Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
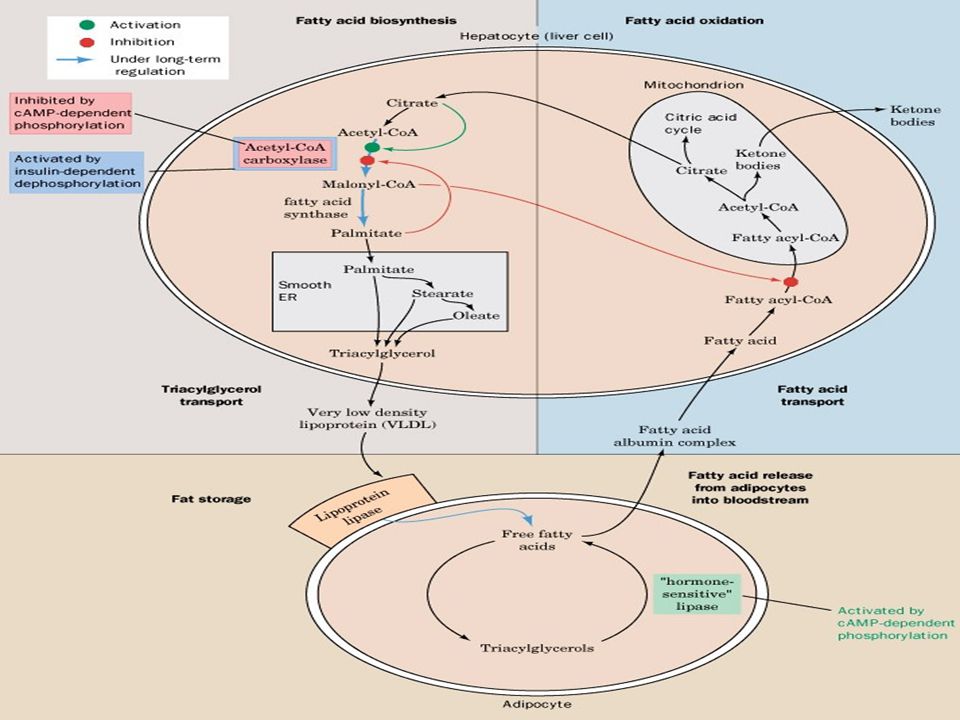
Yağ asitlerinin oksidasyonu
Yağ asitleri TG halinde depo edildiklerinden lipaz enzimleri ile hidrolize edilirler. 1904 yılında Franz Knoop oksidasyon mekanizması hk ilk bilgileri ortaya koydu. 1949 da Kennedy ve Lehninger oks. Mitekondria 1951 de ise aktifleşme basamağı yağlar kh verdiği enerji ile yanar Kc ne zamanki glukozun fazlasından TG yapacak olursa yağların enerjisi azalır

3
Yağ asitlerinin oksidasyonu
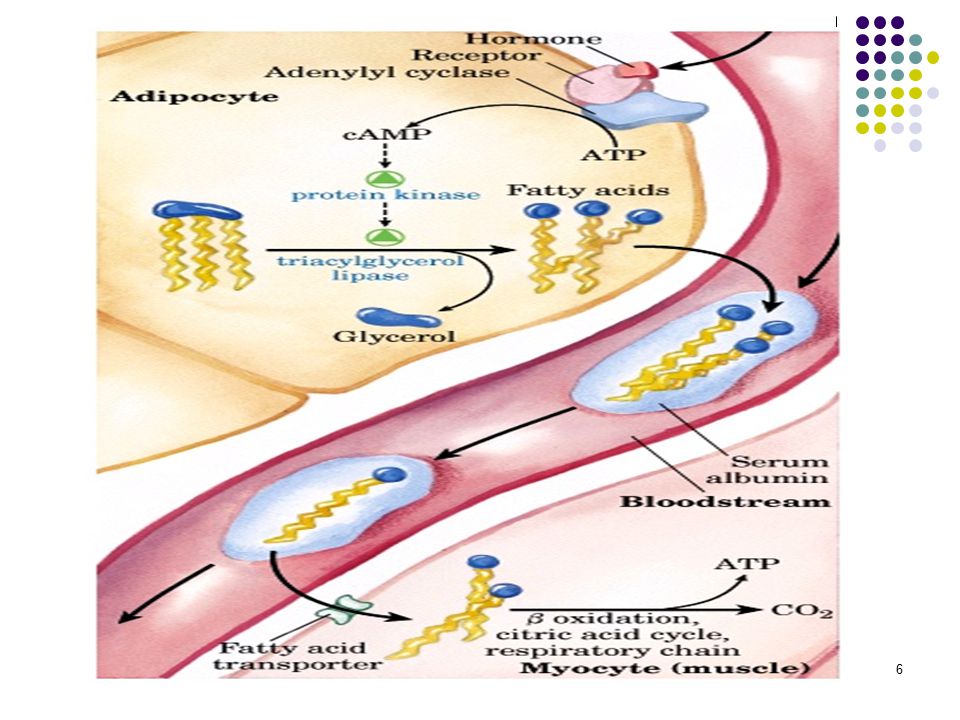
Depo yağları devamlı olarak mobilize olurlar ve yerlerine yenileri depo edilir. Lipidlerin adipoz doku depolarından oksidasyon için karaciğere ve diğer dokulara yağ asitleri şeklinde mobilize edildiklerine inanılır. Yağ dokuda trigliseridlerin hidrolizi lipoliz olarak tanımlanır.

4
Lipoliz Lipoliz, hormona duyarlı triaçilgliserol lipaz tarafından katalizlenir ve düzenlenir. 4

5
Depolanmış triaçilgliserollerin mobilizasyonu:Azalan kan glukoz düzeyi glukagonun salınmasınını tetikler. Bu durumda (1-8) numaralı reaksiyonlar aktif hale gelir ve triasilgliserollerden elde edilen yağ asitleri dolaşıma verilir. Albumine bağlanarak taşınan yağ asitleri kas dokuda β-oksidasyona uğrar. Perilipin: Nötral lipidler yağ dokuda perilipin adı verilen bir protein tarafından sarılarak kaplanırlar. PERİLİPİN yağ damlacıklarının yüzeyini kaplayarak onları hormona duyarlı lipazdan korur. Eğer Protein Kinaz A tarafından fosforlanırsa, Perilipin A'nın yapısı değişir ve artık yağ damlasını örtmez. Bunun üzerine lipaz, yağ damlacığını yüzeyine bağlanıp trigliseritleri hidrolizlemeye başlar.

7
Stage 3

8
Lipoliz, hormona duyarlı triaçilgliserol lipaz tarafından katalizlenir ve düzenlenir.

9
Yağ asitlerinin oksidasyonu
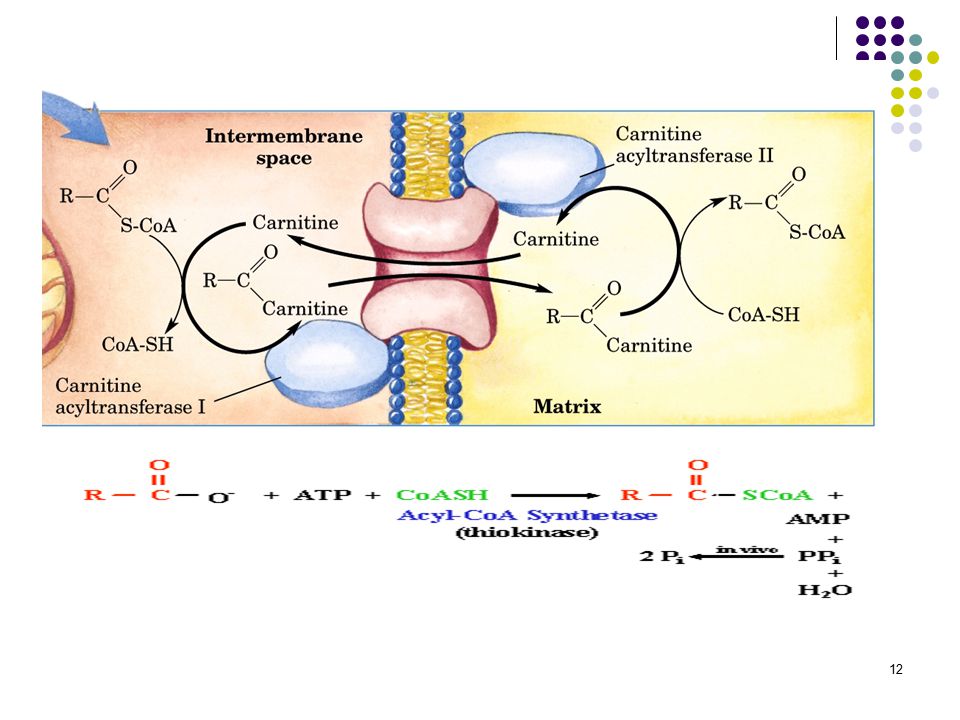
Kandan sitozole giren yağ asitleri, mitokondriyal membranları doğrudan geçemezler; ancak bir seri enzimatik reaksiyona uğradıktan sonra oksidasyon için mitokondri matriksine alınırlar. Bunun için önce yağ asidi, dış mitokondriyal membranda bulunan yağ açil-CoA sentetaz tarafından katalizlenen bir reaksiyonda aktiflenir; yağ açil-CoA oluşur.

10
LPL = lipoprotein lipase FABP = fatty acid binding protein
CAPILLARY L P Lipoproteins (Chylomicrons or VLDL) [2] FABP FA [3] MITOCHONDRION FABP acyl-CoA [4] acetyl-CoA TCA cycle -oxidation [6] [7] A C S albumin FA from fat cell [1] FA carnitine transporter acyl-CoA [5] CYTOPLASM Plazma serbest (esterleşmemiş) yağ asitleri serum albumini tarafından taşınırlar. Serbest yağ asitleri (FFA) kaynak noktası olan yağ dokusunun triaçilgliserolü veya dolaşımdaki lipoproteinlerden enerji üretim yeri olan karaciğer ve kas gibi birçok dokuya kadar bu şekilde taşınırlar. Dokuya geldiklerinde hücre içine geçen serbest yağ asitleri, yağ asidi bağlayıcı proteine (FABP) bağlanır. cell membrane FA = fatty acid LPL = lipoprotein lipase FABP = fatty acid binding protein ACS = acyl CoA synthetase
 [2] FABP. FA. [3] MITOCHONDRION. FABP. acyl-CoA. [4] acetyl-CoA. TCA. cycle. -oxidation. [6] [7] A. C. S. albumin. FA. from. fat. cell. [1] FA. carnitine. transporter. acyl-CoA. [5] CYTOPLASM. Plazma serbest (esterleşmemiş) yağ asitleri serum albumini tarafından taşınırlar. Serbest yağ asitleri (FFA) kaynak noktası olan yağ dokusunun triaçilgliserolü veya dolaşımdaki lipoproteinlerden enerji üretim yeri olan karaciğer ve kas gibi birçok dokuya kadar bu şekilde taşınırlar. Dokuya geldiklerinde hücre içine geçen serbest yağ asitleri, yağ asidi bağlayıcı proteine (FABP) bağlanır. cell membrane. FA = fatty acid. LPL = lipoprotein lipase. FABP = fatty acid binding protein. ACS = acyl CoA synthetase.")
11
Yağ asitlerinin oksidasyonu
Dış mitokondriyal membranda oluşan uzun zincirli yağ açil-CoA bileşikleri, iç mitokondriyal membrandan geçemezler; bunların mitokondriyal matrikse alınmaları karnitin gerekir.4-12 Karbonlu yağ asitlerinin mitokondri matriksine taşınımları için karnitine ihtiyaç yoktur.

13
Yağ asitlerinin oksidasyonu
13

14
Yağ asitlerinin oksidasyonu
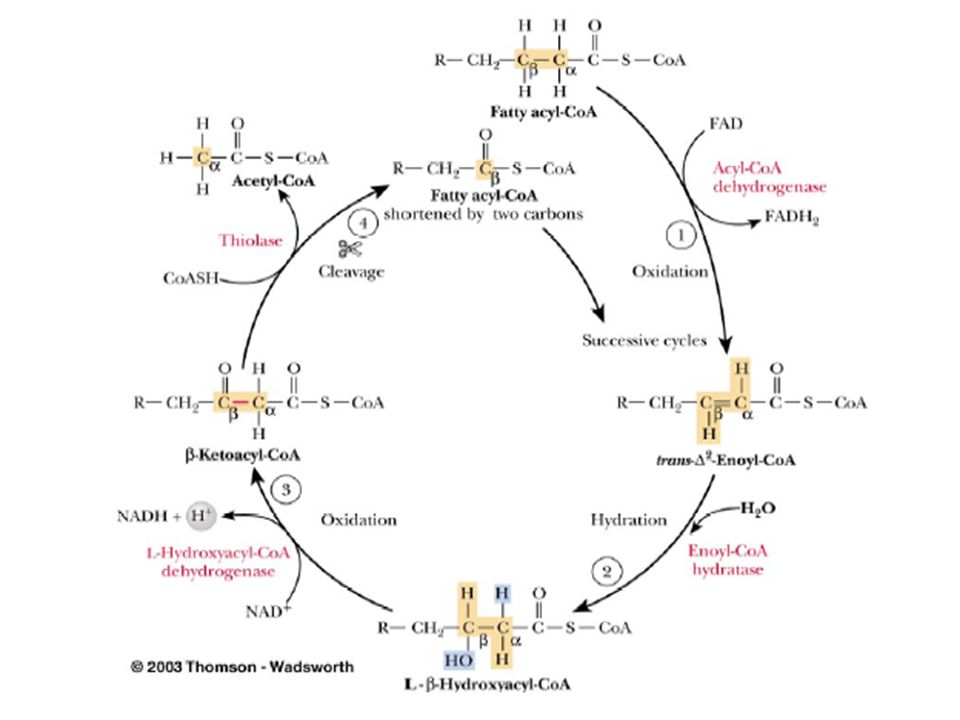
Yağ açil-CoA mitokondriyal matrikse alındıktan sonra oksidasyon denen yolda yağ asitlerinden, karboksilli uçtan başlayarak asetil-CoA şeklinde iki karbonlu üniteler art arda çıkarılır. Yağ asitlerinin oksidasyonundan başka ve oksidasyon denen oksidasyon yolları da vardır. Karnitin biyosentezinden veya böbreklerden geri emiliminden kaynaklanan karnitin yetersizliği, kas krampları, halsizlik ve ölüme kadar değişen klinik belirtiler ortaya çıkarır.Kaslarda patolojik miktarda TAG depolanır.Tedavi karnitin+ orta zincirli yağ asitleri diyeti uygulamaktır.

15
Yağ asitlerinin oksidasyonu
Doymuş yağ asitlerinin oksidasyonunda, dört temel basamak vardır: -İlk oksidasyon ile trans-2-enoil-CoA oluşur -Hidrasyon ile L-3-hidroksiaçil-CoA oluşur -İkinci oksidasyon ile -ketoaçil-CoA oluşur -Tiyolaz ile orijinal yağ asidinin karboksil ucundaki iki karbon parçası, asetil-CoA olarak ayrılır ve geriye karbon sayısı orijinal yağ asidinden iki eksik yağ açil-CoA kalır.

16
Yağ asitlerinin oksidasyonu

17
Yağ asitlerinin oksidasyonu

18
Yağ asitlerinin oksidasyonu

19
Yağ asitlerinin oksidasyonu

20
Yağ asitlerinin oksidasyonunun açıklanan bu dört reaksiyonunun tekrarlanmasıyla yağ asidi tamamen asetil-CoA’lara yıkılmış olur. 20

22
ATP yield from the oxidation of palmitate: N.B. in the mitochondria:
Palmitoyl-CoA 7 x b-oxidation 7 NADH 7 FADH2 17.5 ATP 10.5 ATP 8 Acetyl-CoA N.B. in the mitochondria: NADH = 2.5 ATP FADH2 = 1.5 ATP TCA 24 NADH 60 ATP 8 FADH2 12 ATP Each turn of TCA = 10 ATP 8 GTP 8 ATP 108 ATP -2 ATP 106 ATP DGo’ = 9,800 kJ/mole

23
-Yağ asitlerinin oksidasyonu ile oluşan asetil-CoA’lar;
1) Başka yağ asitlerinin sentezinde kullanılır. 2) Keton cisimlerinin yapımında kullanılır. 3) Kolesterol sentezinde kullanılır. 4) Steroidlerin ön maddesi olarak kullanılır. 5) N-asetilglukozamin gibi maddelerin oluşumu için bazı maddelerin asetillendirilmesinde kullanılır. 6) Sitrik asit döngüsünde yıkılarak organizmaya gerekli olan enerjinin sağlanmasında kullanılır.
 Başka yağ asitlerinin sentezinde kullanılır. 2) Keton cisimlerinin yapımında kullanılır. 3) Kolesterol sentezinde kullanılır. 4) Steroidlerin ön maddesi olarak kullanılır. 5) N-asetilglukozamin gibi maddelerin oluşumu için bazı maddelerin asetillendirilmesinde kullanılır. 6) Sitrik asit döngüsünde yıkılarak organizmaya gerekli olan enerjinin sağlanmasında kullanılır.")
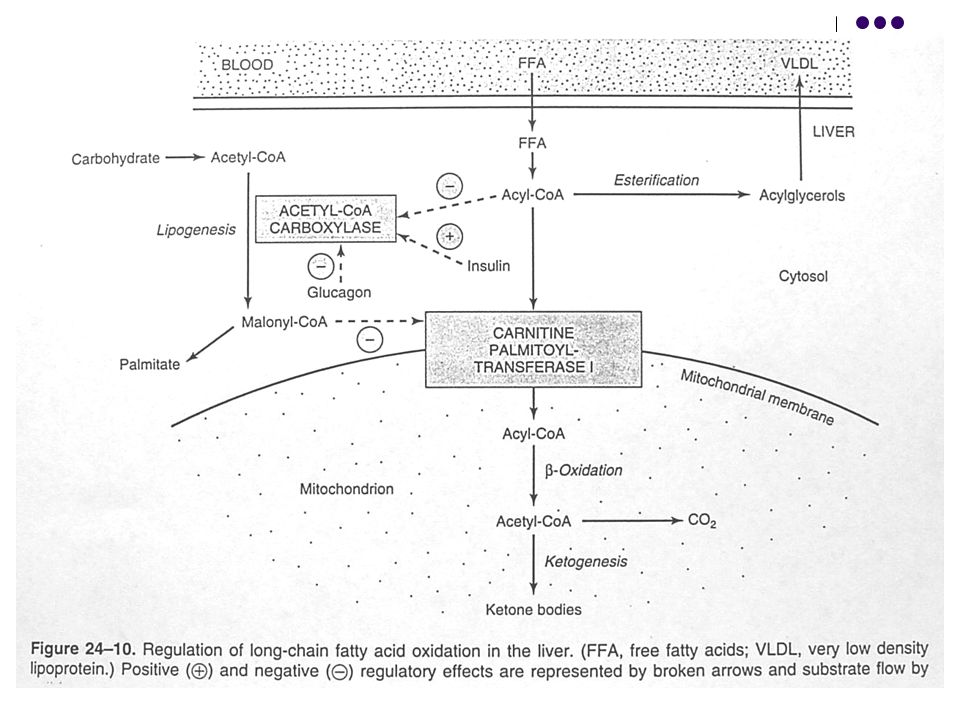
24
Aktiflenmiş yağ asidinin (yağ açil-CoA) mitokondriyal matrikse taşınması süreci, yağ asitlerinin oksidasyonunun düzenlenmesinde hız sınırlayıcıdır.
 mitokondriyal matrikse taşınması süreci, yağ asitlerinin oksidasyonunun düzenlenmesinde hız sınırlayıcıdır.")
26
Oleoil-CoA (9) gibi bir monoansatüre yağ açil-CoA’nın oksidasyonu, çift bağa kadar, doymuş yağ asitlerinin oksidasyonundaki gibi olur. Daha sonra enoil-CoA izomeraz enzimi çift bağı cis-3-izomer durumundan trans-2-izomer durumuna dönüştürür. Trans-2-izomer durumundan sonra oksidasyon devam eder.

27
Poliansatüre yağ asitlerinin oksidasyonu, enoil-CoA izomerazdan başka NADPH’ye bağımlı 2,4-dienoil-CoA redüktaz enzimini de gerektirir; bu iki enzimin etkisiyle trans-2, cis-4-dienoil-CoA ara ürünü, trans-2-enoil-CoA haline dönüştürülür. Bundan sonraki olaylar da oksidasyondaki gibi devam eder.

28
ATP yield from the oxidation of Oleate: N.B. in the mitochondria:
Oleil-CoA 8 x b-oxidation 8 NADH 7 FADH2 20 ATP 10.5 ATP 9 Acetyl-CoA N.B. in the mitochondria: NADH = 2.5 ATP FADH2 = 1.5 ATP TCA 27 NADH 67.5 ATP 9 FADH2 13.5 ATP Each turn of TCA = 10 ATP 9 GTP 9 ATP 120.5 ATP -2 ATP 118.5 ATP

29
Tek karbon sayılı yağ asitleri, oksidasyon ile propiyonil-CoA’ya kadar çift karbon sayılı yağ asitleri gibi yıkılırlar. Propiyonil-CoA da propiyonil-CoA karboksilaz, metilmalonil-CoA epimeraz ve metilmalonil-CoA mutaz enzimlerinin etkisiyle sitrat döngüsünün ara ürünü olan süksinil-CoA’ya çevrilir.

30
ATP yield from the oxidation of 17C Fatty acids:
C17-CoA 7 x b-oxidation 7 NADH 7 FADH2 17.5 ATP 10.5 ATP 7 Acetyl-CoA + 1 Propionil CoA SüksinilCoA +5ATP N.B. in the mitochondria: NADH = 2.5 ATP FADH2 = 1.5 ATP TCA 21 NADH 52.5 ATP 7 FADH2 10.5 ATP Each turn of TCA = 10 ATP 7 GTP 7 ATP 98 ATP -2 ATP 96 ATP +5ATP= 101 ATP

31
Yağ asitlerinin oksidasyonu mitokondrilerden başka peroksizomlarda da gerçekleşir:
-Peroksizomlarda karbonlu veya dallı zincirli veya hidroksillenmiş yapıya sahip yağ asitleri okside edilir. -Yağ açil-CoA’ların peroksizoma girmeleri için karnitine gerek yoktur. -Yağ asitleri tümüyle asetil-CoA’ya yıkılmaz; zincir kısalması olur. -İşlem sırasında yüksek enerjili bağlar elde edilmez. Isı oluşur. -Çoğu enzimler farklıdır.FAD bağımlı Açil CoA dehidrogenaz tarafından alınan elektronlar moleküler oksijene aktarılarak H2O2 oluşturulur.

32
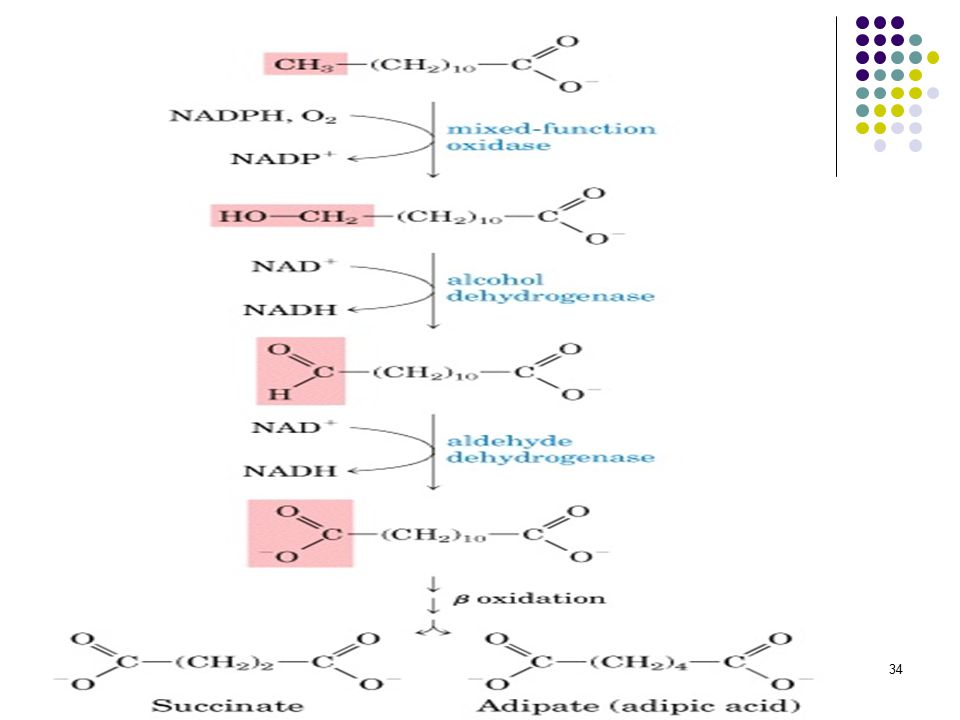
Yağ asitlerinin oksidasyonu
Yağ asitlerinin oksidasyonu, mikrozomlarda gerçekleşir. Molekülün karboksil ucundan her seferinde 1 karbon ayrılır. CoASH ve yüksek enerjili fosfatlar oluşmaz. Her seferinde 1 NADH elde edilir ve 1 karbon eksik yağ asidi oluşur.Özellikle dallanmış yapıdaki yağ asitleri için bu yol kullanılır. Moleküler oksijen, demir ve vitamin C gereklidir. Beyinde fosfolipidlerin yıkılışında izlenebilmektedir; sfingolipid sentezi için gereklidir. Refsum hastalığında fitanik asit birikmektedir. Az klorofilli diyet tedavi amaçlı kullanılır.

33
Yağ asitlerinin oksidasyonu
Yağ asitlerinin oksidasyonu, birçok dokuda endoplazmik retikulumda, karaciğerde mikrozomlarda gerçekleşir.Metil uçtaki karbon atomu oksitlenerek karboksil grubuna dönüşmektedir. Moleküler oksijen, NADPH ve sitokrom P-450 gereklidir. Önce karbonu oksitlenerek dikarboksilik asit oluşur; oluşan dikarboksilik asit de her iki uçtan oksidasyonla yıkılır.

35
Yağ asid oksidasyon bozuklukları
Karnitin eksikliği(Plazma serbest yağ asitlerinde artış) tedavide oral karnitin verilir. Karaciğer karnitin palmitoiltransferaz eksikliği Kas karnitin palmitoiltransferaz eksikliği(kas zayıflığı ve myoglobinüri) Jamaika kusma hastalığı (Akne ağacı meyvesinde bulunan hipoglisin açilCoA dehidrogenazı inaktive eder ve beta oksidasyon durur) Dikarboksilik asidüri (mitekondrial orta zincire spesifik açilCoA dehidrogenaz eksikliğinde görülür.) hekzonoilglisin birikimi Refsum hastalığı (fitanik asit katabolize edilemez dokularda birikir) Zellweger(serebrohepatorenal) sendronu(peroksizom eksikliği-uzun zincirli yağ asitlerinin oksidasyonu gerçekleşmez dokularda birikir)
 tedavide oral karnitin verilir. Karaciğer karnitin palmitoiltransferaz eksikliği. Kas karnitin palmitoiltransferaz eksikliği(kas zayıflığı ve myoglobinüri) Jamaika kusma hastalığı (Akne ağacı meyvesinde bulunan hipoglisin açilCoA dehidrogenazı inaktive eder ve beta oksidasyon durur) Dikarboksilik asidüri (mitekondrial orta zincire spesifik açilCoA dehidrogenaz eksikliğinde görülür.) hekzonoilglisin birikimi. Refsum hastalığı (fitanik asit katabolize edilemez dokularda birikir) Zellweger(serebrohepatorenal) sendronu(peroksizom eksikliği-uzun zincirli yağ asitlerinin oksidasyonu gerçekleşmez dokularda birikir)")
Benzer bir sunumlar