Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Biyoreseptörler

2
GİRİŞ Doğadaki bütün canlıların, ister prokaryotik ister ökaryotik olsunlar, müşterek özellikleri canlı moleküller olan biyomolekülleri içermeleridir. Canlılar yaşamlarını bu biyomoleküllerin hücre organelleri, hücreler, dokular ve organlardaki anabolik ve katabolik reaksiyonları gerçekleştirmelenyle sürdürürler. Biyomoleküllerin, hücre organellerinin hücrelerin, dokuların ve organların birbirleriyle ve çevreleri ile etkileşimleri, birbirlerine karşı alfiniteleri analitik olarak incelenebilir. Bu da her birinin biyosensörler denilen analitik cihazların oluşumunda biyo- kısmında kullanılabilmelerini sağlar. Sensor kısım ise bu biyo kısımdaki metabolik değişiklerin optik, elektriksel veya kalorimelrik olarak algılanıp kaydedildiği kısımdır.

3
Bütün biyomoleküllerden, hücre organellerinden, hücrelerden, dokulardan veya organlardan biyosensörler yapılabilir ancak, ideal bir biyosensör doğada uygulanan bir sistem örnek alınarak modellenmelidir yani Iipidli hücre mernbranını kat eden özgün bir reseptör. Membran normal şartlarda bariyer olarak davranır ve reseptörler bu bariyer üzerinden sinyallerin transmisyonundan sorumludur. Genelde takip edilen yol şu şekilde özetlenebilir: Ligand yani reseptör için özgün olan molekül reseptöredifüze olur ve reseptörün hücre membranının dışındaki özgün bir yere bağlanır ya bağlanma reseptör yapısında konformasvonel bir değişikliğe yol açar ve bu değişiklik membranın iç yüzüne aktarılır ve sonuçla genellikle ya reseptörün ortasındaki kanal açılır ki bu kanaldan iyonlar geçer ya da "ikincil haberci" tabir edilen diğer moleküller aktive edilir ki bu da hücre içinde bir seri başka reaksiyonları başlatır.

4
Biyosensör cihazlarının oluşturulmasında ilk ve en önemli adım moleküler tanımadır. Bu adımdaki özgünlük ve afinite -hangi sinyal alıcı teknolojisi kullanılırsa kullanılsın- analitik sistemin performansini tayin edecektir. Biyoafiniteye dayanan biyoreseptör Enzimler ile oluşturulan biyosensörler genelde tek bir bileşeni tanımlarken diğer biyomoleküller daha geniş bir kimyasal sınıfın herhangi bir üyesinitanınılayabilir. Biyoreseptörlerle etkin herhangi bir biyosensör tasarımı için öncelikle biyoreseptörlerin ve agonistlerinin yapıları, bağlandıklarında oluşan konformasyonel değişiklikler ve bunun sonunda verilen ikincil sinyalleri sağlayan kimyasal molekül veya moleküllerin çok iyi bilinmesi ve anlaşılması gerekir.

5
2. BİYORESEPTÖRLERİN TANIMLANMASI
Hücreler arası iletiş bir hücresin yanındaki hücreye değmesiyle veya hormonlar, otokoidler veya nörotransmitörler tarafından gerçekleştirilir. Bu kimyasal maddeler hücre tarafından salgılanır ve hedef hücrelerdeki reseptörler tarafından tanınırlar. 3. BİYORESEPTÖRLERİN SINIFLANDIRILMASI Reseptörler bulunduklari yere göre iki gruba ayrılabilir: membran reseptörleri ve nükleer reseptörler. Reseptör kavramı 1900 yılları başlarında Langley tarafından ileri atıldığından beri endojenik ligandlarina göre isimlendirilmiştir.

6
Ancak reseptör moleküler birimlerinin son on onbeş yıldır tanımlanmalarıyla reseptörlerin fonksiyon ve temel yapılarına göre sınıflandırılmalarına başlanılmıştır. Reseptör proteinlerini kodlayan komplementer DNA'lann klonlanması ve amino asid dizilimlerinin tanımlanması birçok reseptörün nikotinik veya muskarinik reseptörlerine benzer yapısal karakteristiklere sahip olduklarini ve iyon kanalı reseptörleri ve G-proteini-bağımlı reseptörler süper ailelerini oluşturduklarını ortaya koymuştur. Membran Proteinleri İyon kanalı reseptörleri ve G-proteini-bağımlı reseptörler süper aileleri üyelerinin müşterek yapısal özellikleri transmembran bölümlerini oluşturan birkaç hidrofobik kısımdır. İyon kanalı reseptörlerinde dört, G-proteini-bağımlı reseptörlerde ise yedi tane hidrofobik kısım varken büyüme faktörü reseptörlerinin ve diğer protein reseptörlerinin tek bir transmembran bölgesi vardır. Her hangi bir reseptör için özgün ligandlarca tanımlanan bir çok ait tipler bulunmuştur

7
3.1.1. İyon Kanalı Reseptörleri
İyon kanalı reseptörlerinin endojenik ligandları nörotransmitörler olan asetilkolin, glutamat, ɤ -aminobutrik asid (GABA), glisin, serotonin ve muhtelemen ATP dir. İyon kanalı reseptörleri oligomerlerden oluşmuştur. Bazıları homojen alt birimler içerse de çoğu heterojen alt birimlerden oluşmuştur. Asetilkolin, glutamat ve serotonin katyon kanalları olup hem Na+ hem de K+ iyonları için geçirgendir. GABA ve glisin ise Cl" iyonları kanallarıdır. Nörotransmittörlerin postsinaptik membranlardaki bu reseptörlere bağlanmasıyla iyon kanalları açılır ve bu iyonlar içeri alınır.
, glisin, serotonin ve muhtelemen ATP dir. İyon kanalı reseptörleri oligomerlerden oluşmuştur. Bazıları homojen alt birimler içerse de çoğu heterojen alt birimlerden oluşmuştur. Asetilkolin, glutamat ve serotonin katyon kanalları olup hem Na+ hem de K+ iyonları için geçirgendir. GABA ve glisin ise Cl iyonları kanallarıdır. Nörotransmittörlerin postsinaptik membranlardaki bu reseptörlere bağlanmasıyla iyon kanalları açılır ve bu iyonlar içeri alınır.")
8
3.1.1.1. Nikotinik Reseptörler
İki asetilkolin molekülünün oligomere bağlanması konformasyon değişikliği indükler ki bu da kanalın açılmasını sağlar. Bunu takiben kanal desensitize olarak kapanır. Nikotinik reseptörlerin cAMP-bağımlı protein kinaz, protein kinaz C veya tirozin kinaz ile fosforilasyonunun bu desensitziasyonu kolaylaştırdığı bulunmuştur. Glutamat Reseptörleri İyonotropik (iyon kanalları) ve metabotropik (G-proteini-bağımlı) reseptörler olarak iki sınıfa ayrılırlar, lyonotropik reseptörler özgün agonistlerine göre alt sınıflara ayrılır. Moleküler özellikleri nikotinik reseptörlerine oranla daha az anlaşılmıştır ancak temel özelliklerinin benzer oldukları sanılmaktadır.
 ve metabotropik (G-proteini-bağımlı) reseptörler olarak iki sınıfa ayrılırlar, lyonotropik reseptörler özgün agonistlerine göre alt sınıflara ayrılır. Moleküler özellikleri nikotinik reseptörlerine oranla daha az anlaşılmıştır ancak temel özelliklerinin benzer oldukları sanılmaktadır.")
9
GABA Reseptörleri GABAA( iyon kanalı) ve GABAB (G-proteini-bağımlı) olmak üzere iki tur reseptör vardır, on dört çeşit GABAA reseptör alt birimi tanımlanmıştır. Bu alt birimlerin farklı kombinasyonları ile oluşturulan in situ bir çok oligomerin olduğu düşünülmektedir. GABAA anyon kanalı reseptörüdür ve temel yapısı nikotinik reseptörüne benzer sadece GABAA transmembran bölgesinin iki yanında kalan kısımlarda daha çok pozitif amino asidler bulunur ki bunların Cl* iyonlarının geçmesine katkısı olduğu sanılmaktadır
 ve GABAB (G-proteini-bağımlı) olmak üzere iki tur reseptör vardır, on dört çeşit GABAA reseptör alt birimi tanımlanmıştır. Bu alt birimlerin farklı kombinasyonları ile oluşturulan in situ bir çok oligomerin olduğu düşünülmektedir. GABAA anyon kanalı reseptörüdür ve temel yapısı nikotinik reseptörüne benzer sadece GABAA transmembran bölgesinin iki yanında kalan kısımlarda daha çok pozitif amino asidler bulunur ki bunların Cl* iyonlarının geçmesine katkısı olduğu sanılmaktadır.")
10
3.1.2. G-Proteini-Bağımlı Reseptörler
G-proteini-bağımlı reseptörlerin endojenik ligantlan glisin dışındaki bütün nörotransmitörleri çoğu hormonları ve otokoidleri, birkaç kemotaktik faktörü ve koku gibi eksogenik stimülantları içerir. Bu ligantların kimyasal türleri çok farklı olup aminleri, amino asidleri. nükleotidleri, lipidieri, peptidleri ve proteinleri kapsar. Işık reseptörü olan rodopsin de G-proteini bağımlı bir reseptördür. Her G-proteini bağımlı reseptör homojen bir proteinden oluşmuş olup büyük bir ihtimalle monomer halinde bulunur. Bu reseptörlerin fonksiyonu G-proteinlerini aktive etmektir yani G-proteininden GDP nin ayrılmasını stimüle etmek böylece GTP nin G-proteinlerine bağlanmasını kolaylaştırmaktır.

11
3.1.2.2. G-Protein-Bağıınlı Reseptörlerin
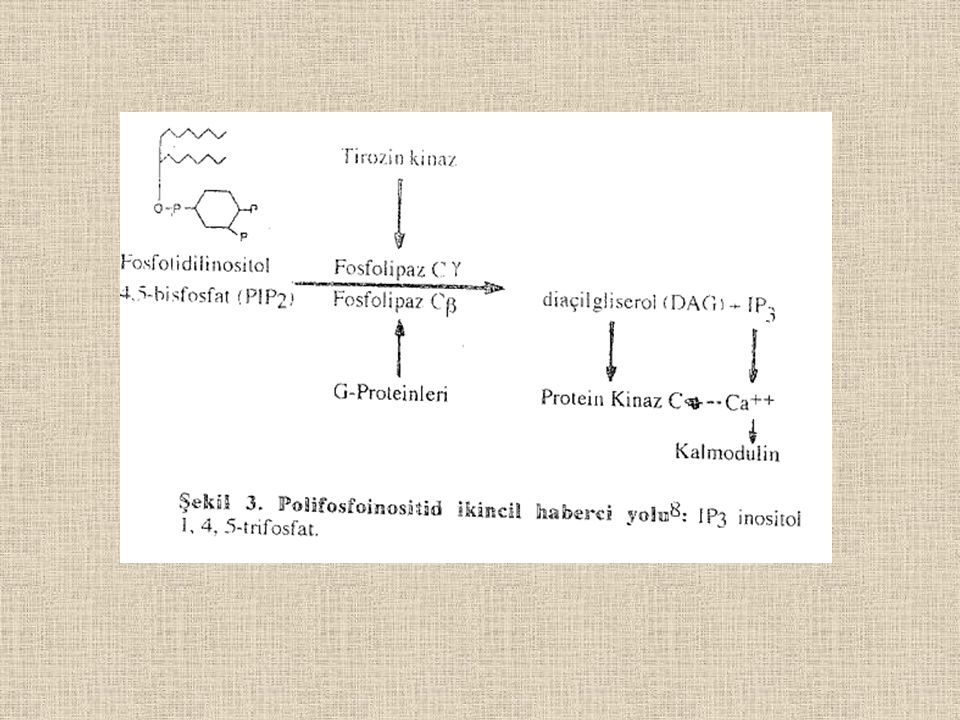
Sınıflandırılması ve Aksiyon Mekanizmaları Reseptörler aktive ettikleri G-proteinlerinin a- alt birim türlerine göre sınıflandırılabilir. ökarvotık hücrelerde metabolizmanın regülasyonu temel olarak önemli ve yaygın olan cAMP bileşiği ile bağlantılı protein kinazeler denilen enzimler sistemine gerçekleştirilir. Agonistin reseptörüne bağlanmasıyla aktifleşen G-proteini aracılığı ile aktifleşen adenilat siklaz, ATP den cAMP bileşiğini üretir ve adenilat siklaz ve G-proteini membrana bağlanırken ikincil haberci olarak harakel etmek üzere cAMP hücre içine bırakılır. Adenilat siklazı aktive eden ve adenilat siklazı inhibe eden G-proleini-bağımlı reseptörlerin mekanizması Şekil 1 ve 2 de verilmiştir.

13
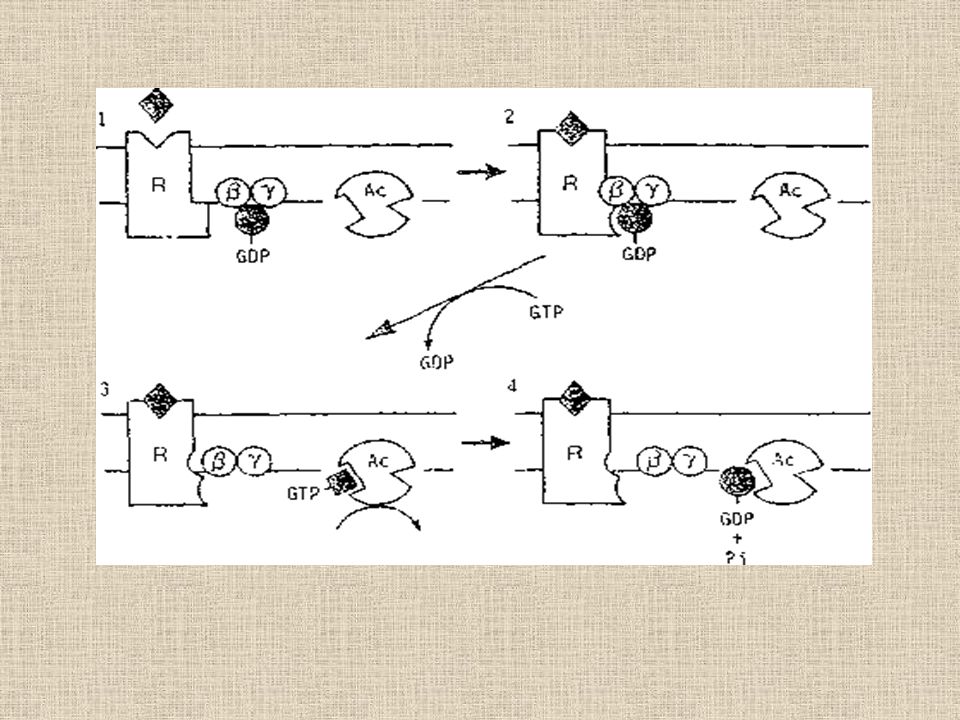
Şekil 1.adenilat siklaz enziminin G-Proteini aracılığı ,ile Aktivasyonu
(1) Reseptör- adenilat siklaz sistemi reseptör(R) Enzimi (AC adenilat siklazin katalitik birimi) ve modulatör protein (2) Hormon gibi bir agonistin reseptörüne bağlanması G-proteininin hormon-reseptör kompleksine bağlanmasına neden olur ki bu şeklinde GDPnin yerine GTP yi alır (3) GTP-a alt birimi ayrılır ve AC enzimine bağlanarak enzimi aktive eder ve ATP den siklik AMP (cAMP) oluşur (4) alfa-alt biriminde GTPaz aktivitesi vardır dolayısı ile GTP, GDP ve Pi hidrolizlenir. Hidrolizlenme AC yi inaktifleştirerek (2) deki kompleksin tekrar oluşmasını sağlar.
 Reseptör- adenilat siklaz sistemi reseptör(R) Enzimi (AC adenilat siklazin katalitik birimi) ve modulatör protein. (2) Hormon gibi bir agonistin reseptörüne bağlanması G-proteininin hormon-reseptör kompleksine bağlanmasına neden olur ki bu şeklinde GDPnin yerine GTP yi alır. (3) GTP-a alt birimi ayrılır ve AC enzimine bağlanarak enzimi aktive eder ve ATP den siklik AMP (cAMP) oluşur. (4) alfa-alt biriminde GTPaz aktivitesi vardır dolayısı ile GTP, GDP ve Pi hidrolizlenir. Hidrolizlenme AC yi inaktifleştirerek (2) deki kompleksin tekrar oluşmasını sağlar.")
14
Oluşan cAMPnin konsantrasyonu adenilat siklaz ve ribozun 3'-OH ve fosfat arasındaki bağı hidrolizliyen fosfodiesteraz ile birlikte regule edilir.

15
Şekiil 2. Adenilat siklaz enziminin G-proteini aracılığıyla inhibisyonu:
Bazı reseptörlerin aktivasyonu adenilat siklazın inhibisyonuna sebep olur ki bu belirgin bir G-proteini aracılığıyla olur. Sitimüle edici G proteini αs inhibe edici G proteini αi olarak gösterilmiştir. Aktive edici ve inhibe edici G-proteınlennin β ve γ alt birimleri aynıdır ancak α – altbirimleri belirgin olarak farklıdır.

16
Protein Kınazlar cAMP nin aksiyonu, iki regülatör ve iki katalitik alt birimi olan tetramer yapıdaki bir protein kinaz enzimini sitimüle etmektir. cAMP enzimin regülatör dimerine bağlandığında, iki aktif katalitik altbirimler serbest bırakılır. Aktif kinaz bir hedef enzimin fosforilasyomımı katalizler. Fosforilasyon yeri çoğunlukla serin ve tirozinin hidrodksil gruplarıdır. Enzimlere transfer edilen fosfat gruplarının kaynağı ATPdir. Bundan sonra hedef enzim hücresel cevabın verilmesini sağlar.

18
3.1.3. Tek Transmembran Bölümlü Reseptörler
Epidermal büyüme faktörleri, fibroblast büyüme faktörleri ve sinir büyüme faktörleri gibi bir grup büyüme faktörü reseptörleri ligand sitimülasyonlu tirozin kinazlardır. Bu reseptörler üç kısımdan oluşurlar: ligandın bağlanmasından sorumlu olan ekstrasellüler kısım, tek transmembran kısmı ve hem tirozin kinaz hem tirozin kinaz sübstratı aktivite sitoplazmik kısım. 3.2. Nükleer Reseptörler Steroid reseptörleri gen ifadesini özgün yapısal genlerle doğrudan doğruya etkileşime girerek regüle eden süper aile moleküllerinin bir bölümünü oluştururlar. Süper ailenin diğer üyeleri arasında tiroid hormon reseptörleri, retinol reseptörleri ve onkogen v-erb A da bulunmakladır. Bu reseptörler steroidlere yüksek bir affiniteyle bağlanırlar. "Boş" yani steroidlerle bağlanmamış reseptörler hedef hücrelerin nükleuslarında bulunurlar.

19
3.2.1. Nükleer Reseptörlerin Yapıları
Steroid hormon reseptörlerin hepsi homolog yapıdadır hepsinin N-terminal kısmı transkripsiyon aktivasyon bölgesi (TAD); orta kısmı DNA-bağlayıcı bölge, ile hormon ve ısı şok proteinleri (hps) bağlanma bölgelerini; C- terminal kısmı dimerizasyon bölgesi ve ikinci bir TAD bölgesi içerir. Nükleer reseptörler yapılarına ve bağlandıkları DNA dizilimlerine göre üç aileye bölünebilir. Glukokortikoid ailesi en son ortaya çıkmış olan ailedir ve kortisol, aldosteron, androjen ve progesteron reseptörlerini içerir. Bu ailenin üyeleri temelde homodimerlerlerdir ve ısı şok proteini 90 a ihtiyaç gösterirler ve hormon cevap elementleri TGTTCT ters tekrarlanın bağlarlar.
; orta kısmı DNA-bağlayıcı bölge, ile hormon ve ısı şok proteinleri (hps) bağlanma bölgelerini; C- terminal kısmı dimerizasyon bölgesi ve ikinci bir TAD bölgesi içerir. Nükleer reseptörler yapılarına ve bağlandıkları DNA dizilimlerine göre üç aileye bölünebilir. Glukokortikoid ailesi en son ortaya çıkmış olan ailedir ve kortisol, aldosteron, androjen ve progesteron reseptörlerini içerir. Bu ailenin üyeleri temelde homodimerlerlerdir ve ısı şok proteini 90 a ihtiyaç gösterirler ve hormon cevap elementleri TGTTCT ters tekrarlanın bağlarlar.")
20
Tiroid hormon ailesi en eski ve en farklı grupu oluşturur
Tiroid hormon ailesi en eski ve en farklı grupu oluşturur. Bu grupta tiroid hormonu, vitamin A ve D, ekdison ve araşidonik asid reseptörleri bulunur. Bu reseptörler heterodimerier olarak çok aktiftirler, ısı şok proteini 90 a ihtiyaç göstermezler ve hem doğru hem de ters hormon cevap elementleri TGACC yi bağlarlar. Östrojen ailesi sadece östrojen reseptörünü ve henüz ligandları tanımlanmamış birkaç benzer reseptörü içerir. Bu aile diğer iki aile özellikleri arasında özellik sergiler: tiroid hormon ailesi gibi hormon cevap elementleri TGACC yi sadece ters tekrarlar şeklinde bağlarlar, glukokortikoid ailesi gibi homodimerdirler ve ısı şok proteinlerine ihtiyaç gösterirler

21
4. PROTEİN OLMAYAN RESEPTÖR VE ANTİJENLER
Glikosifingolipidler Hücre membran molekülleri olan glikosifıngosinler bütün hayvan ve bazı bitki hücrelerinde dağılmış olarak bulunurlar. Doku-bağımlı bir dağılım sergileyen glikosifıngolipidlerin hücresel seviyede en önemli fonksiyonu türler, aynı türün bireyleri ve birey deki hücreler arasında oluşturdukları farklılıktır. Bunun en iyi örneği kan grubu antijenlerinde görülür. Glikosifingolipidler hücre yüzeylerinde bulunan reseptör proteinleriyle etkileşime girerek hücre dışında bulunan büyüme faktörlerine cevabı inhibe edebilirler.

22
4.2. Gükosifingosidlerin Yapısı
Bütün glikosifingolipidlerdekarbohidrat zinciri lipid molekülü seramide dik açı ile bağlanmış olarak bulunur. Glikosifingolipidleriu hepsi olmamakla birlikte büyük bir çoğunluğu seramide en yakın şeker moleküllerinin cins ve kimyasal bağlanışlarına göre tanımlanmış dört temel çekirdek yapısından birini paylaşırlar. Dört çekirdek yapısında da seramid glukoza, glıukoz ise galaktoza bağlanmıştır: (1) Ganglio-serisi glikosifingolipidlerde şeker zinciri başka bir galaktozun bağlı olduğu N-asetilgalaktozamin ile devanı eder. Hem iç hem de terminal galaktoz molekülü asidik şeker sialik asitle bağlıdır. (2) Globo-serisi glikosifingolipidlerde şeker zinciri başka bir gataktoz ve N-asetil-galaktozamin ile devam eder. (3) Lakto Tip I-serisi ve (4) Lakto Tip H-sehsi glikosifingolipidlerde şeker zinciri N-asetilglukozamin ve galaktoz ile devam eder ve son bağın pozisyonuna göre ayrılırlar. Lakto-serisi glikosifingolipidler uzayıp dallanarak aralarında klinik olarak önemli ABH kan-grubu antijenlerinin de bulunduğu çeşitli moleküller oluşturabilirler.
 Ganglio-serisi glikosifingolipidlerde şeker zinciri başka bir galaktozun bağlı olduğu N-asetilgalaktozamin ile devanı eder. Hem iç hem de terminal galaktoz molekülü asidik şeker sialik asitle bağlıdır. (2) Globo-serisi glikosifingolipidlerde şeker zinciri başka bir gataktoz ve N-asetil-galaktozamin ile devam eder. (3) Lakto Tip I-serisi ve (4) Lakto Tip H-sehsi glikosifingolipidlerde şeker zinciri N-asetilglukozamin ve galaktoz ile devam eder ve son bağın pozisyonuna göre ayrılırlar. Lakto-serisi glikosifingolipidler uzayıp dallanarak aralarında klinik olarak önemli ABH kan-grubu antijenlerinin de bulunduğu çeşitli moleküller oluşturabilirler.")
23
5.1. Immünolojik Biyoreseptörler 5.1.1. Antibodiler
5. BİYOSENSÖRLERDE KULLANILAN BİYORESEPTÖR VE BİYORESEPTÖR KISIMLARI Biyosensörlerde kullanılan biyoreseptörler ve biyoreseptorlerin aktif kısımları genelde ait oldukları biyolojik sisteme, kullanım amacına ve alanına göre sınıflandırılabilir: sağlık alanı (immünolojik reseptörler) ve endüstride ilaç sanayisinde(Farmakolojik reseptörler) ve gıda sanayisinde (Nükleik asidler). 5.1. Immünolojik Biyoreseptörler Antibodiler İmmün cevabını başlatan ana proteinler antibodi denilen immünoglobinlerdir. Tek membran bölgeli B-hücre yüzeyinde bulunan bu antijen reseptörlerinden en iyi anlaşılmış ve temini en kolay olanı lgG sınıfı antibodilerdir. Bu antibodiler in vitro kullanım için tavşan, koyun, keçi, fare sıçan gibi Hayvanların özgün antijenle immunize edilmeleriyle elde edilir.
 ve endüstride ilaç sanayisinde(Farmakolojik reseptörler) ve gıda sanayisinde (Nükleik asidler) Immünolojik Biyoreseptörler Antibodiler. İmmün cevabını başlatan ana proteinler antibodi denilen immünoglobinlerdir. Tek membran bölgeli B-hücre yüzeyinde bulunan bu antijen reseptörlerinden en iyi anlaşılmış ve temini en kolay olanı lgG sınıfı antibodilerdir. Bu antibodiler in vitro kullanım için tavşan, koyun, keçi, fare sıçan gibi Hayvanların özgün antijenle immunize edilmeleriyle elde edilir.")
24
Aynı antijene farklı affinitede birçok antibodi kolonisi oluştuğundan elde edilen bu antibodilere poliklonal antibodiler denilir. Daha özgün ve homolog antibodi üretimi için hibridoma teknolojisi kullanılır ve bu şekilde tek bir antibodi üreten B-limfositleri, B-hücre tümör çizgisi ile füzyona uğratılarak tek kolonili hibridoma çizgisi elde-edilir-ve bunlara monoklonal antibodiler denilir. Hibrid hücre çizgisinin avantajı homojen özgünlükte, afinitede ve izotipte antibodi üretmesidir ki bu antibodilerin in vivo ve in vitro olarak tanı amaçlı kullanımı çok yaygındır

25
Antibodilerin dikkate değer karakteristiklerinden birisi bifonksiyonel reseptörler olmaları ve molekülün hiper değişken N-terminal kısımlarında ikiz tanımlama yeri olmasıdır. Antijen bağlama kısımlarının bulunduğu iki F(ab)2 bölgesi ve kristalize edilebilen ağır zincirlerin C terminal bölgesi Fc -ki komplementer bağlanma ve-makrofej bağlanma bölgelerini içerir-birbirinden biyoafinitelerini kaybetmeden ayrılabilir ve biyosensörlerde tanımlama komponenfi olarak kullanılabilir. Sadece tek bir Fab bölgesi de aynı amaçla kullanılabilir. Hatta biyoafiniteyi arttırmak ve geniş özgünlük aralığı temini amacıyla genetik mühendisliği ile rekombinant DNA teknolojisi kullanılarak hafif ve ağır zincirlerin süper değişken bölgeleri tek bir zincir üzerinde peş peşe konularak sFv geni inşa edilmek suretiyle (ve bu genin PCR ile çoğaltılarak plazmid içine klonlanmasıyla ve uygun bir vektör ile uygun bir E. Coli susu içinde ifade ettirilerek bol miktarda ) yeni bir sistem ve yapıda antibodiler oluşturulmuştur.
 yeni bir sistem ve yapıda antibodiler oluşturulmuştur.")
26
5.2. Farmakolojik Reseptörler
Tarihsel olarak ilaç endüstrisinin araştırma çalışmaları vücuttaki farklı kontrol sistemlerini özgün bir şekilde etkiliyen ilaçları tasarlamaya yöneliktir. Son birkaç onyılda biriken bilgiler bu ilaçların etkilerinin genellikle membran üzerindeki protein yapıdaki reseptörler olduklarını ortaya koymuştur. Farmakolojik reseptörlerin özel tanımlama özelliği ilaçların, inhibitörierin ve loksik maddelerin miktarını ayarlama da bu reseptörleri biyosensörler için ideal kılmaktadır. Antibodilerde olduğu gibi ilaçların membran biyoreseptörlerine bağlanma kinetiği basit bir biyokimyasal olay olmayabilir zira çoğunlukla bu reseptörlerin birden fazla bağlanma yeri vardır ve bağlanmayla konformasyon değişikliğinden geçerler ki bu da fonksiyonel kinetiklerini çok kompleks bir hale getirebilir.

27
5.2.1. Biyomimetik Yaklaşımlar
Biyoreseptörterdeki moleküler tanımlamanın anlaşılması, istenen bağlanma karakteristiği içeren, fonksiyonel kinetiği basitleştirilmiş ve biyosensör imaline uygun reseptörlerin sentezlenmesini şimdilerde daha avantajlı hale getirmiştir. Optimal moleküler konfigürasyonlar artık bilgisayar programlarıyla modellenip simüle edilebilmektedir. 5.3.Nükleik Asidler RNAların da diğer RNA ları ve özellikle mRNA ların amino asidleri yüksek tanıma potansiyeli gelişigüzel polinükleotid kütüphaneleri sentezleyerek bu kütüphanelerin belirli ligandiarı bağlanma kapasitelerinin taraması yeni reseptör moneküllerinin tanımlanması kadar mevcut reseptörler arasından biyosensör tasarımı için aday reseptörlerin de seçimine imkan verebilecekgörülmektedir. , Bu teknik yeni ilaçlann tasarımında kullanılmaya başlanmıştır.

Benzer bir sunumlar








>")



>")
 Nedir? Lipitlerdir.>")

.>")



 PC KOPAT>")