Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Statik manyetizmaya maruz bırakılan rat hipokampal nöron hücrelerinde N-Terminal Asparagine için amiodohidrolase vasıtasıyla ubikitin-proteosome yolunun stimulasyonu. Dr. Serdal ÖZDEMİR MALATYA

2
Takao Hirai,1,2 Hideo Taniura,2 Yasuaki Goto, Masato Ogura, Judy C. G. Sng and Yukio Yoneda Laboratory of Molecular Pharmacology, Division of Pharmaceutical Sciences, Kanazawa University Graduate School of Natural Science and Technology, Kanazawa, Ishikawa, Japan

3
ÖZET Beyin hücreleri arasındaki bütünleşme ve ayarlanımların statik manyetizma tarafından düzenlenmesinin altında yatan mekanizmaları aydınlatmak için, kültüre edilmiş rat hipokampüs nöronlarının zayıf manyetizmaya verdikleri gen ekspresyon yanıtlarını farklı analiz yöntemleriyle gözledik.

4
Rat beyninde manyetizma yanıtı sonucu olarak ilk tanımladığımız klon Ntan1(amidohydrolase for N-terminal asparagine) oldu.Ntan1 proteinlerin N- terminal kısımlarını bozan bir protein yıkım yolağının önemli bir bileşenidir, İn vivo olarak, in situ hibridizasyon yöntemi kullanılarak hipokampal nöronlarda bol miktarda Ntan1 mRNA sı eksprese edildiği görüldü.
 oldu.Ntan1 proteinlerin N- terminal kısımlarını bozan bir protein yıkım yolağının önemli bir bileşenidir, İn vivo olarak, in situ hibridizasyon yöntemi kullanılarak hipokampal nöronlarda bol miktarda Ntan1 mRNA sı eksprese edildiği görüldü.")
5
Nothern Blot analizleride zayıf manyetizmaya maruziyetten 3 saat sonra Ntan1 mRNA ekspresyonunun yaklaşık üç kat arttığını gösterdi. Zayıf manyetizma ayrıca Ntan1 promotor bölgesindeki transkripsiyonel aktiviteyide arttırdı.

6
Zayıf manyetizma, hipokampal nöronlarda 26S proteosomun selektif inhibitörü tarafından engellenen ve hücre morfolojisini ve yaşayabilirliğini etkilemeyen MAP2(mikrotübül association protein 2) yıkımını indükledi.
 yıkımını indükledi.")
7
Rekombinant Ntan1 adenovirüs vektörü kullanılarak aşırı miktarda Ntan1 ekspresyonu yapılan hücrelerde MAP2 ekspresyonunda belirgin azalma saptandı. Sonuçlarımız zayıf manyetizmanın rat hipokampüs hücrelerinde Ntan1 indüksiyonuna yol açarak ubikitin- proteosome yolu vasıtası ile MAP2 yıkımından sorumlu olduğunu gösterdi.

8
GİRİŞ Yaşayan organizmalar Yerkürenin yüzeyinde bulunan 50-70mT jeomanyetik alana sürekli olarak maruz kalmaktalar. Elektromanyetiğin ve süperiletkenlerin geliştirilmesi insanoğlunun güçlü manyetik alanlarla yüzyüze kalma ihtimalini doğurdu.

9
Manyetik rezonans görüntüleme yönteminin klinik kullanımda daha fazla geliştirilmesiyle jeomanyetik alanın onbinlerce kat daha büyükleriyle karşılaşmak bir rutin haline geldi.

10
Bununla birlikte manyetik alanların hayvan ve insan hücreleri üzerindeki etkilerine çalışılmış olan birkaç çalışma var. Merkezi sinir sisteminde, hem titreşimli hem de statik manyetik alanlar glial hücreler üzerinde ve elbetteki nöronlar arasında ki elektriksel ve mekanik mobiliteyi etkileyerek hücre fonksiyonlarında bir takım değişikliklere yol açabilir.

11
200 mT büyüklüğündeki statik manyetik alana maruz kalmak sadece morfolojik değişikliklere yol açmakla kalmayıp hem insan nöron hücre çizgisi FNC-B4 te inozitol lipit sinyallerinde hemde timidin birleşmelerinde anlamlı bir azalmaya yol açıyor. (Pacini et al. 1999).
..")
12
Sensorial nöron kültürlerindeki aksiyon potansiyelleri statik manyetik alanlar tarafından bloke ediliyor(Cavopol et all. 1995), değişik manyetik alanlar astrosit hücre kültürlerinde glial fibriler asidik protein ekspresyonunu arttırıyor.(Chan at all 1999).
, değişik manyetik alanlar astrosit hücre kültürlerinde glial fibriler asidik protein ekspresyonunu arttırıyor.(Chan at all 1999)..")
13
Bundan başka statik manyetik alana sürekli veya yineleyici olarak maruz kalmanın rat hipkampal nöronlarında nöron hücre markerı olan MAP2 protein ekspresyonununu hücre yaşamsallığını, nöron nükleusundaki diğer protein markırları ve büyüme markırı olan growth marker protein 43’ü belirgin derecede etkilemeden azalttığını kanıtladık.

14
Klinik çalışmalarda tekrarlayan transkraniel manyetik stimülasyonun(rTMS) BAB, şizofreni ve depresyon hastalarının tedavi ve terapilerinde, dirençli depresyon tedavisinde kullanılan EKT’ye bir alternatif olabileceği gösterilmiş. (George et al. 1995; Hasey 2001;McNamara et al. 2001; Post and Keck 2001)
.")
15
Bizde daha önce 100 mT statik manyetik alana maruz bırakılan rat hipokampal nöron kültürlerinde nükleer transkripsiyon faktör aktivatör protein-1 in DNA bağlanımında belirgin fakat geçici bir artış göstermiştik. (Hirai et al. 2002).
..")
16
Ap1, Jun ve Fos protein ailesi üyeleri arasında bir hetero ve homodimerdir ve çekirdekte TGACGTCA hedef dizelerini spesifik olarak tanıyarak hedef genlerin transkripsiyonunu düzenler.

17
Bir immünohistokimya çalışması, statik manyetik alana 30 dakika maruziyet sonucunda rat beyin hücrelerinin viseral ve vestibuler nukleuslarında c-Fos protein expresyonunun in vivo olarak indüklendiğini göstermiş.(Snyder et all 2000.)
")
18
Manyetik stimülasyon uygulanan rat paryetal korteks organotipik beyin kesitlerinde, 3-6 saat sonra tetradotoxin duyarlı sodyum kanalları aracılığıyla nöronlarda c-Fos protein ekspresyonunda geçici bir artışla sonuçlanırken astroglialarda artış görülmemiş.(Hausmann et all. 2001)
.")
19
Benzer şekilde parietal korteks ve hipokampus(Hausmann et all. 2000) haricinde frontal korteks, striatum, amigdala ve Ammon boynuzunda(Doi et all. 2001) tekrarlayan rTMS seansları sonrasında in situ hibridizasyon ve immmunohistokimya yöntemleriyle c-Fos ekspresyonunda belirgin bir indüksiyon gösterilmiş.
 haricinde frontal korteks, striatum, amigdala ve Ammon boynuzunda(Doi et all. 2001) tekrarlayan rTMS seansları sonrasında in situ hibridizasyon ve immmunohistokimya yöntemleriyle c-Fos ekspresyonunda belirgin bir indüksiyon gösterilmiş..")
20
Bu raporlar manyetizmanın nöronlarda gen transkripsiyonunu düzenleyerek uzun süreli etkilere yol açabileceğini önermektedir. Bu çalışmada kültüre edilmiş rat hipokampüs nöronlarının zayıf manyetizmaya verdikleri gen ekspresyon yanıtlarını farklı analiz yöntemleriyle gözledik ve Ntan1(N Terminal Asparagine for Amylhydroalse) genini manyetizmaya bir cevap olarak tanımladık.
 genini manyetizmaya bir cevap olarak tanımladık..")
21
Materyal ve Yöntem Nöron hücre kültürü:Primer hücre kültürleri Di Porzio’nun(1980) tanımladığı yöntemlerde küçük değişiklikler yapılarak taşındı.Özet olarak 18 günlük rat emriyolarının ayrıştırılan hipokampus hücreleri oda sıcaklığında,on iki dakikalığına Versen (Invitrogen, Grand Island, NY, USA) besiyerine ekildi.
 tanımladığı yöntemlerde küçük değişiklikler yapılarak taşındı.Özet olarak 18 günlük rat emriyolarının ayrıştırılan hipokampus hücreleri oda sıcaklığında,on iki dakikalığına Versen (Invitrogen, Grand Island, NY, USA) besiyerine ekildi.")
22
Daha sonra hücreler pasteur pipeti kullanılarak mekanik olarak ayrıştırıldı ve Dulmecco’nun modifiye Eagle besiyerinde[Besleyici karışım F-12 (1:1), invitrogen, %10 luk fetal sığır serumu] poly-L lizin (7.5 lg/mL, Sigma, St Louis, MO, USA) ile kaplanmış plastik tabak üzerine 2X105 hücre/cm2 olacak şekilde yayıldı.
![Daha sonra hücreler pasteur pipeti kullanılarak mekanik olarak ayrıştırıldı ve Dulmecco’nun modifiye Eagle besiyerinde[Besleyici karışım F-12 (1:1), invitrogen, %10 luk fetal sığır serumu] poly-L lizin (7.5 lg/mL, Sigma, St Louis, MO, USA) ile kaplanmış plastik tabak üzerine 2X105 hücre/cm2 olacak şekilde yayıldı.](http://images.slideplayer.biz.tr/33/10376914/slides/slide_22.jpg "Daha sonra hücreler pasteur pipeti kullanılarak mekanik olarak ayrıştırıldı ve Dulmecco’nun modifiye Eagle besiyerinde[Besleyici karışım F-12 (1:1), invitrogen, %10 luk fetal sığır serumu] poly-L lizin (7.5 lg/mL, Sigma, St Louis, MO, USA) ile kaplanmış plastik tabak üzerine 2X105 hücre/cm2 olacak şekilde yayıldı.")
23
Laboratuardaki 3 günden sonra besiyeri ;33 mM glikoz, 2 mM glutamin, 100 U/mL penisilin,100 lg/mL streptomisin, 5 mM HEPES, 13 mM sodyum bikarbonate,50 lg/ mL apo-transferrin, 500 ng/mL insulin, 1 pM b-estradiol,3 nM triiyodotironin, 20 nM progesteron, 8 ng/mL sodium seleniate, ve 100 lM putressine ile desteklendi.

24
Kültürlere 2-3. günler arası,çoğalan hücreleri azaltmak için 24 saat boyunca, sitosina arabinosid uygulandı. Kültürler %5 karbondioksit/%95 nemlendirilmiş hava ortamında 37 derecede 11-13 gün bekletildi.

25
Manyetizma maruziyeti:in vitro 11. günde, primer hipokampal nöronlar kültür plaklarının 10 cm. uzağına yerleştirilmiş demir oksit içeren mıknatıslarla 15 dakika süre boyunca 100mT statik manyetik alana maruz bırakıldı.Manyetik alanın şiddeti sürekli olarak ölçüldü[Tesla Meter TM-601 (Kanetec, Udea, Nagano,Japan)]
].")
26
Bir kontrol grubuna aynı çevresel koşullara sahip farklı bir ortamda mıknatıslar yerine tahta çubuklar kullanılarak benzer prosedürler uygulandı,Statik manyetik alana maruz bırakıldıktan sonra hücreler toplanana kadar 37 derecede CO2 etüvlerinde bekletildi.

27
Ayrımsa ekran analizleri:Statik manyetik alan uygulandıktan 11 gün sonra kullanılan ISOGEN kullanılarak hipokampal hücre kültürlerinin RNA’ları çıkarıldı.kontamine kromozamal DNA’dan ayırmak için DNAase I (Amersham Life Science, Little Chalfont, UK) uygulaması yapıldı.
 uygulaması yapıldı.")
28
Northern Blotting:cDNA fragmanları northern blot analizleri için REPIDRIME II primer etiketleme sistemi kullanılarak [a- 32P]dCTP ile işaretlendi.
![Northern Blotting:cDNA fragmanları northern blot analizleri için REPIDRIME II primer etiketleme sistemi kullanılarak [a- 32P]dCTP ile işaretlendi.](http://images.slideplayer.biz.tr/33/10376914/slides/slide_28.jpg "Northern Blotting:cDNA fragmanları northern blot analizleri için REPIDRIME II primer etiketleme sistemi kullanılarak [a- 32P]dCTP ile işaretlendi.")
29
Digoksijenin işaretli cRNA problarının hazırlanması: İn Situ Hibridizasyon: Revers trasnkriptaz polimeraz zincir reaksiyon analizleri: Western blott: Rekombinant Adenovirus: Protein yıkımının ve Ubikitin düzeyinin ölçümü:

30
SONUÇLAR: Farklı olarak regüle edilen bir gen olarak Ntan1 in differential display(DD) kullanılarak tanımlanması: Rat hipokampal nöron kültürleri 15 dakika boyunca 100 mT statik manyetik alana bırakıldıktan sonra AP-1 in DNA ya bağlanmasında geçici fakat belirgin bir artış olduğunu daha önce söylemiştik.
 kullanılarak tanımlanması: Rat hipokampal nöron kültürleri 15 dakika boyunca 100 mT statik manyetik alana bırakıldıktan sonra AP-1 in DNA ya bağlanmasında geçici fakat belirgin bir artış olduğunu daha önce söylemiştik.")
31
Manyetik alanda bırakılan hipokampal nöronlarda AP-1 aracılığıyla veya başka bir şekilde gen regülasyonu olup olmadığını tanımlamak için DD uygulandı. Manyetik maruziyetten 3 saat sonra total RNA hazırlandı ve daha sonra,dokuz aşağı inen yirmi dörtte yukarı çıkan primer içeren DD-PCR uygulanarak RNA lardan cDNA lar elde edildi.

32
.Biz çıkan sekizinci ve inen onbirinci primerlerin birleşiminden oluşan,manyetik alan amaruz bırakılan nöronların RNA havuzlarında daha fazla amplifikasyon veren bir cDNA fragmanına odaklandık. (Fig. 1a). Daha sonra farklı görünen bu ekspresyonu PCR reamplifikasyıon ile kontrol ettik. (Fig. 1b).
. Daha sonra farklı görünen bu ekspresyonu PCR reamplifikasyıon ile kontrol ettik. (Fig. 1b)..")
34
Fragman jelden çıkarılarak temizlendi, kopyalandı ve otomatize sekanslara sunuldu.Çoğaltılmış fragman,farenin N- terminal asparagine amidas’ının C- terminali ile yüksek düzeyde homolog olan ( %93) aynı dizileri içeriyordu
 aynı dizileri içeriyordu")
35
Rat Ntan1 DNA sı rat hücrelerinden elde edilen RNA lar kullanılarak reverse transkripsiyon ile sentezlendi.Bizim kullandığımız rat hücrelerinde Ntan1 dizileri farelerinkinden farklı olarak Ekson 9 delesyonunua uğramıştı ve 271 amino asitten oluşan bir proteini şifreliyordu.

36
Rat Ntan1 aminoasit dizileri fareninkisiyle %95 oranında homoloji gösterdi..Rat beyninde farklı Ntan1 izoformlarının ekspresyonunu gözlemlemek için,ekson spesifik primerler kullanarak RT-PCR analizleri yaptık.

38
Laboratuar ortamında rat beyninde Ntan1 mRNA larının ekspresyonu: Ntan1 aslında domuz karacieğrinin n- terminal asparaginini deaminasyona uğratan N-terminal amidase olarak tanımlanır.Ntan1’in bir proteinin N terminal rezidülerini destabilize eden ve N-end Rule oalrak isimlendirilen bir protein yıkım sinyalinin önemli bir komponenti olduğu düşünülüyor.

39
N-end Rule yolağına öbakterilerden E.coli, mayalardan Saccharomyces cerevisiae ve memeli hücrelerinden olmak üzere araştırıldığı her hücrede rastlanılmış. Farelerdeki Ntan1 geninin azalmış spontan aktivite göstermesi sebebiyle uzaysal hafızaları daha sınıldı oluyor, konjenik* fareler arasında sosyallikte daha resesif kalıyorlar (Kwon et al. 2000).
..")
40
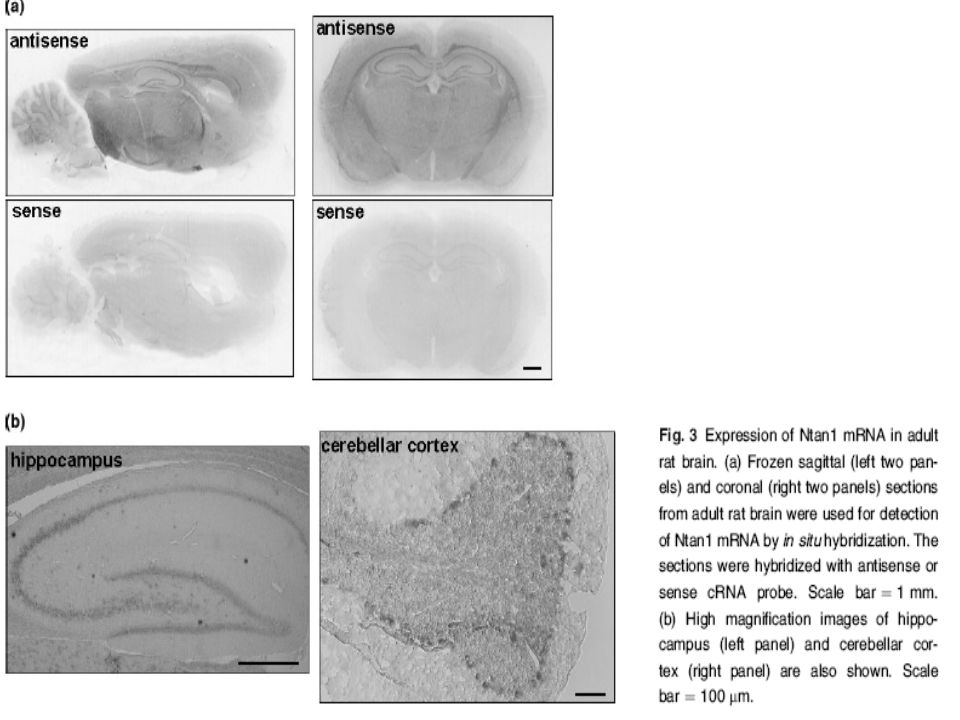
Bizde Ntan1 cDNA probları kullanarak yaptığımız in situ hibridizasyon yöntemleriyle Ntan1 mRNA’sı eksprese eden hücrelerin beyindeki dağılımını araştırdık.ntan1 mRNA pozitifhücreler bütün beyine yayılmışlardı fakat hipokampus ve serebellar kortekste daha yoğun şekilde bulunuyorlardı.

41
CA1 bölgesinde stratum lacunosum molekulare nöronları.CA3 bölgesinde strata oriens,radiatum ve lacunosum molakularede ve dentat girusun stratum molakülare kısmında Ntan1 mRNA sı dolaylı olarak daha yüksek düzeylerdeydi. (resim 3a ve 3b sol panel)Ayrıca serebellar korteksteki pürkinje hücreleri de daha yüksek mRNA seviyelerine sahipti.(Şekil 3b sağ panel)
Ayrıca serebellar korteksteki pürkinje hücreleri de daha yüksek mRNA seviyelerine sahipti.(Şekil 3b sağ panel).")
43
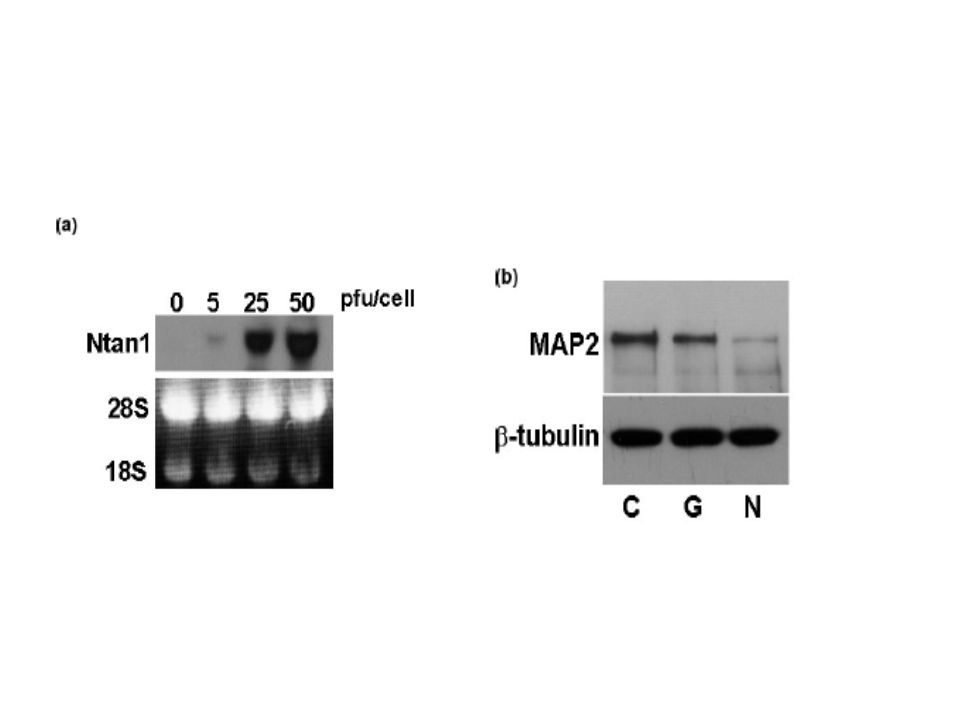
Zayıf manyetizmayla hipokampal nöronlarda Ntan1 ekspresyonu: Daha sonra zayıf manyetizmayla Ntan1 mRNA sı eksprese eden hipokampal nöronları tanımladık.Deneyin onbirinci gününde 15 daklika manyetik alana maruz bırakılan hipokampal nöronların RNA ları 1,3 ve 6 saat sonra alındı ve northern blot yöntemleri kullanılarak analiz edildi.Rat Ntan1 mRNA ekspresyonu yaklaşık 1.4 kb iken 3. ve 6. saatlerde 3 ve 2.5 kat olmak üzere belirgin bir artış gösterdi.

44
Biz ayrıca zayıf manyetik alan uygulamasıyla N1E-115 nöroblastom hücrelerinde de Ntan1 mRNA ekspresyonunu da test ettik.Northern blot zayıf manyetik maruziyetten 3-6 saat sonra,hipokampal nöronlardakine paralel olarak. mRNA düzeylerinde bir up- regulasyon olduğunu gösterdi.

45
Daha sonra Luciferase Reported Analiz yöntemiyle zayıf manyetizmanın Ntan1 promotor aktivitesi üzerine olan etkisini araştırdık.Genomik bilgi bankasındaki rat Ntan1 cDNA dizileribi kullanarak yaklaşık 1.4 kb olan Ntan1 promotor bölgesini tanımladık.1.4kb fare Ntan1 promotorunu taşıyan Luciferase Reported vektörünün N1e-115 nöroblastome hücresine transfeksiyonundan* 1 gün sonra 12 saatlik manyetik maruziyete bırakılan hücreler toplandı.

46
Manyetizmayla beraber luciferase aktivitesi yaklaşık iki kat artmıştı,ve manyetizmanın bilinmeyen bir şekilde Ntan1 promotorunu aktive ettiğini gösteriyordu.

47
Manyetik etkiyle ortaya çıkan Ntan1 artışının sebep olduğu MAP2 ekspresyonunda azalma: Daha önce tekrarlayan manyetik uygulamaların rat hipokampus hücrelerinde MAP2 protein ekspresyonunu azalttığını raporlamıştık. (Hirai and Yoneda 2004).Bu nedenle,bizde burada MAP2 yi Ntan1 bağımlı protein yıkım yolağının bir hedefi olarak test etmeye çalıştık.
.Bu nedenle,bizde burada MAP2 yi Ntan1 bağımlı protein yıkım yolağının bir hedefi olarak test etmeye çalıştık..")
48
Hipokampal nöronlar deneyin 11.gününde 15 dakika statik manyetik alanda bırakıldıktan 6-48 saat sonra MAP2 protein ekspresyonunu western blotting ile belirledik.Manyetik alana hafif maruziyet bile,MAP2 protein ekspresyponunu kontrol ölçümü yapıldıktan 24-48 saat sonra yaklaşık %50 azaltmıştı.

49
a)MAP2 ve beta-tubulin düzeyleri 12-24-48. saatlerde spesifik antikorlar kullanılarak ölçülmüş. d)MAP2 ve beta–tubulin ekspresyonları dansitografi ile değerlendirilmiş.
MAP2 ve beta–tubulin ekspresyonları dansitografi ile değerlendirilmiş..")
50
Daha sonra RT-PCr yöntemini kullanarak Manyetik alanda bırakılmış ve bırakılmamış hücrelerde MAP2 mRNA düzeylerini belirledik ve mRNA düzeylerinde herhangi bir değişiklik olmadığını gösterdik.

51
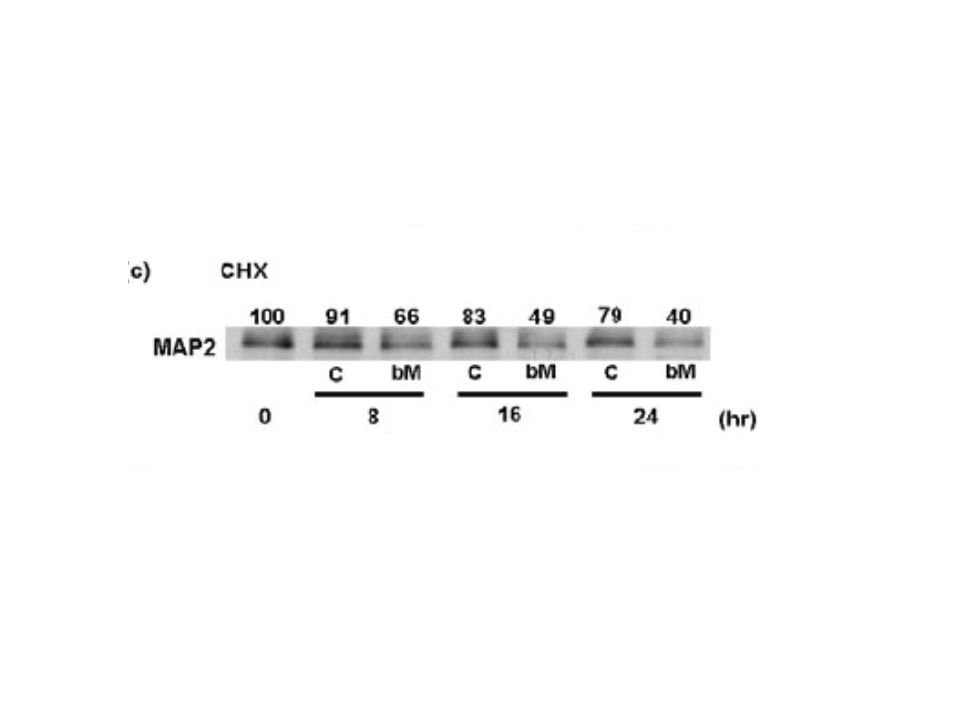
Buda MAP2 ekspresyonunun protein düzeyinde azaltıldığını gösteriyordu. MAP2 protein yıkımını belirlemek için de,bir protein sentez inhibitörü olan CHX(siklohekzimid) maddesini manyetizmanın uygulanmasından 30 dakika önce ekledik.
 maddesini manyetizmanın uygulanmasından 30 dakika önce ekledik..")
52
Yirmi dört saat sonrakontrol grubunda MAP2 protein miktarı ilk andaki değerinin %79una düşmüştü. Manyetik stimülasyon uygulanan grupta ise MAP2 yıkımı daha da kolaylaşmıştı ve 8,16,24. saatlerde ilk andaki düzeyinin %66,%49 ve %40’ına düşmüştü.

54
Buna rağmen rat hipokampus hücrelerine manyetik alan uygulaması yapıldıktan sonraki 48 saatlik zaman dilimi içerisinde hücre yaşayabilirliğinde(survey) ve morfolojisinde bir değişiklik bulunmadı.
 ve morfolojisinde bir değişiklik bulunmadı.")
55
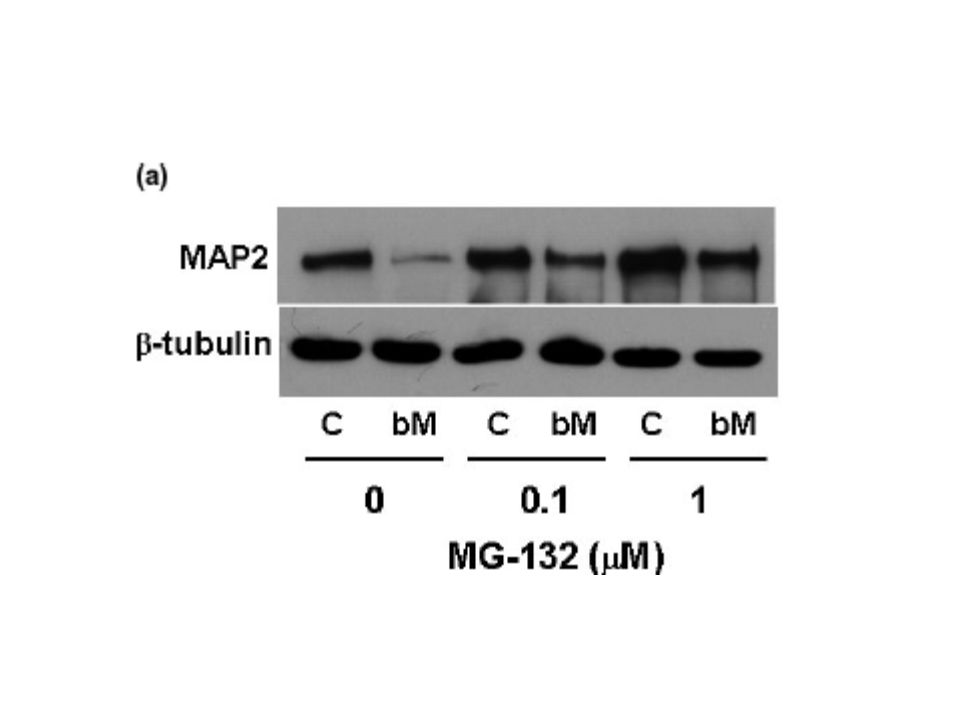
Zayıf manyetik alana maruz bırakıldıktan sonra MAP2 ekspresyonunda ubikitin- proteozom sisteminin rolü olup olmadığını belirlemek için 26S proteozomun selektif inhibitörü olan MG-132’nin etkisini 0,1-1 mikro mol konsantrasyonlarda test ettik.Manyetik uygulamadan 24 saat sonra yapılan ölçümlerle MG-132 eklenmesinin MAP2 protein yıkımındaki azalmayı engellediğini gösterdik.

57
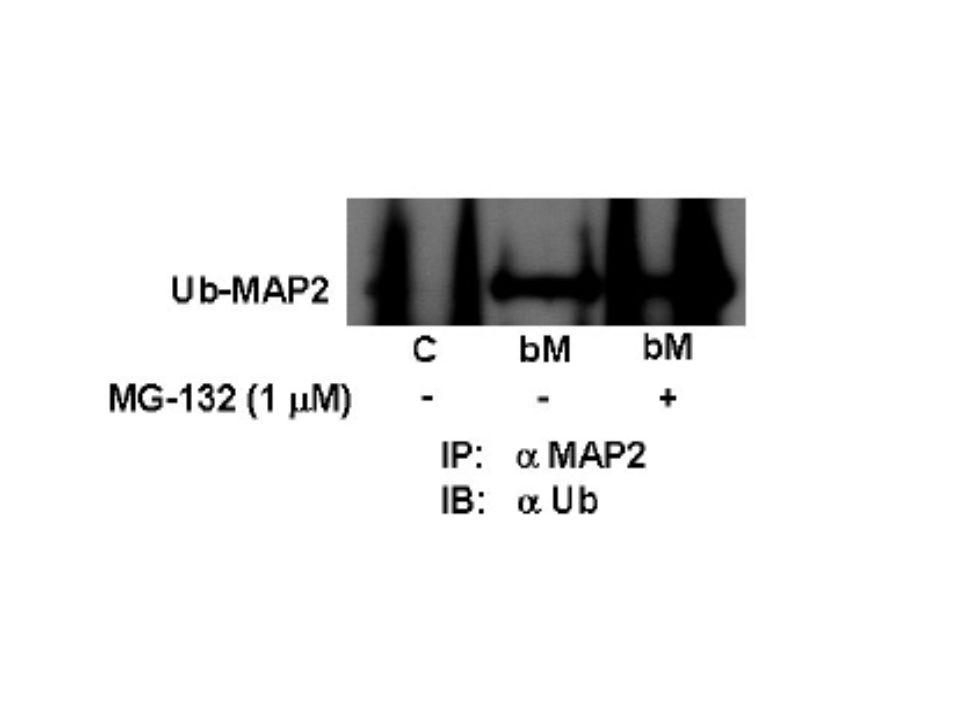
MG-132 konsantrasyonları ile MAP2 düzeyindeki değişim Proteozom inhibitörünün varlığında ve proteozom inhibitörsüz olarak 15 dakika statik manyetik alana maruz bırakılan hipokampal nöronlar fazladan bir 24 saat daha kültürde tutuldular ve bunu takiben hücre lisatı anti-MAP2 antikorlarıyla immunopresipitasyon için hazırlandı.

58
Bu immunopresipitatlarda anti-ubikitin antikorların kullanıldığı western blot analizleriyle ubikitinlenmiş MAP2(Ub- MAP2) düzeyinin belirlenmesi amaçlandı. Zayıf manyetizma maruziyeti hücrelerdeki ubikitinlenmiş MAP2 seviyesinin yükselmesiyle sonuçlandı.1 mikromol MG- 132 varlığında olan ubikitinlenmiş MAP2 değerleri daha fazla arttı.

60
Kantitatif sonuçlar, manyetik alanda bırakılmış hipokampal nöronlarda MAP2 düzeylerinde MG-132 tarafından engellenebilen belirgin bir düşüş ortaya koydu. Bu sonuçlarla manyetik maruziyetten sonra MAP2 protein yıkımının proteosome kaynaklı olduğunu söyleyebiliriz.

61
Daha sonra adenovirus Ntan1 vektörünün transfer edildiği rat hipokampal hücrelerinde ki Ntan 1 in manyetizmanın etkilediği MAP2 yıkımını etkileyip etkilemediğiydi.

62
Doz bağımlı bir şekilde,adenovirus Ntan 1 geniyle enfeksiyondan 48 saat sonra ntan1 ekspresyonu northern blot ile belirlendi.Ntan 1 ekspresyonu infekte hipokampal hücrelerde hücre yaşamsallığını etkiledmeden MAP2 protein seviyelerinde belirgin bir azalmaya sebep oldu.yrıca Ntan1 artışı ubikitinlenmiş MAP2 düzeylerinde dramatik bir artışla sonuçlandı.

64
Bu analizler hipokampal nöronlarda manyetizma tarafından oluşturulan MAP2 yıkımının Ntan1 vasıtasıyla yapıldığını ve MAP2’nin Ntan1 bağımlı bir protein yıkım yolağının hedefi olduğunu gösteriyor.

65
TARTIŞMA Bu çalışmada, DD analizlerini kullanarak rat hipokampüs nöron kültürlerinde statik manyetik alan uygulamasına cevap olarak Ntan1 genini tanımladık. Zayıf manyetizmadan sonraki MAP2 yıkımının proteozome bağımlı olduğunu kanıtladık.

66
Ubikitin proteolitik yolu sinyal iletimi, hücre döngüleri arası geçişler, DNA tamiri, apoptoz ve gen ekspresyonu gibi birçok hücresel sürecin düzenlenmesinde rol alır.(Varshavsky 1997; Yangand Yu 2003).
.")
67
N-End Rule yolağı ubikitin yolağının bir alt gurubudur ve in vivo ortamda bir proteinin yarı ömrü ile bağlantılıdır. Nterminal asparajin ve glutamin sırasıyla Ntan1 ve Ntaq1 tarafından deaminasyona uğratılır.Nterminal aspartat ve glutamat daha sonra ATE1 tarafından arjinin kullanılarak konjuge edilir.

68
N-terminal arjinin ve diğer destabilizan residüler de direkt olarak UBR1 ve N-end rule yolağının bir komponenti olan E3 ile bağlantılıdır. Bu bağlantıda E3’ün tip 1 ve tip 2 substrat bağlanma bölgeleri vasıtasıyla kurulur.Hedef proteinler daha sonra 26 S proteozom tarafından yıkıma uğratılır.

69
MAP2 deki N terminal amino asit asparajin olmamasına rağmen muhtemelen MAP2 proteozom için bir N-terminal asparajin ortaya çıkana kadar hücre içindeki farklı proteazlar tarafından yıkılarak N-end Rule yolağı için bir substrat haline getiriliyor.

70
Son zamanlarda amino ucu ubikitinlenmiş substratlar tanımlandı.Apoptozun düzenlenmesinde vazgeçilmez bir süreç olan,apoptoz inhibitörü drosophilanın yıkımı N- end rule yolağıyla gerçekleştirilir. (Ditzel et al. 2003; Varshavsky 2003).insan lenfositlerinin radyasyon indüksiyonlu yıkımı da Nend rule yolağı tarafından poli(ADP riboz) polimeraz vasıtası ile gerçekleştirilir. (Masdehors et al. 2000).
.insan lenfositlerinin radyasyon indüksiyonlu yıkımı da Nend rule yolağı tarafından poli(ADP riboz) polimeraz vasıtası ile gerçekleştirilir. (Masdehors et al. 2000)..")
71
Bizim bulgularımızda MAP2 nin zayıf manyetizma tarafından Nend rule yolağıyla yıkımı gerçekleştirilen bir substrat olduğunu gösteriyor. Bilgilerimize göre,bu bilgi rat hipokampus hücrelerinde zayıf manyetik alana maruz bırakılmakla ortaya çıkan Ntan 1 cevabının gösterilmesindeki ilk kanıttır.

72
Bu çalışmada,bundan başka,MAP2 Nend rule üzerindeki ubikitin proteosome yolağında ilk defa bir substrat oalrak tanımlanmıştır. Nöronal hücre iskeletinin bir komponenti olan mikrotübüller hücre maturasyonunda önemli bir rol oynar. (Hirokawa1994)Dentritik mikrotübüllerin selektif stabilizasyonu vasıtasıyla gerçekleşen dentritik büyümede hayati önemi olan MAP2, dentritlerde bulunan mikrotübüllerdeki çapraz köprülerin oluşumunda önemli bir komponenttir. (Harada et al. 2002)
Dentritik mikrotübüllerin selektif stabilizasyonu vasıtasıyla gerçekleşen dentritik büyümede hayati önemi olan MAP2, dentritlerde bulunan mikrotübüllerdeki çapraz köprülerin oluşumunda önemli bir komponenttir. (Harada et al. 2002).")
73
Ve MAP2 nöronsal plastisiteyi de içeren dentritlerin rekonstrüksiyonunda önemli bir rol oynayabilir. (Sanchez et al. 2000).Manyetik stimülasyonla MAP2 düzeylerinin azaltıldığı nöronlarda morfolojik olarak bir değişiklik göremedik.MAP2-/- olan sıçan beyin hücrelerinde de major hücre iskeletinde ve hücre yapısında belirgin bir değişiklik gözlemlemedik.
.Manyetik stimülasyonla MAP2 düzeylerinin azaltıldığı nöronlarda morfolojik olarak bir değişiklik göremedik.MAP2-/- olan sıçan beyin hücrelerinde de major hücre iskeletinde ve hücre yapısında belirgin bir değişiklik gözlemlemedik..")
74
Bundan dolayı manyetik stimülasyonla MAP2 protein ekspresyonu azalmış olan hipokampal nöronlardaki normal morfoloji, kısmi fonksiyonel bozulmayla beraber olabilir İn vivo ve in vitro (Marx et al. 2001; Bouras et al. 2001) olarak(Whitaker- Azmitia et al. 1995; Mazer et al.1997), bipolar bozukluk ve şizofreni tanısında MAP2 ekspresyonunun kullanışlı bir belirteç olabileceğine dair kanıtlar birikiyor
 olarak(Whitaker- Azmitia et al. 1995; Mazer et al.1997), bipolar bozukluk ve şizofreni tanısında MAP2 ekspresyonunun kullanışlı bir belirteç olabileceğine dair kanıtlar birikiyor.")
75
Son zamanlarda, şizofreni hastalarının dorsal prefrontal korteks mikroarray* cDNA çalışmalarıyla ubikitin metabolizmasınıda içeren genlerin (ubiquitin specific protease 9, ubiquitin- specific protease 14 and ubiquitin C- terminal esterase L1) ekspresyonunda azalmalar bildirildi. (Middleton et al.2002; Vawter et al. 2002)
.")
76
Ubikitin yolağının sinaptik biçimlenme ve aktivitede önemli bir rol oynamasından dolayı (Hegde et al. 1997; DiAntonio et al.2001),bu moleküllerin azalmış ekspresyonu şizofrenide hücreler arası iletişimi etkileyebilir. İlginç olarak Ntan1 genleri bölünmüş olan fareler açık alanlarda daha az hareket ediyor ve birkaç basit uzamsal hafıza testinde bozukluklar gösteriyor. (Kwon et al. 2000; Balogh et al. 2000).
,bu moleküllerin azalmış ekspresyonu şizofrenide hücreler arası iletişimi etkileyebilir. İlginç olarak Ntan1 genleri bölünmüş olan fareler açık alanlarda daha az hareket ediyor ve birkaç basit uzamsal hafıza testinde bozukluklar gösteriyor. (Kwon et al. 2000; Balogh et al. 2000)..")
77
Bunlar bir bütün olarak ele alındığında Zayıf manyetizmanın indüklediği ubikitin- proteosome yolağının gerçekleştirdiği MAP2 protein yıkımı, rTMS’ye duyarlı olan psikiyatrik hastalıkların tedavisinin mekanizmaları arasında olabilir.

78
Ntan1 in zorlu ekspresyonunu hipokampal nöronlarda MAP2 düzeylerini azalttığını gösterdik.MAP2 yıkımı için destabilizan rezidüleri tam olarak açıklayamadık. Bu yolaktaki destabilizan rezidüleri substratlarını tanımlayan sadece birkaç durum var. (e.g.budding yeast Cup9 and Drosophila IAP1; Turner et al.2000; Ditzel et al. 2003)
.")
79
İlginç olarak her iki substratta da destabilizan rezidüler N terminali yerine internal kesimlerde.MAP2nin N terminal amino asit rezidüsü asparajin olmamasına rağmen, MAP 2 nin diğer proteazlarla yıkıma uğradıktan sonra Nend rule yolağı tarafından yıkılabileceğini göz ardı etmemek gerekir.

80
Sonuçlarımız zayıf manyetizmanın, hipokampüs hücre kültürlerinde Ntan1 ekspresyonunuda içeren ubikitin proteosome yolağı vasıtasıyla nöronal integrasyon ve fonksiyonlarda uzun dönemli değişikliklere yol açtığını gösteriyor.Bu çalışma ayrıca fare Ntan1 geniyle %90 üzerinde homolojiye sahip rat Ntan1 genini ilk defa klonlayıp,tanımlayarak öne çıkıyor.

81
Bütün bunlar beraber ele alındığında,statik manyetizma fizyolojik ve patolojik durumlarda Ntan1 gen ekspresyonu vasıtasıyla nöronal fonksiyonları ayarlayabilir.Altta yatan mekanizmalar üzerinde yapılan daha ileri çalışmalar nöropsikiyatrik bozukluklardan dolay ızdırap çeken birçok hastanın tedavisinde büyük yararlar sağlayabilir.

Benzer bir sunumlar











 Yenelim! Dr Pavel Ursu Dünya Sağlık Örgütü Türkiye Temsilcisi.>")



>")



” “Yaşlılara Doğru Bir Adım Projesi” Dr. Halis YEŞİL Çıraklık ve Yaygın Eğitim Genel Müdürlüğü.>")
>")

