Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Tarihçe, genel özelikler. Adlandırılmaları,koenzim,kofaktör.
ENZİMLER Tarihçe, genel özelikler. Adlandırılmaları,koenzim,kofaktör. Etkili oldukları bölgelere göre sınıflandırma Hız,aktivite, Kinetik Yrd.Doç.Dr.V.Kenan ÇELİK

2
TARİHÇE 1800 yılların başında “sütün ekşimesi, şekerin alkole fermantasyonu gibi” olayların canlı organizmalar aracılığı ile oluştuğu düşünülürdü. 1833’de şekeri parçalayan aktif madde kismi olarak saflaştırıldı ve DİASTAZ (şimdi amilaz)olarak adlandırıldı.Gastrik sıvıdan PEPSİN izole edildi. 1850’lerde LOUİS PASTEUR maya tarafından şekerin alkole dönüşmesinde “FERMENTLER” diye tanımladığı ve canlı organiz-mada bulunan maddeler yardımı ile oluştuğunu ileri sürdü. 1878’ de ilk olarak KÜHNE tarafından ENZİM terimi önerildi. Enzim yunanca da “maya’da” anlamı taşıyan enzume (έγξνμη) den gelmektedir. 1897’de EDUARD BUCHNER hiç canlı hücre içermemesine rağmen maya hücresi özütü eklendiğinde şekerin fermante olduğunu göstermesi bu görüşü ortadan kaldırmıştır. 1926’da SUMMER soyadan ÜREAZ’ı kristallendirmiştir.
olarak adlandırıldı.Gastrik sıvıdan PEPSİN izole edildi. 1850’lerde LOUİS PASTEUR maya tarafından şekerin alkole dönüşmesinde FERMENTLER diye tanımladığı ve canlı organiz-mada bulunan maddeler yardımı ile oluştuğunu ileri sürdü. 1878’ de ilk olarak KÜHNE tarafından ENZİM terimi önerildi. Enzim yunanca da maya’da anlamı taşıyan enzume (έγξνμη) den gelmektedir. 1897’de EDUARD BUCHNER hiç canlı hücre içermemesine rağmen maya hücresi özütü eklendiğinde şekerin fermante olduğunu göstermesi bu görüşü ortadan kaldırmıştır. 1926’da SUMMER soyadan ÜREAZ’ı kristallendirmiştir.")
3
Sonra ki yıllarda saflaştırılan ve kristal yapıları incelenen enzimlerin çoğunun protein (RNA molekülleri (ribozim) hariç,Thomas Cech ve Sydney Altman 1982, 1989 Nobel kimya) yapısında oldukları belirlenmiştir. Ribozimler ribozomlarda peptit transferi ve splicezomda intron transferinden sorumludur.

4
Günümüzde enzim hücreden çıkmış günlük ve ekonomik hayata girmiştir (1 milyar dolar)
")
5
Lipazlar: Deterjan end
Proteazlar:besin end (papain) Karbohidrazlar besin end Tekstil (Selülaz) Farmakoliji (ilaç,) Diagnostik
 Karbohidrazlar besin end. Tekstil (Selülaz) Farmakoliji (ilaç,) Diagnostik.")
6
ENZİMLER NE YAPAR? ∆G (-)
G (serbest enerji) Gibbs’in geliştirdiği bir değer. ∆G=∆H - T∆S ∆G > 0 ise endergonik,tep istemsiz ∆G < 0 ise ekzergonik, tep istemli ∆G = 0 ise tep dengede
 Gibbs’in geliştirdiği bir değer. ∆G=∆H - T∆S. ∆G > 0 ise endergonik,tep istemsiz. ∆G < 0 ise ekzergonik, tep istemli. ∆G = 0 ise tep dengede.")
7
Enzimler: Biyolojik sistemlerde kimyasal tepkimeleri katalizleyen ve tepkime sonunda yapıları değişmeyen moleküllerdir.Katalitik aktivite göstermeleri için yardımcı faktörlere gereksinirler. Enzimlerin aktivite göstermeleri için gerekli olan ve protein yapısında olmayan,genellikle metal iyonlarından oluşan yan gruplara “KOFAKTÖR” denilir. Tablo 1

8
Kofaktör olarak Zn2+ (karbonik anhidraz)
")
9
Enzimlerin aktivite göstermeleri için gereksinim duyduğu kompleks organik moleküllere KOENZİM denir. Özgül atomların ve işlevsel grupların geçici taşıyıcıları olarak görev yapan koenzimler: Koenzim Transfer edilen grup Besinsel öncülü Biositin CO2(yağ asi.sen) Biyotin (0.3 mg kc,yum.fıs) KoenzimA (CoA) Açil grupları Pantotenik asit (Vit B 5) (aa/lipidlerin Glu.dönüş.) (10 mg,Salmon,kc,yum,tahıl ) Flavin Adenin Elektron Riboflavin (Vit B2) Dinükleotid.FADH (1.7 mg kc,tavuk,yum,yeş.seb) Nikotinamid Adenin Hidrit iyonu (:H-) Nikotinik asit (Niasin,Vit B3) Dinükleotid NADH ,NADPH (13-18 mg et,maya,kah.ren.prinç) Pridoksal Fosfat PLP Amino grup Pridoksin (Vit B6) (2 mg, et,balık,ıspanak,fındık,fıstık) Tetrahidro Folat THF Tek C’lu grup Folat (folik asit) (0.4 mg yeşil lifl sebz,kuru bakla, et, kuşkonmaz,hubu) Tiamin pirofosfat TPP Aldehit Tiyamin (Vit B1) 2 mg, kc,süt,tahıl Lipoat ê, ve açil grup diyette şart değil 5’-deoksi kobalamin H , alkil grup Vit B12 3 µg et,süt,yoğurt
 Biyotin (0.3 mg kc,yum.fıs) KoenzimA (CoA) Açil grupları Pantotenik asit (Vit B 5) (aa/lipidlerin Glu.dönüş.) (10 mg,Salmon,kc,yum,tahıl ) Flavin Adenin Elektron Riboflavin (Vit B2) Dinükleotid.FADH (1.7 mg kc,tavuk,yum,yeş.seb) Nikotinamid Adenin Hidrit iyonu (:H-) Nikotinik asit (Niasin,Vit B3) Dinükleotid NADH ,NADPH (13-18 mg et,maya,kah.ren.prinç) Pridoksal Fosfat PLP Amino grup Pridoksin (Vit B6) (2 mg, et,balık,ıspanak,fındık,fıstık) Tetrahidro Folat THF Tek C’lu grup Folat (folik asit) (0.4 mg yeşil lifl. sebz,kuru bakla, et, kuşkonmaz,hubu) Tiamin pirofosfat TPP Aldehit Tiyamin (Vit B1) 2 mg, kc,süt,tahıl. Lipoat ê, ve açil grup diyette şart değil. 5’-deoksi kobalamin H , alkil grup Vit B12 3 µg et,süt,yoğurt.")
10
Koenzim fonksiyonu:

11
Bazı durumlarda enzim aktivite göstermek için hem kofaktöre hemde koenzime gereksinim duyabilir ve enzime kovalent olarak bağlana-bilir, bu durumda diyaliz ile uzaklaştırılmaları mümkün olmaz.Enzim yüzeyine sıkıca bağlanmış ve protein yapısında olmayan bu grup-lara “PROSTETİK GRUP” denir. Karboksilaz-Biotin (Lys’e bağlı) Eğer enzim koenzim veya kofaktörü ile birlikte ve katalitik bakımdan aktif durumda ise enzimin bu haline “HOLOENZİM” denir. Koenzim ve kofaktör enzimden ayrılacak olur ve enzim inaktif hale gelirse, diyalizle ayrılmayan yalnız proteinden oluşan enzimin inaktif şekline “APOENZİM” denir.
 Eğer enzim koenzim veya kofaktörü ile birlikte ve katalitik bakımdan aktif durumda ise enzimin bu haline HOLOENZİM denir. Koenzim ve kofaktör enzimden ayrılacak olur ve enzim inaktif hale gelirse, diyalizle ayrılmayan yalnız proteinden oluşan enzimin inaktif şekline APOENZİM denir.")
12
Enzimleri diğer katalizörlerden ayıran 3 önemli özellik:
1- Son derece hızlı çalışırlar: CO H2O H2CO3 (çok yavaş) (Normal) Karbonik anhidraz ,3x (sn’de) Β-amilaz “ DNA polimeraz “ Laktat Dehidrogenaz “ Karboksi peptidaz A x Üreaz Fosfogluko mutaz
 (Normal) Karbonik anhidraz 1,3x (sn’de) Β-amilaz DNA polimeraz 900. Laktat Dehidrogenaz Karboksi peptidaz A 3x Üreaz Fosfogluko mutaz")
13
2- ÖZGÜLDÜRLER (KEMOSELEKTİF-STERO)
Tüm enzimler Özgül değildir

14
1 mol Hidrojen peroksit’in kendi kendine yıkılması için 18000 kal
3-Biyokimyasal reaksiyonları düşük enerji ve vücut ısısında gerçekleştirirler: 2 H2O H2O O2 1 mol Hidrojen peroksit’in kendi kendine yıkılması için kal Enerjiye ihtiyaç vardır. KATALAZ bu reaksiyonu 2000 kal’ik Enerji ile yapabilmektedir.

15
ENZİMLERİN ADLANDIRILMASI:
Enzimler genellikle katelizledikleri reaksiyonların veya substratın sonuna “AZ” eki getirilerek adlandırılır. Ör. Üreaz , Fosfataz , Laktaz gibi Çok azda “İN” takısı getirilerek adlandırılan özel enzimler vardır. Bunlar genellikle proteolitik enzimlerdir. (Pepsin, Kimotripsin, Tripsin) IUB (Uluslar arası Biyokimya birliği) tarafından enzimler yaptıkları işe göre 6 sınıfa ayrılmıştır.
 IUB (Uluslar arası Biyokimya birliği) tarafından enzimler yaptıkları işe göre 6 sınıfa ayrılmıştır.")
16
ENZİMLERİN SINIFLANDIRILMASI:
OKSİDO-REDÜKTAZLAR TRANSFERAZLAR HİDROLAZLAR LİYAZLAR İZOMERAZLAR LİGAZLAR EC:Enzim sınıfını belirtir. (Oks…,Trans….,Hidrol…) EC:( ) sınıfı Alt sınıfı (Etkilediği bağ) Alkoldehidrogenaz Enzimin bu gruptaki sıra numarası Akseptör (OH, P,NADH)
 EC:( ) sınıfı. Alt sınıfı (Etkilediği bağ) Alkoldehidrogenaz. Enzimin bu gruptaki sıra numarası. Akseptör (OH, P,NADH)")
17
Redoks tepkimelerini katelizlerler.
1-OKSİDO-REDÜKTAZLAR Redoks tepkimelerini katelizlerler. Koenzimler (Donor-Akseptör) NADH,NADPH;FADH2 Dehidrogenazlar,Oksidazlar, Redüktazlar. CH3-CH-COO- ll O Piruvat CH3-CH-COO- │ OH Laktat NADH + H+ NAD+ + + Donor Akseptör Laktat Dehidrogenaz
 NADH,NADPH;FADH2. Dehidrogenazlar,Oksidazlar, Redüktazlar. CH3-CH-COO- ll. O. Piruvat. CH3-CH-COO- │ OH. Laktat. NADH + H+ NAD+ + + Donor. Akseptör. Laktat Dehidrogenaz.")
18
Koenzimleri: TPP,THF,PLP,Lipoik asit, Vit.B12, CoASH
2-TRANSFERAZLAR CH2;NH2;PO4,S gibi grupları transfer eder. Koenzimleri: TPP,THF,PLP,Lipoik asit, Vit.B12, CoASH AST (SGOT), ALT (SGPT), KİNAZLAR GLUKOZ ATP Glukoz-6-Fosfat ADP + + Hekzokinaz EC: 2.1→ Tek Karbon 2.2→Aldehit, Keton 2.3→Açil 2.1.1→Hidroksimetil 2.1.1→Formil
, ALT (SGPT), KİNAZLAR. GLUKOZ. ATP. Glukoz-6-Fosfat. ADP. + + Hekzokinaz. EC: 2.1→ Tek Karbon. 2.2→Aldehit, Keton. 2.3→Açil →Hidroksimetil →Formil.")
19
C-C, C-N, C-P, C-S gibi bağlara H20 katarak yıkımı katelizler.
3- HİDROLAZLAR C-C, C-N, C-P, C-S gibi bağlara H20 katarak yıkımı katelizler. Genelde kofaktör kullanırlar Proteazlar, ester hidrolazlar. NH2-C-NH2 ll O Üre H2O CO2 2 NH3 Ni2+ + + ÜREAZ

20
PİRUVAT DEKARBOKSİLAZ
4-LİYAZLAR C-C, C-S, C-O ve belirli C-N bağlarını yıkıma uğratırlar. Dekarboksilazlar, Dehidratazlar, Sentazlar (tersinir rea) O ll CH3-C-COO- Piruvat O ll CH3-CH Asetaldehit CO2 H+ + + PİRUVAT DEKARBOKSİLAZ
 O. ll. CH3-C-COO- Piruvat. O. ll. CH3-CH. Asetaldehit. CO2. H+ + + PİRUVAT DEKARBOKSİLAZ.")
21
Rasemazlar, mutazlar, epimerazlar,izomerazlar
Optik ve geometrik izomerlerin rasemizasyonunu katelizlerler.Molekül ağırlığı değişmez. Rasemazlar, mutazlar, epimerazlar,izomerazlar O ll -OOCCH2-C-CoA Süksinil CoA 3HC O l ll -OOC-CH-C-CoA Metilmalonil CoA MMCoA MUTAZ

22
Sentetaz’lar olarakta kullanılır (Tersinmez reak)
6-LİGAZLAR ATP’nin hidrolizi ile C-C, C-S, C-O,ve C-N arasında bağ oluşumunu katelizler. Sentetaz’lar olarakta kullanılır (Tersinmez reak) Tirozin-tRNA-Ligaz, Karboksilazlar ATP ADP +Pi O ll CH3-C-COO- Piruvat O ll HOOC-CH2-C-COO- Okzaloasetat CO2 + Piruvat karboksilaz
 Tirozin-tRNA-Ligaz, Karboksilazlar. ATP. ADP +Pi. O. ll. CH3-C-COO- Piruvat. O. ll. HOOC-CH2-C-COO- Okzaloasetat. CO2. + Piruvat karboksilaz.")
23
DİAGNOSTİK (TEŞHİS) ENZİMOLOGİ
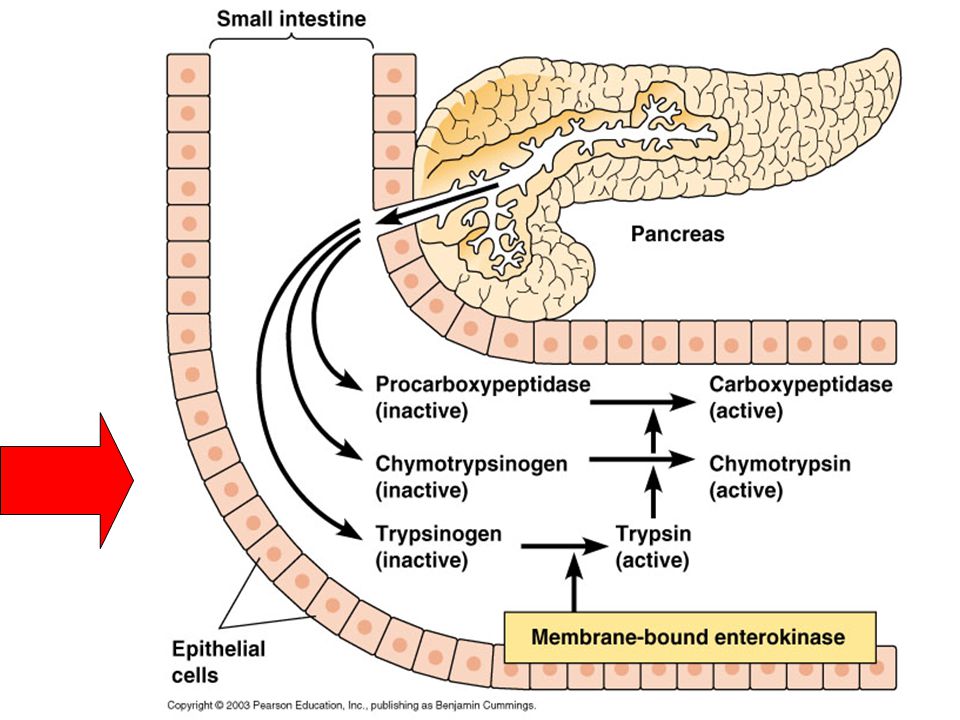
Enzimlerin etkili oldukları yere göre sınıflandırma: Plazma özgü (spesifik) enzimler: Serin proteaz, Plazmalojen, Prokoagülanlar Sekresyon Enzimleri: Salgılandıkları hücrenin dışın-da görev yaparlar.Genellikle PRECURSOR (öncül) olarak salgılanırlar, etkili olduğu bölgede aktifleşirler.α-amilaz (tükrük,pankreas), Lipaz, Pepsinojen (Mide) Tripsinojen,Prokarboksipeptidaz,Proelastaz (Pankreas.), Prostetik asit fosfataz Hücresel enzimler: Sadece hücre içinde görev yapar-lar.Dokularda hasar olduğunda plazmaya geçerler.Plazmada belli bir yarılanma ömürleri vardır.
 enzimler: Serin proteaz, Plazmalojen, Prokoagülanlar. Sekresyon Enzimleri: Salgılandıkları hücrenin dışın-da görev yaparlar.Genellikle PRECURSOR (öncül) olarak salgılanırlar, etkili olduğu bölgede aktifleşirler.α-amilaz (tükrük,pankreas), Lipaz, Pepsinojen (Mide) Tripsinojen,Prokarboksipeptidaz,Proelastaz (Pankreas.), Prostetik asit fosfataz. Hücresel enzimler: Sadece hücre içinde görev yapar-lar.Dokularda hasar olduğunda plazmaya geçerler.Plazmada belli bir yarılanma ömürleri vardır.")
25
Glutamat Dehidrogenaz g-Glutamil Trans Peptitaz(GGT)
ENZİM DOKU Klinik Uygulanım Plazma yarı ömürleri (Saat) ALT kc Karaciğer hasarı ve hastalığı 50 AST Kc, kas, böbrek MI, kc, iskelet kası MI 12 Sitoplazmik 20 Mitokondrial 10 Alkalen Fosfataz Kc, kemik, Germ Hüc Kemik oluşumu İnce bar< 1 Kemik 40 Plesenta 170 Amilaz Tükrük bezi, pancreas Pankreas hastalıklarında Creatin Kinaz Tüm kas ve beyin MI, kas CK-3 (MM) 13 CK-2 (MB) 6 CK-1 (BB) 2 Laktat Dehidrogenaz Hemen hemen tüm dokuların sitoplazmasında MI, Hemoliz, Kanser gibi pekçok hastalık LDH-1 113 LDH-5 10 Glutamat Dehidrogenaz kc (Mitokondri) Hepati parankimal hastalıklarda g-Glutamil Trans Peptitaz(GGT) Kc,böbrek Hepatobiliyer Prostate specific antigen prostat Prostat kanseri Enzimler bulundukları Yer bakımından özellik Gösterirler. LDH ve ALT Yanlızca sitoplazmada Bulunur.Unilokuler.AST MDH,İzositrat DH, hem Sito hemde mito. Bulunur Bilokuler enz. denir
 ALT. kc. Karaciğer hasarı ve hastalığı. 50. AST. Kc, kas, böbrek. MI, kc, iskelet kası. MI 12. Sitoplazmik 20. Mitokondrial 10. Alkalen Fosfataz. Kc, kemik, Germ Hüc. Kemik oluşumu. İnce bar< 1. Kemik 40. Plesenta 170. Amilaz. Tükrük bezi, pancreas. Pankreas hastalıklarında. Creatin Kinaz. Tüm kas ve beyin. MI, kas. CK-3 (MM) 13. CK-2 (MB) 6. CK-1 (BB) 2. Laktat. Dehidrogenaz. Hemen hemen tüm dokuların sitoplazmasında. MI, Hemoliz, Kanser gibi pekçok hastalık. LDH LDH Glutamat Dehidrogenaz. kc (Mitokondri) Hepati parankimal hastalıklarda. g-Glutamil Trans. Peptitaz(GGT) Kc,böbrek. Hepatobiliyer. Prostate specific antigen. prostat. Prostat kanseri. Enzimler bulundukları. Yer bakımından özellik. Gösterirler. LDH ve ALT. Yanlızca sitoplazmada. Bulunur.Unilokuler.AST. MDH,İzositrat DH, hem. Sito hemde mito. Bulunur. Bilokuler enz. denir.")
26
İzozimler (izoenzimler)
Aynı reaksiyonu katelizlerler İki yada daha fazla polipeptit zinciri içerirler (Dimer, trimer, tetramer vb) Farklı genlerin ürünleridir. Farklı amino asit dizilimlerine sahiptirler. Dokuya özgüldürler.
 Farklı genlerin ürünleridir. Farklı amino asit dizilimlerine sahiptirler. Dokuya özgüldürler.")
27
LDH- LAKTAT DEHİDROGENAZ LDH iki eşdeğer olmayan alt üniteye sahip tetramer yapıdadır. 11 ve 12. kromozom (LDH5(M4), LDH4(M3H), LDH3(M2H2), LDH2(MH3), LDH1(H4) Hemen hemen tüm dokularda. LDH-1 ve LDH-2 (Kalp kası, böbrek ve eritrosit) LDH-3(Sarılık ve toksik hepatit) LDH-4 ve LDH-5 (Karaciğer, iskelet kası)
 LDH-3(Sarılık ve toksik hepatit) LDH-4 ve LDH-5 (Karaciğer, iskelet kası)")
28
CK- KREATİN KİNAZ (EC:2.7.3.2) CK-1 (BB), CK-2 (MB), CK-3 (MM)
CK-1 izoenzimi genellik-le by-pass operasyon sonrası, Gastroints.has. Prostat, lösemi, akciğer CK-2 MI da (12 saat) CK-3 Merkezi Sinir Sistemi Normal Miyokard infarktüs
 CK-3 Merkezi Sinir Sistemi. Normal. Miyokard. infarktüs.")
29
MİYOKART ENFARKTÜSÜ TAKİBEN SERUM DÜZEYLERİ
TROPONİNLER(T,I), Kas hücrelerin de aktin ile miyozin etkileşimini düzenleyen motor ünite bileşenleridir. Spesifik kardiak izozimleridir 4 saat gibi kısa bir sürede aktivitesi artar.Çok duyarlı immunolojik tekniklerle saptanabilir. HBDH—α-Hydroxybutyrate dehydrogenase
, Kas hücrelerin de aktin ile miyozin etkileşimini düzenleyen motor ünite bileşenleridir. Spesifik kardiak izozimleridir 4 saat gibi kısa bir sürede aktivitesi artar.Çok duyarlı immunolojik tekniklerle saptanabilir. HBDH—α-Hydroxybutyrate dehydrogenase.")
31
ENZİM AKTİF MERKEZİ Aktif bölge İnduced-fit Anahtar-Kilit modeli

32
Anahtar-kilit modeli 1894 Ficher İnduced-fit modeli 1958 Koshlan
Substrat bağlanması, altivasyonu ve reaksiyon enzim aktif merkezinde gerçekleşir. Substrat bağlanma enerjisi 3-12 kcal/mol Anahtar-kilit modeli 1894 Ficher İnduced-fit modeli Koshlan

33
Enzim “AKTİF MERKEZİN” de yer alan amino asitler
Mavi=S’ın cep’e bağlandiği aromatik halka . Mor=Aromatik halkadaki karbonil grubu . Serin (ser) Lizin (lys) Histidin (his) Sistein (Cys) Tirozin (Tyr) Triptofan (Trp) Fenilalanin (Phe) Glutamat (Glu)
 Lizin (lys) Histidin (his) Sistein (Cys) Tirozin (Tyr) Triptofan (Trp) Fenilalanin (Phe) Glutamat (Glu)")
34
Yakınlaştırma ve yönlendirme etkisi
ENZİM KATALİZİ Asit-Baz Kovalent Metal iyon Elektrostatik Yakınlaştırma ve yönlendirme etkisi Geçiş komplekslerinde tercihli bağlanma Çok S’ lı tepkimeler PİNG-PONG (Bİ-Bİ) ya da sıralı (ardıl)- Gelişigüzel bağlanma mekanizmaları gösterir.
 ya da sıralı (ardıl)- Gelişigüzel bağlanma mekanizmaları gösterir.")
35
ASİT-BAZ KATALİZİ

36
KOVALENT KATALİZ Substrat üzerindeki elektrofilik grupla katalist üzerindeki nükleofilik grup arasındaki kovalent bağ geçişini hızlandırırlar.

38
METAL İYON Karbonik Anhidraz

39
Elektrostatikkataliz.
Enzim aktif merkezinin hidrofobik veya hidrofilik bir cep oluşturarak (yük dengesi değişikliği) substratın bağlanması. Yakınlaştırma ve yönlendirme. Oldukca önemli enzimler substrat üzerinde oranyantasyon ve rotasyon etkisi ile reaksiyonun hızlı bir şekilde gerçekleşmesini sağlar.Bu işlemi geçiş pozisyonunda yapar.
 substratın bağlanması. Yakınlaştırma ve yönlendirme. Oldukca önemli enzimler substrat üzerinde oranyantasyon ve rotasyon etkisi ile reaksiyonun hızlı bir şekilde gerçekleşmesini sağlar.Bu işlemi geçiş pozisyonunda yapar.")
40
ENZİM AKTİVİTESİNİ ETKİLEYEN FAKTÖRLER
1-Ortam pH’sı 2-Sıcaklık 3- Enzim konsantrasyonu 4- Substrat “ 5- Zaman 6-Reaksiyon ürünü 7- Çeşitli iyonların derişimleri,özellikleri 8- Işık ve diğer fiziksel faktörlerin etkisi

41
1-ORTAM pH’sı pH = - log [ H+ ]
![1-ORTAM pH’sı pH = - log [ H+ ]](http://slideplayer.biz.tr/slide/3691074/12/images/41/1-ORTAM+pH%E2%80%99s%C4%B1+pH+%3D+-+log+%5B+H%2B+%5D.jpg "1-ORTAM pH’sı pH = - log [ H+ ]")
42
2-SICAKLIK Reaks. Hızı Sıcaklık Maksimum aktivasyon Optimum sıcaklık

43
3- ENZİM KONSANTRASYONU

44
4- SUBSTRAT KONSANTRASYONU
Velocity, v Substrate conc., [S]

45
[S] = [S]0 e-kt »» HIZ (velocity, v) S (substrat) → P (product)
-d [S] d [ P] d [S] v= ——— = ——— ise ——— = v= k. [ S ]n ( n= 1,2,3)derece dt dt dt [S] t [S]o - d[S] = k dt ise 2,3 log —— = k.t [S] [S ] [S]o [S] = [S]0 e-kt »» k log [S] = - —— t log [ S ]o 2,3 y = m x a
![[S] = [S]0 e-kt »» HIZ (velocity, v) S (substrat) → P (product)](http://slideplayer.biz.tr/slide/3691074/12/images/45/%5BS%5D+%3D+%5BS%5D0+e-kt+%C2%BB%C2%BB+HIZ+%28velocity%2C+v%29+S+%28substrat%29+%E2%86%92+P+%28product%29.jpg "-d [S] d [ P] -d [S] v= ——— = ——— ise ——— = v= k. [ S ]n ( n= 1,2,3)derece. dt dt dt. [S] t [S]o. - d[S] = k dt ise 2,3 log —— = k.t. [S] [S ] [S]o 0. [S] = [S]0 e-kt »» k. log [S] = - —— t + log [ S ]o. 2,3. y = m x + a.")
46
MİCHAELİS-MENTEN eşitliği
log[S] MİCHAELİS-MENTEN eşitliği 1902 Brown “ES” kompleks’ ini ileri sürüyor.1903 Henry hız eşitliğini ileri sürüyor. 1913 yılında Michaelis-Menten tarafından hız (v0 ilk hız) ve [S] arasındaki ilişkiyi matematize ederek Km gibi Enzim ve substrat arasındaki ilişkiyi veren sabit bir değer buldular. Böylece Michaelis-Menten eşitliği Enzim miktarı ile ilişkili bir reaksiyonun hızının ölçülmesine olanak sağlamıştır. Log [S]0 Eğim = - k / 2,3 [S] 1/2 t t1/2 Bir reaksiyonun yarı ömrü t1/2 = 0,69 / k
 ve [S] arasındaki ilişkiyi matematize ederek Km gibi Enzim ve substrat arasındaki ilişkiyi veren sabit bir değer buldular. Böylece Michaelis-Menten eşitliği Enzim miktarı ile ilişkili bir reaksiyonun hızının ölçülmesine olanak sağlamıştır. Log [S]0. Eğim = - k / 2,3. [S] 1/ t. t1/2. Bir reaksiyonun yarı. ömrü. t1/2 = 0,69 / k.")
47
Tek S’li bir enzimin katelizlediği reaksiyon
ES k2 E P S + + k-1 3 Temel yaklaşım yapılmıştır; 1- Reaksiyonun başlangıç süresince ES kompleks derişimi (her ne kadar ürün oluş sada) sabittir. [ES]=sbt 2- Tüm E doygunluğa ulaştığında ES kompleksi oluşturulur. Serbest (boş) enzim yoktur. Bu yüksek [S] olur. 3- Tüm enzim “ES” kompleksini oluşturmuşsa, ürünlerin oluşum Hızı da maksimum orandadır. Vmax = k2 [ES]
 sabittir. [ES]=sbt. 2- Tüm E doygunluğa ulaştığında ES kompleksi oluşturulur. Serbest (boş) enzim yoktur. Bu yüksek [S] olur. 3- Tüm enzim ES kompleksini oluşturmuşsa, ürünlerin oluşum. Hızı da maksimum orandadır. Vmax = k2 [ES]")
48
E + S ES E + P k1 k2 k-1 [ES] için voluş = k1[E][ S],
v yıkım = k2 [ES] + k-1 [ES] = [ ES ] (k2 + k-1) Steady-steate (kararlı hal) yapım = yıkım k1[E][ S] = [ ES ] (k2 + k-1) [E][S] =[ES] (k2 + k-1) k1 [E]=[Et]-[ES] [S] [Et]-[ES] =[ES] Km [S]’ [ES] bölers. [Et] Km + [S] [ES] [S] Km = k2 + k-1 k1 = 1
![E + S ES E + P k1 k2 k-1 [ES] için voluş = k1[E][ S],](http://slideplayer.biz.tr/slide/3691074/12/images/48/E+%2B+S+ES+E+%2B+P+k1+k2+k-1+%5BES%5D+i%C3%A7in+volu%C5%9F+%3D+k1%5BE%5D%5B+S%5D%2C.jpg "v yıkım = k2 [ES] + k-1 [ES] = [ ES ] (k2 + k-1) Steady-steate (kararlı hal) yapım = yıkım. k1[E][ S] = [ ES ] (k2 + k-1) [E][S] =[ES] (k2 + k-1) k1. [E]=[Et]-[ES] [S] [Et]-[ES] =[ES] Km [S]’ [ES] bölers. [Et] Km + [S] [ES] [S] Km = k2 + k-1. k1. = 1.")
49
Et’ ==> ES’ ye eşit olmadığında v= k2 [ES] ==> [ES] = v / k2
Enzim S’ la tamamen dolmuş olsaydı [Et] = [ES] olacaktı. Bu durumda hız da max. Olur. Vmax = k2 [ES] ==> [Et]=Vmax/ k2 Et’ ==> ES’ ye eşit olmadığında v= k2 [ES] ==> [ES] = v / k2 [Et] Vmax / k2 [ES] v/ k2 1’ de yerine konursa Vmax km +[S] v [S] Vmax [S] ya da v = ————— km + [S] Vmax / v = = =
![Et’ ==> ES’ ye eşit olmadığında v= k2 [ES] ==> [ES] = v / k2](http://slideplayer.biz.tr/slide/3691074/12/images/49/Et%E2%80%99+%3D%3D%3E+ES%E2%80%99+ye+e%C5%9Fit+olmad%C4%B1%C4%9F%C4%B1nda+v%3D+k2+%5BES%5D+%3D%3D%3E+%5BES%5D+%3D+v+%2F+k2.jpg "Enzim S’ la tamamen dolmuş olsaydı [Et] = [ES] olacaktı. Bu durumda hız da max. Olur. Vmax = k2 [ES] ==> [Et]=Vmax/ k2. Et’ ==> ES’ ye eşit olmadığında v= k2 [ES] ==> [ES] = v / k2. [Et] Vmax / k2. [ES] v/ k2. 1’ de yerine konursa Vmax km +[S] v [S] Vmax [S] ya da v = ————— km + [S] Vmax / v. = = =")
50
Vmax [S] v = ————— Michaelis-Menten hız denklemi km + [S]
Bir kimyasal reaksiyonun karakterini n (derece) belirtir. v= k [ S ]n 0° hız kinetiği [S] >> km değerinde gerçekleşir.Bu durumda Km ihmal v = k0 dır. Hız eşitliğinde reaktanların (S) derişimine ait bir ifade olmadığından reaktan ilavesi hızı artırmaz, hız maksimum hızda başlar ve sabittir. v 1° hız kinetiği [S] << Km değerinden Olduğundan hız eşitliğinde ihmal edilir.Bu nedenle v= k.[S] dir v [S] 2° hız kinetiğinde v=k [A]1[B]1=[S]2 V [S]
![Vmax [S] v = ————— Michaelis-Menten hız denklemi km + [S]](http://slideplayer.biz.tr/slide/3691074/12/images/50/Vmax+%5BS%5D+v+%3D+%E2%80%94%E2%80%94%E2%80%94%E2%80%94%E2%80%94+Michaelis-Menten+h%C4%B1z+denklemi+km+%2B+%5BS%5D.jpg "Bir kimyasal reaksiyonun karakterini n (derece) belirtir. v= k [ S ]n. 0° hız kinetiği [S] >> km değerinde gerçekleşir.Bu durumda Km ihmal. v = k0 dır. Hız eşitliğinde reaktanların (S) derişimine ait bir ifade olmadığından reaktan ilavesi hızı artırmaz, hız maksimum hızda başlar ve sabittir. v. 1° hız kinetiği [S] << Km değerinden. Olduğundan hız eşitliğinde ihmal. edilir.Bu nedenle v= k.[S] dir. v. [S] 2° hız kinetiğinde v=k [A]1[B]1=[S]2. V. [S]")
51
R-C-O-CH3 (aq) + H2O (s) R-C-OH (aq) + CH3OH (aq) v = k [Ester][H2O]
2° kinetiği; İki S’ ın olduğu (Su yada herhangi bir reaktan ) büyük çoğunluğu “pseudo-first-order” (yalancı 1° reaksiyon) olarak adlandırılır. O O ll ll R-C-O-CH3 (aq) + H2O (s) R-C-OH (aq) + CH3OH (aq) Ester Asit Alkol v = k [Ester][H2O] Saf sıvılar ve katılar “denge sabitinde yer almaz” . Su için Kden = 55.5 , Ester derişimi ( 10-3, 10-2 M)(0,003) Bu yüzden 1° kinetiğine uyar.Biyolojik sistemlerde oluşan böylesi reaksiyonlar genelde Hidratasyon, Dehidratasyon ve Hidroliz reaksiyonlardır.
![R-C-O-CH3 (aq) + H2O (s) R-C-OH (aq) + CH3OH (aq) v = k [Ester][H2O]](http://slideplayer.biz.tr/slide/3691074/12/images/51/R-C-O-CH3+%28aq%29+%2B+H2O+%28s%29+R-C-OH+%28aq%29+%2B+CH3OH+%28aq%29+v+%3D+k+%5BEster%5D%5BH2O%5D.jpg "2° kinetiği; İki S’ ın olduğu (Su yada herhangi bir reaktan ) büyük çoğunluğu pseudo-first-order (yalancı 1° reaksiyon) olarak adlandırılır. O O. ll ll. R-C-O-CH3 (aq) + H2O (s) R-C-OH (aq) + CH3OH (aq) Ester Asit Alkol. v = k [Ester][H2O] Saf sıvılar ve katılar denge sabitinde yer almaz . Su için Kden = 55.5 , Ester derişimi ( 10-3, 10-2 M)(0,003) Bu yüzden 1° kinetiğine uyar.Biyolojik sistemlerde oluşan böylesi reaksiyonlar genelde Hidratasyon, Dehidratasyon ve Hidroliz reaksiyonlardır.")
52
Mikaelis-Menten grafiği
0° 2[S] = KM + [S] 1° [S] = KM Km , Enzimin substratına Olan ilgisini gösterir.Km ne Kadar küçük ise enzim S’na Olan ilgisi çok fazladır. Km ne Kadar büyükse enzimin S’na İlgisi o kadar azdır.

53
kcat = Vmax/Et kcat, turnover sayısı;,
Birim zamanda enzim molekülü başına ürüne çevrilen S molekülünün maksimum sayısı M-M modeline göre; kcat = Vmax/Et

54
Katalitik verim kcat/Km S derişimi çok düşük olduğunda enzim performansında ki değişiklikleri ölçmek.

55
Molar soğurma katsayısı=mM-1cm-1
TERMİNOLOJİ Standart ünite: (U), Dakikada 1 µmol dönüşümü katalizleyen aktivite miktarıdır. (µmol / dk) Volum aktivite: V (Toplam hacim) VA= ——— x ∆A/∆t (U/ml) ε . d. v. Spesifik aktivite; mg protein başına enzim ünit aktivitesidir. (U / mg protein.) V Spesifik aktivite:————— x ∆A/∆t ε . d. v. Cprotein A=ε.d.C Molar derişim Molar soğurma katsayısı=mM-1cm-1 Kuvet kalınlığı (cm)
, Dakikada 1 µmol dönüşümü katalizleyen aktivite miktarıdır. (µmol / dk) Volum aktivite: V (Toplam hacim) VA= ——— x ∆A/∆t (U/ml) ε . d. v. Spesifik aktivite; mg protein başına enzim ünit aktivitesidir. (U / mg protein.) V. Spesifik aktivite:————— x ∆A/∆t. ε . d. v. Cprotein. A=ε.d.C. Molar derişim. Molar soğurma katsayısı=mM-1cm-1. Kuvet kalınlığı (cm)")
56
Lineweaver-Burk

57
Glucose + ATP glucose-6-phosphate + ADP

58
Eadie-Hofstee eğrisi (M-M’e göre)
v Vmax Slope=-Km Vmax/Km v/[S]

59
Hanes-Wolff eğrisi (Lineweaver-burk’e göre)
[S]/v Km/vmax -Km Slope=1/vmax

60
ENZİM İNHİBİSYONLARI Enzimler inhibitörler tarafından reversibl (tersinir) ve irreversibl (tersinmez) olarak inhibisyona uğrarlar: Önemi: Aktiviteleri düzenlenerek kontrol sisteminde etkili olurlar. Enzim fonksiyonlarını inhibe eden ilaç ve zehirli bileşiklerin fonksiyon mekanizmalarının anlaşılması. Substrat analogları kullanarak önemli araştırma olanakları sağlanması. Irreversible inhibitörler; Enzime kovalent olarak bağlanırlar, enzimin katalitik etkisini tamamen durdururlar. Örn. Ağır metaller, Sinir gazı zehirlenmeleri ve bazı insektisitler. Reversible inhibitörler; Kompetetiv inhibitörler (Enzim aktif bölgesine bağlanmak için substratla yarışan),Nonkompetetiv inhibitörler (Enzim aktif bölgesinden başka bir bölgeye bağlanan) .
,Nonkompetetiv inhibitörler (Enzim aktif bölgesinden başka bir bölgeye bağlanan) .")
62
Kompetetiv İnhibisyon
Kompetetiv inhibitör yapı Olarak S’la benzerdir (analog) İnhibitör enzim aktif bölgesi İçin S’la yarışır. S derişimi artırılırsa inhibitör Etkisi azaltılabilir.
 İnhibitör enzim aktif bölgesi. İçin S’la yarışır. S derişimi artırılırsa inhibitör. Etkisi azaltılabilir.")
63
Süksinat dehidrogenazın
Kompetetiv bir inhibitörü Yapı olarak süksinata Benzer Süksinat ilavesi İnhibisyonu tersine Çevirir.

64
Kompetitive Reversible Inhibitörler
Km Artar vmax Değişmez KI Kmapp +inhibitor v 1/v Slope=Km/vmax vmax Slope= Km(1+[I]/KI)/vmax -1/Km 1/vmax Km(1+[I]/KI) [S] 1/[S] Km -1/(Km(1+[I]/KI))
/vmax. -1/Km. 1/vmax. Km(1+[I]/KI) [S] 1/[S] Km. -1/(Km(1+[I]/KI))")
65
Nonkompetetiv İnhibisyon
Yapısı S’ın yapısından farklıdır. Enzim aktif bölgesi dışında Bir bölgeye bağlanır. Enzimin şeklini değiştirerek Aktif bölgeyide etkiler ve S’ın Bağlanması engellenir. S ilavesi inhibisyonu Değiştirmez.

66
Nonkompetitive (Pure) Reversible Inhibitörler
KI’ KI Km Değişmez vmax Azalır +inhibitor 1/[S] 1/v 1/vmax -1/Km Slope=Km/vmax (1+[I]/KI)/Vmax Slope= Km(1+[I]/KI)/vmax v [S] vmax Km Vmax/(1+[I]/KI)
/Vmax. Slope= Km(1+[I]/KI)/vmax. v. [S] vmax. Km. Vmax/(1+[I]/KI)")
67
Unkompetitive Reversible Inhibitörler
Km ve vmax Azalır Eğim değişmez KI’ +inhibitor v 1/v vmax Slope=Km/vmax (1+ KI/[I])/Vmax Slope= Km/vmax Vmax/(1+KI/[I]) -1/Km 1/vmax [S] 1/[S] Km/(1+ KI/[I]) Km - (1+ KI/[I])/Km
/Vmax. Slope= Km/vmax. Vmax/(1+KI/[I]) -1/Km. 1/vmax. [S] 1/[S] Km/(1+ KI/[I]) Km. - (1+ KI/[I])/Km.")
68
Mixed Inhibisyonlar (Inh. E ve ES den ayrışma sabiti farklı olduğunda )
+inhibitor 1/v Slope=Km/vmax -1/Km 1/vmax 1/[S] +inhibitor 1/v Slope=Km/vmax -1/Km 1/vmax 1/[S]

69
Hücreler enzim aktivitesini nasıl düzenler?
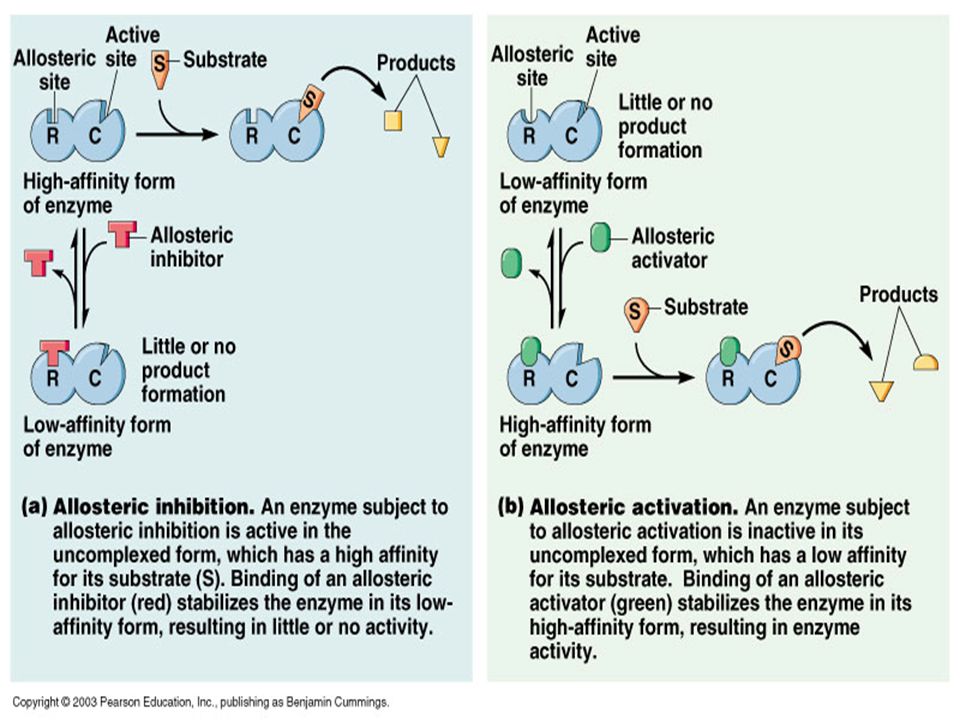
Enzim aktivasyonunun düzenlenmesi Substrat düzeyi regülasyon (G-6-P tarafından HEKZOKİNAZ’ın inhibisyonu Allosterik- Kovalent modifikasyon Enzimlerde iki bağlanma bölgesi vardır. Aktif bölge Regulatör bölge (düzenleyici) Düzenleme LİGAND lar aracılığı ile olur. Enzimi sentezler yada yıkarlar Böylece enzim aktivitesi başlatılır yada sonlandırılır. LİGAND: Efektör(akt.artırıcı),Modülatör(akt.azaltıcı), Modifiyer, Etkilerle enzim aktivitesini değiştiren moleküller. Ligandlar tarafından modülasyona uğrayan enzimler metaboliz mada hız belirleyen enzimlerdir.
 Düzenleme LİGAND lar aracılığı ile olur. Enzimi sentezler yada yıkarlar. Böylece enzim aktivitesi başlatılır yada sonlandırılır. LİGAND: Efektör(akt.artırıcı),Modülatör(akt.azaltıcı), Modifiyer, Etkilerle enzim aktivitesini değiştiren moleküller. Ligandlar tarafından modülasyona uğrayan enzimler metaboliz. mada hız belirleyen enzimlerdir.")
70
S düzeyi inhibisyon

71
Çok küçük [S] (+) efektörler incelenebilir.
Allosterik Efektörler Km ve Vmax üzerinde etkilidirler. Km üzerinde etkili efektörlere K sınıfı Vmax üzerinde etkili efektörlere V sınıfı V sınıfı efektörler: (+) ve (-) allosterik etki ile ES yıkım hızını artırır yada azaltır. Ürün oluşumu ve K2 hız sabiti etkilenir. S’ ın bağlanma bölgesini etkilemez. K sınıfı efektörler: İnhibitör allosterik bölgeye bağlanır ve S bağlanma bölgesinin afinitesini etkiler. Çok küçük [S] (+) efektörler incelenebilir. Çok yüksek [S] (-) efektörler incelenebilir. Buda “metabolik kontrolde” enzim aktivitesinin çok daha Hassas düzenlenmesine olanak verir.
![Çok küçük [S] (+) efektörler incelenebilir.](http://slideplayer.biz.tr/slide/3691074/12/images/71/%C3%87ok+k%C3%BC%C3%A7%C3%BCk+%5BS%5D+%28%2B%29+efekt%C3%B6rler+incelenebilir..jpg "Allosterik Efektörler Km ve Vmax üzerinde etkilidirler. Km üzerinde etkili efektörlere K sınıfı Vmax üzerinde etkili efektörlere V sınıfı. V sınıfı efektörler: (+) ve (-) allosterik etki ile ES yıkım hızını artırır yada azaltır. Ürün oluşumu ve K2 hız sabiti etkilenir. S’ ın bağlanma bölgesini etkilemez. K sınıfı efektörler: İnhibitör allosterik bölgeye bağlanır ve S bağlanma bölgesinin afinitesini etkiler. Çok küçük [S] (+) efektörler incelenebilir. Çok yüksek [S] (-) efektörler incelenebilir. Buda metabolik kontrolde enzim aktivitesinin çok daha. Hassas düzenlenmesine olanak verir.")
73
Allosterik enzimler metabolizmada FeedBack inhi.sağ.
A B C D P E1 E2 E3 E4

74
Çoklu Feedback

75
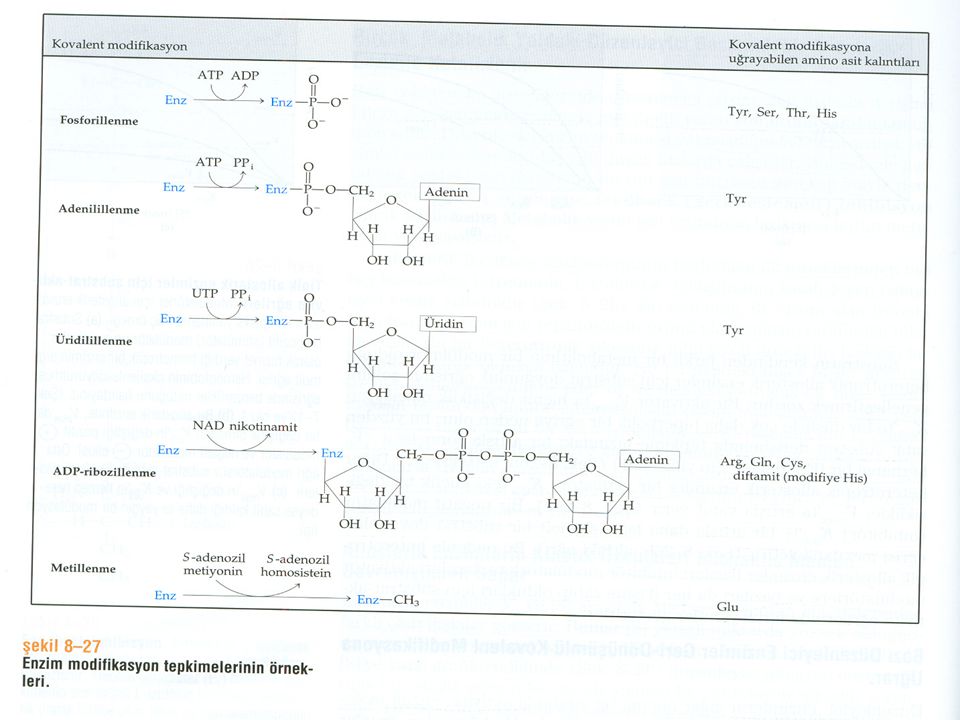
Allosterik enzimler oligomerik yapılar içerir, ligandın bir protomere bağlanması oligomerdeki diğer protomerler üzerine ligandların bağlanmasını etkileyebilir (S-S, Akt-Akt, İnh-İnh ) . Ligandların böylesi etkileri “HOMOTROPİK ETKİLEŞİM” lere neden olurlar. Bu etkileşimler Kooperativ Etkileşimler olarak adlandırılır. “HETEROTROPİK ETKİLEŞİM” ise bir ligandın farklı bir ligandı bağlamasında etkilidir. Kooperativite: Homotropik etkileşimler sonucunda çalışma bölgesi komformasyonal değişeme uğrar.(+) ve (-) etkiler oluşur. Enzimler kimyasal grupların eklenmesi yada çıkarılması ile de düzenlenebilir. KOVALENT MODİFİKASYONLAR olarak adlandırılır. FOSFORİLASYON- DEFOSFORİLASYON GLİKOZİLASYON ASETİLASYON METİLASYON KARBOKSİLASYON UBİQUİTİNASYON MİRİSTİLASYON
 ve (-) etkiler oluşur. Enzimler kimyasal grupların eklenmesi yada çıkarılması ile de düzenlenebilir. KOVALENT MODİFİKASYONLAR olarak adlandırılır. FOSFORİLASYON- DEFOSFORİLASYON. GLİKOZİLASYON. ASETİLASYON. METİLASYON. KARBOKSİLASYON. UBİQUİTİNASYON. MİRİSTİLASYON.")
76
Glikojen sentaz enzimi
Fosforlandığında inaktif Defosforile edildiğinde ise İnaktiftir.

80
ENZİM AKTİVİTE TAYİNİNDE KULLANILAN YÖNTEMLER
Spektrofotometrik ( S, P veya Koenzim) Monometrik (O2,CO2,) Elektrot (asit oluşumu) Polarimetrik (S yada P optikce aktif) Kromatografik Kimyasal tayin yöntemi (kolorimetrik) Flourometrik
 Monometrik (O2,CO2,) Elektrot (asit oluşumu) Polarimetrik (S yada P optikce aktif) Kromatografik. Kimyasal tayin yöntemi (kolorimetrik) Flourometrik.")
81
Catalytic Antibodies: Abzymes
Antibodies are immunoglobulins. Antibodies are elicited in an organism in response to immunological challenge by a foreign molecule called antigens; Antibodies elicited in response to transition state analogs have the ability to stabilize the transition state and thus can catalyze a reaction by forcing the substrate into the transition state structure;

Benzer bir sunumlar