadan hücre yeniden uyarılamaz (refrakter dönem). Kalbin AP’i bitene kadar kalp gevşemiş olur. Kalpte tetani veya summasyon görülmez.
. Kalbin AP’i bitene kadar kalp gevşemiş olur. Kalpte tetani veya summasyon görülmez.")
66
Aksiyon potansiyelinin başlamasından hemen sonra iskelet kasında gözlenmeyen bir olay meydana gelir.
Kalp kası zarının potasyum geçirgenliği yaklaşık beşte birine kadar azalır. Bu azalma aksiyon potansiyelinin platosunun oluşmasına katkıda bulunur. Yavaş kalsiyum kanalları kapanınca zarın potasyum geçirgenliği hızla artar. Lif hızla potasyum kaybedince zar potansiyeli hızla istirahat düzeyine geri döner, aksiyon potansiyeli son bulur.

67
Neden kalp kontrol merkezinin zarı repolarize olduktan hemen sonra depolarize olmuyor? Sonraki aksiyon potansiyelinin başlamasından önce yaklaşık 1 sn’lik gecikme oluyor? Her aksiyon potansiyelinin sonuna doğru zarın, kısa bir dönem için potasyuma geçirgenliği ileri derecede artar. Aşırı miktarda potasyum iyonunun dışarı çıkışı, beraberinde büyük miktarda pozitif yükleri zarın dışına taşır, böylece lifin iç yüzünde normalden daha fazla negatiflik oluşur Bir önceki aksiyon potansiyelini tamamladıktan sonra, yaklaşık 1 sn bu durum devam eder. Böylece zar potansiyeli, Nernst potansiyeline çok yakın bir değere yaklaştırılır. Bu döneme HİPERPOLARİZASYON denir. Aşırı K+ iletkenliği yavaş yavaş kaybolarak, zar potansiyelinin yeni bir uyarı için eşik değere yükselmesine izin verilir.

69
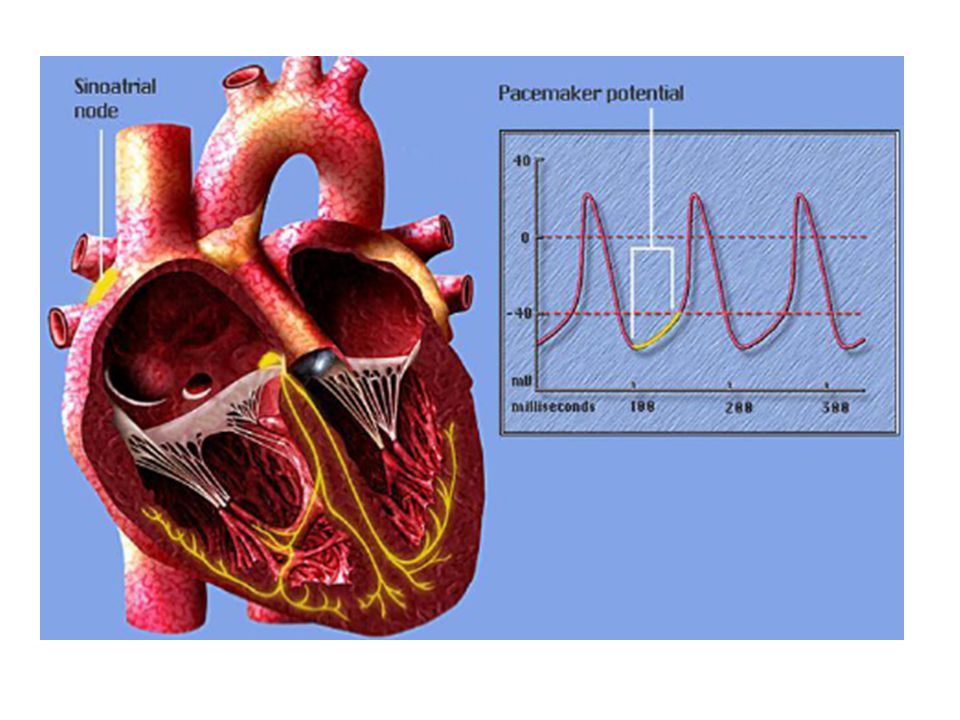
Pacemaker Hücresinde Aksiyon Potansiyeli ve İyon Kanalları
Sinüs düğümü (Sinoatrial düğüm ) Sağ atriumun süperior lateral duvarında , vena kava süperiorun ağzının hemen altında ve hafif dış tarafındadır. Bu düğümün lifleri hemen hiç kasılabilir lif içermez. Bu liflerin hücre zarları sodyum iyonunu sızdırma özelliği bulunduğundan istirahat potansiyelleri daha az negatiftir, bu da liflerden ritmik uyarıların çıkmasına neden olur.
 Sağ atriumun süperior lateral duvarında , vena kava süperiorun ağzının hemen altında ve hafif dış tarafındadır. Bu düğümün lifleri hemen hiç kasılabilir lif içermez. Bu liflerin hücre zarları sodyum iyonunu sızdırma özelliği bulunduğundan istirahat potansiyelleri daha az negatiftir, bu da liflerden ritmik uyarıların çıkmasına neden olur.")
72
Ventrikül Miyokard Hücresinde Aksiyon Potansiyeli
Kaslar uyarılmaya karşı (AP) kasılma yanıtı verirler. AP tamamlanmadan hücre yeniden uyarılamaz (refrakter dönem). Kalbin AP’i bitene kadar kalp gevşemiş olur. Kalpte tetani veya summasyon görülmez.
 kasılma yanıtı verirler. AP tamamlanmadan hücre yeniden uyarılamaz (refrakter dönem). Kalbin AP’i bitene kadar kalp gevşemiş olur. Kalpte tetani veya summasyon görülmez.")
73
Ventrikül Miyokard Hücresinde Aksiyon Potansiyeli
Depolarizasyon: Spontan (+) iyon girişi (Na+, Ca++) AP ateşlemesi (- 45 mV), Hızlı Na+ kanallarının açılması, (hücre içi voltaj yükselir) Yavaş Ca++ kanallarının açılması, K+ kanallarının açılmasının gecikmesi Repolarizasyon: K+ kanallarının açılması Hücre içi voltajın tekrar düşmesi SONUÇ: uzun ve platolu AP oluşması
 iyon girişi. (Na+, Ca++) AP ateşlemesi (- 45 mV), Hızlı Na+ kanallarının açılması, (hücre içi voltaj yükselir) Yavaş Ca++ kanallarının açılması, K+ kanallarının açılmasının gecikmesi. Repolarizasyon: K+ kanallarının açılması. Hücre içi voltajın tekrar düşmesi. SONUÇ: uzun ve platolu AP oluşması.")
75
AP-kontraksiyon ilişkisi:
AP iskelet kasında oldukça kısa sürelidir AP biter bitmez kasılma ölçülebilir AP kalp kasında uzun sürelidir AP ve kasılma birbiri üzerine binmiştir. AP bittiğinde aynı zamanda kasılma da sona erer

76
- +
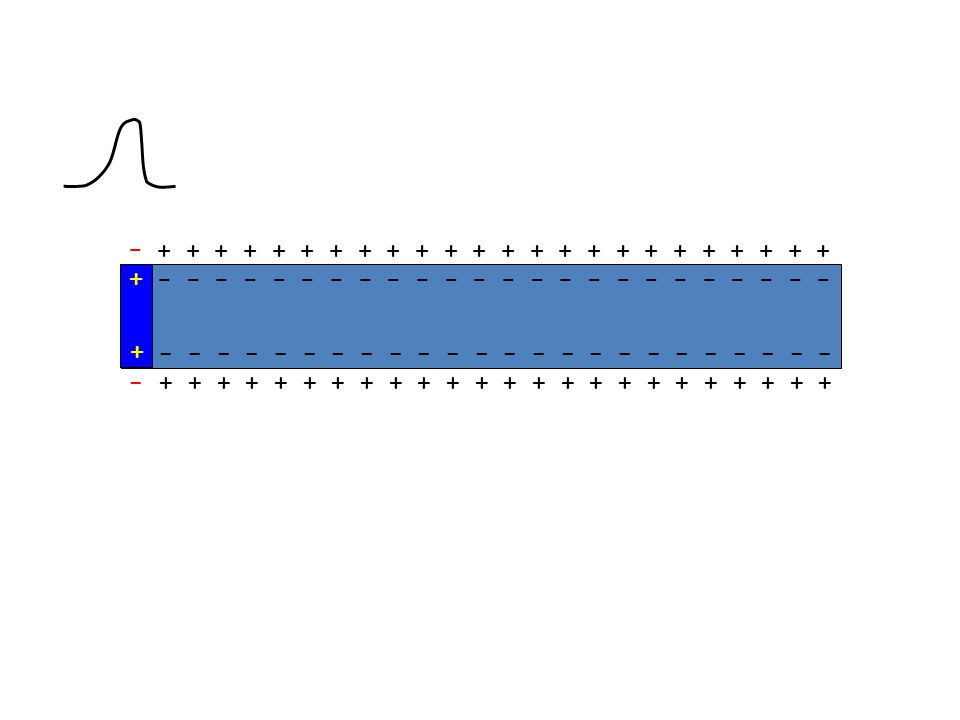
77
DÜZ KASLARIN SINIFLANDIRILMASI
1- TEK ÜNİTELİ DÜZ KASLAR (ÜNİTER- VİSSERAL) İnce barsaklar Safra kanalı Kolon Mesane Üreterler Uterus (myometriyum) Lenf damarları Küçük arterioller 2- ÇOK ÜNİTELİ DÜZ KASLAR Gözde Silyer kas İris kası Trakeal kas Bronşial kas GI sfinkterleri Piloerektör kaslar
 İnce barsaklar. Safra kanalı. Kolon. Mesane. Üreterler. Uterus (myometriyum) Lenf damarları. Küçük arterioller. 2- ÇOK ÜNİTELİ DÜZ KASLAR. Gözde Silyer kas. İris kası. Trakeal kas. Bronşial kas. GI sfinkterleri. Piloerektör kaslar.")
78
Düz kasta membran ve aksiyon potansiyelleri
Normal istirahat membran pot. -50,-60 mv ‘dur Visseral düz kasta da AP iskelet kasındaki yolla oluşur Çok birimli düz kas liflerinin çoğunda ise oluşmaz

79
Tek Üniteli (Visseral) Düz Kas Aksiyon Potansiyelleri
SİVRİ POTANİYELLER; Süresi msn olup, iskelet kasındakine benzer PLATOLU AP; AP’nin başlangıcı tipik sivri potansiyelle aynıdır. Ancak hızlı repolarizasyon görülmez, birkaç yüz-bin msn gecikir PLATO; üreter, uterus, damar düz kaslarında oluşan uzun süreli kasılmalardan sorumludur

80
Ca+2 Kanallarının Önemi
Düz kas, iskelet kasına göre daha fazla voltaj kapılı Ca+2 kanalına, daha az voltaj kapılı Na+ kanalına sahiptir. Bu nedenle AP’den primer sorumlu Ca+2 kanalıdır, çok daha yavaş açılır

81
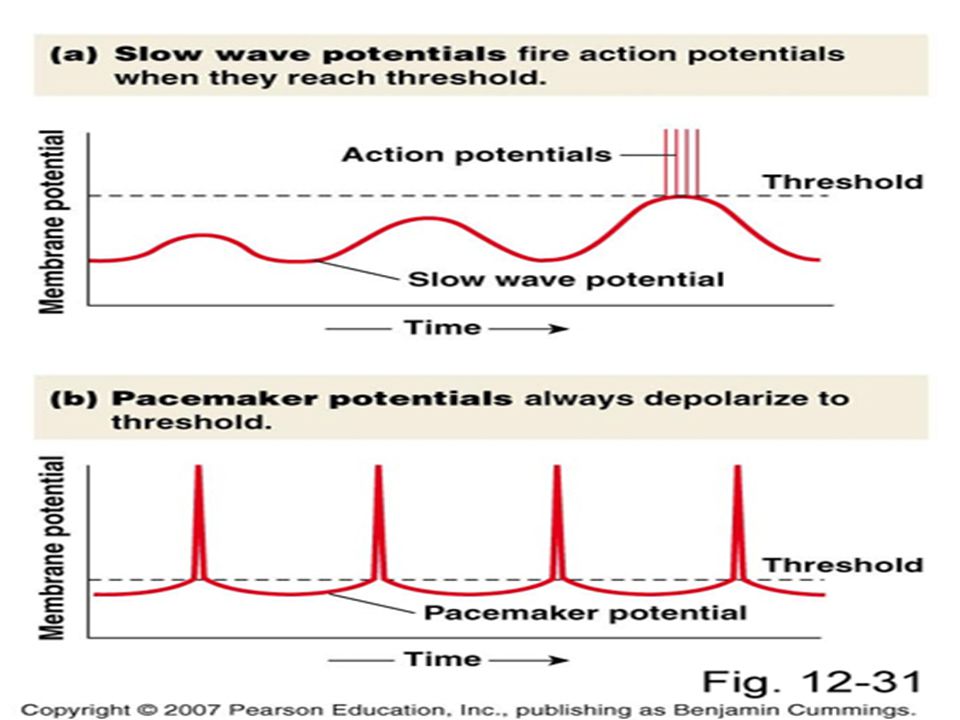
Üniter düz kasta yavaş dalga potansiyelleri ve spontan aksiyon potansiyeli oluşumu
Bazı düz kaslar kendi kendini uyarabilir (intestinal damar düz kası) Genelde membran potansiyelinde bazal bir yavaş dalga ritmi vardır Yavaş dalga bir AP değildir, lokal bir özelliktir Yavaş dalga ritmi, Na+ iyonlarının kas lifinden dışarı pompalanmasındaki artış ve azalmaya bağlı olarak ortaya çıkmaktadır
 Genelde membran potansiyelinde bazal bir yavaş dalga ritmi vardır. Yavaş dalga bir AP değildir, lokal bir özelliktir. Yavaş dalga ritmi, Na+ iyonlarının kas lifinden dışarı pompalanmasındaki artış ve azalmaya bağlı olarak ortaya çıkmaktadır.")
82
Yavaş dalga potansiyellerinin önemi
Yavaş dalga potansiyellerinin önemi, AP’ni başlatabilmeleridir. Yavaş dalga pot. -35 mv üzerine çıkarsa AP doğar. Her yavaş dalga pikinde 1 veya daha fazla AP oluşur, böylece düz kas kitlesinde bir seri kasılmalar oluşabilir, bu dalgalara PACEMAKER DALGALAR da denir

83
Tek Üniteli Düz Kas Elektriksel Özellikleri
Yavaş dalga Potansiyelleri (Slow Wave Potentials) Bazen kasılmaya neden olur bazen da olmaz. Bir aksiyon potansiyeli değildir. -35 mV ulaştığı zaman A.P oluşturur. Pacemaker dalga denir. Aksiyon Potansiyelleri (hep veya hiç) Spontan A.P oluşur. Yavaş dalga potansiyelleri etkileri Membran negatifliğinin azalması 21
 Bazen kasılmaya neden olur bazen da olmaz. Bir aksiyon potansiyeli değildir. -35 mV ulaştığı zaman A.P oluşturur. Pacemaker dalga denir. Aksiyon Potansiyelleri (hep veya hiç) Spontan A.P oluşur. Yavaş dalga potansiyelleri etkileri. Membran negatifliğinin azalması. 21.")
84
Multi Ünit Düz Kas Elektriksel Özellikleri
Multi ünit düz kasın membran potansiyeli stabildir Lifleri küçüktür. Kasılması için uyarıldığında aksiyon potansiyelleri oluşturmazlar. AP olmadan depolarizasyon gerçekleşir. Transmiter madde tarafından oluşturulan kavşak potansiyeller oluşur. Bu potansiyeller elektrotonik olarak tüm kasa yayılır 26

87
Teşekkür Ederim

Benzer bir sunumlar