Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
BİYOLOJİK ONARIM MEKANİZMALARI
Yaşayan hücreler çeşitli yollarla DNA hasarlarını onaran enzim sistemlerine sahip olacak biçimde evrimleşmiştir Onarım sisteminin etkin olduğu spontan mutasyonların az olmasından görülebilir Error-free yolak (hatasız onarım yolağı) DNA bazlarını kimyasal olarak değiştiren hasarları onarır Onarım yolağı hasarlı bazı çıkarır ve komplementer diziyi kullanarak normal diziyi restore eder.
 DNA bazlarını kimyasal olarak değiştiren hasarları onarır. Onarım yolağı hasarlı bazı çıkarır ve komplementer diziyi kullanarak normal diziyi restore eder.")
2
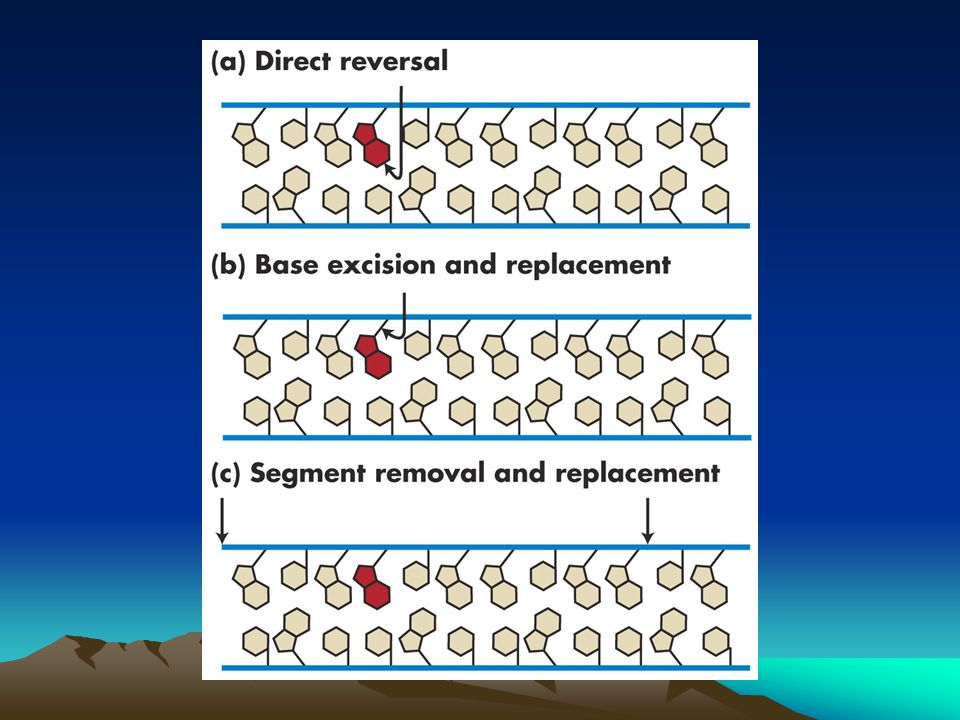
Hasarlı bazın direkt geri dönüşümü
En etkin onarım yolağı lezyonu doğrudan tamir etmektir ve normal baza geri dönüştürmektir. Bazı hasarlar geri dönüşümsüz olmasına karşın bazı durumlarda doğrudan onarım gerçekleşir.bunlardan biri UV ile oluşan fotodimerlerdir.siklobütan pirimidin fotodimerleri fotoliyaz denilen bir enzimle onarılır.enzim fotodimere bağlanır ve dimeri ikiye bölerek orijinal diziye geri dönüştürür. Alkil transferazlar lezyonları direk onaran başka bir enzim grubudur.Bu enzimler O6 guanindeki alkil grubunu uzaklaştırır.Bu alkil grupları EMS ve NG ile eklenirler.

3
Homoloji bağımlı onarım sistemleri
Bilindiği gibi onarım sistemlerinin yararlandığı en önemli özellik baz komplementerliğidir. Bu onarım sistemleri yanlış bazı çıkararak kalıp zinciri kullanıp doğru bazı DNA yapısına katarlar.Bu komplementerlik nedeniyle bu sistemlere homoloji bağımlı onarım sistemleri adı verilir. İki büyük tipi vardır 1. excision onarımı (kesip çıkarma):hasar replikasyondan önce tanınır Baz kesip çıkarma onarımı:baz excision repair Nükleotid kesip çıkarma onarımı:nt excision repair Ökaryotlarda transkripsiyonla birleşen onarım 2.postreplikasyon onarımı:replikasyon sırasında yada sonrasında hasar tanınır.
:hasar replikasyondan önce tanınır. Baz kesip çıkarma onarımı:baz excision repair. Nükleotid kesip çıkarma onarımı:nt excision repair. Ökaryotlarda transkripsiyonla birleşen onarım. 2.postreplikasyon onarımı:replikasyon sırasında yada sonrasında hasar tanınır.")
6
Excision onarım yolakları
Hasarlı bazın direk çıkarılması yerine bu onarım sistemi bazı tamamen keser çıkarır ve yerine yenisini takar.

7
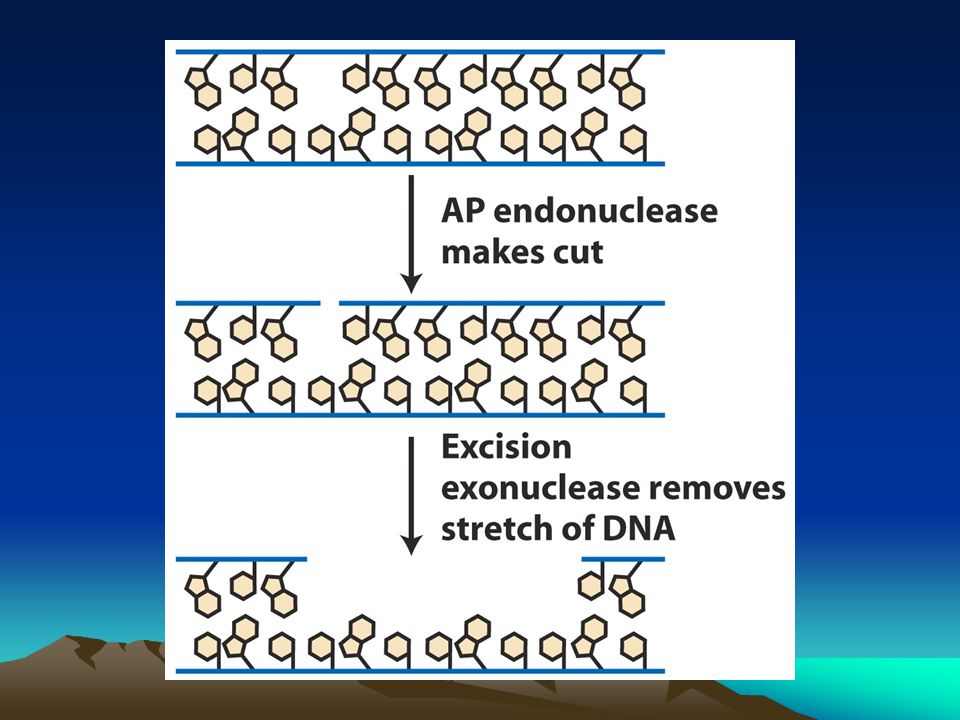
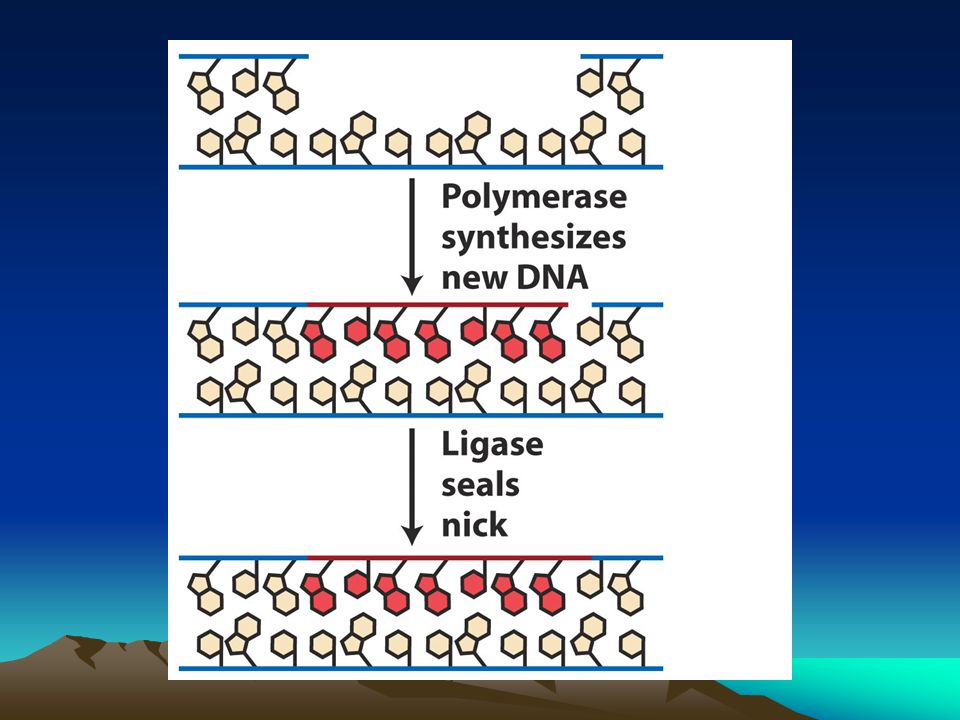
Baz kesme onarımı Bu sistem DNA glikozilazlar tarafından yürütülür.Enzim şeker-baz arasındaki bağı keser ve apurinik yada apirimidinik bölge oluşturur.AP endonükleaz denilen başka bir enzim bazını kaybeden bölgede şeker-P arasındaki bağı keser.deoksiribofosfodiesteraz denilen başka bir enzim bazın kaybedildiği yerin çevresini temizleyerek DNA pol enziminin rahat çalışmasına olanak verir. Bu enzim de boşluğu kalıp zinciri kullanarak doldurur. Çeşitli DNA glikozilaz enzimleri vardır bunlardan urasil DNA glikozilaz DNA yapısında yer alan urasilleri uzaklaştırır.

10
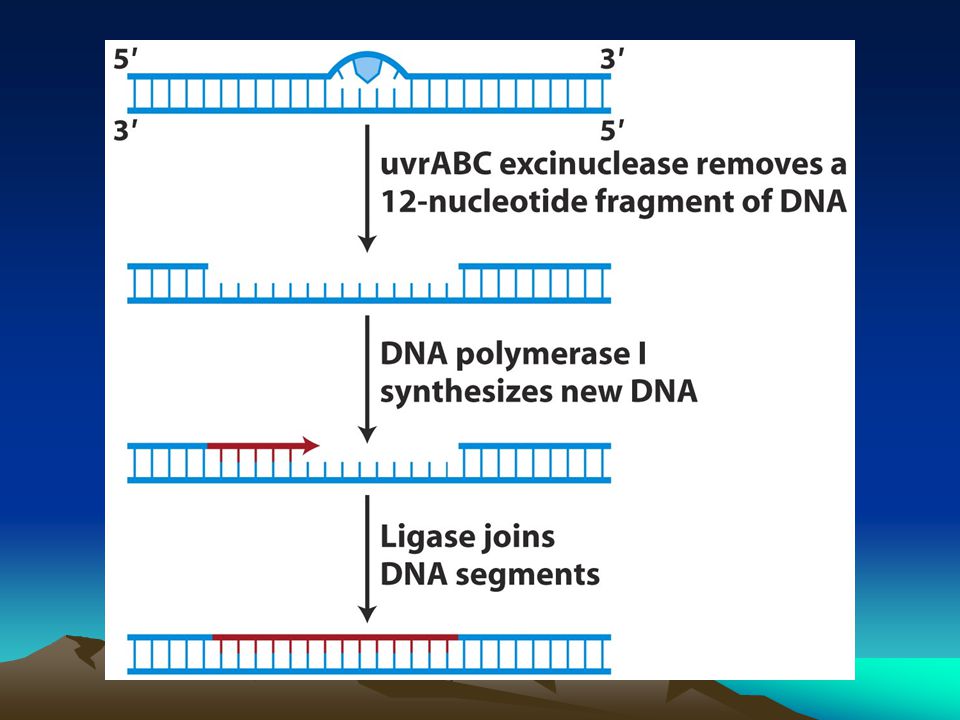
Nt kesme onarımı Eğer DNA heliksinde çift sarmallı yapıyı bozacak bir baz varsa bu sistem tarafından algılanır ve nt buradan uzaklaştırılır.Bu tür değişimler genellikle UV ve aflatoksin gibi ajanlar tarafından yapıya sokulur.Bu tür bir bozukluk E.colide çoklu enzim sistemlerinin devreye girmesine neden olur.uvrABC excinükleaz hasarı tanır ve hasarın sağından ve solundan kesik oluşturur ve 12 nt kısmı bu bölgeden uzaklaştırır:8 bir yandan 4 diğer yandan.Bu açıklık DNA pol I tarafından doldurulur ve kalıp zincir referans olarak kullanılır.DNA ligaz son P bağını kurar.

12
ÖkaryotlardaTranskripsiyonla birleşik onarım
Prokaryotlar gibi ökaryotlarda da baz ve nt kesim onarım sistemleri vardır.Mayadan insana bu sistemler oldukça korunmuş sistemlerdir. Mayalarda nt kesme onarımı kompleks bir enzim grubu tarafından yürütülür buna repairozom denir ve yapısında 20den fazla protein vardır.Bu kompleks hasarı tanır ve hasarın çevresinden 30nt uzaklaştırır ve kalıbı kullanarak boşluğu doldurur.Bu sistem öncelikli olarak transkribe olacak DNA zincirini onarır.Peki bu sistem hangi zincirin onarılacağına nasıl karar verir? Repairozomun 7 polipeptidi bazal transkripsiyon aygıtının elemanıdır ve bunlar iki zincirden hangisinin kalıp hangisinin kalıp olmadığını tanır. Neden tarnskripsiyon ve onarım birlikte çalışır? Aktif olarak bölünen E.colinin aksine çok hücreli organizmalar terminal olarak farklılaşmıştır ve artık bölünmüyorlardır dolaysıyla replikasyon onarımı da mevcut değildir. Bu hücreler ölü hücreler değildir ve genleri aktif olarak mRNAya transribe edilir.Bu hücrelerde hasar gören DNA da onarılmalıdır

15
Postreplikasyon onarımı
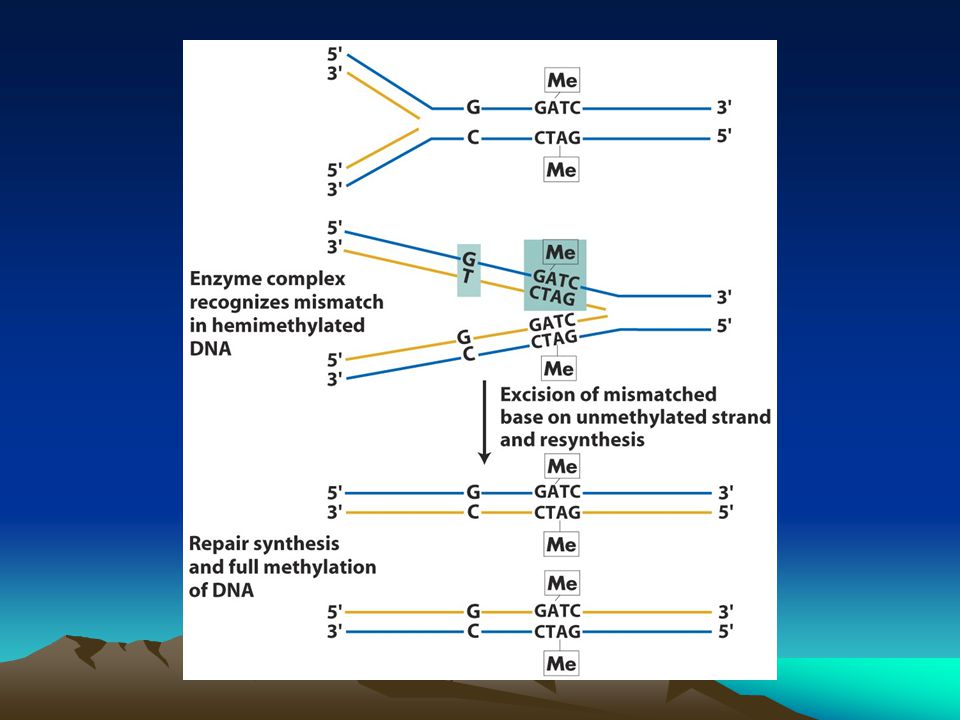
Bazı onarım yolakları replikasyon sırasında meydana gelen hataları 3’-5’ doğrulama yoluyla düzeltirken bazı hatalar bu yolla düzeltilemez.mismatch repair sistemi denen bir sistem bu hasarları tanır ve ortadan kaldırır.Bu sistem 3 işi başarıyla yerine getirmelidir yanlış eşleşen bazları tanımalı Yanlış eşleşen bazlarda hangisinin yanlış eşleştiğini bulmalı Yanlış bazı kesip çıkarmalı ve onarımı yapmalı 2. özellik çok önemlidir çünkü sonraki aşama ve genin işlev kaybı yada kazanımı tamamen bu özelliğe bağlıdır.replikasyon hataları genellikle yanlış bazı yeni sentezlenen zincire takar dolayısıyla onarım sistemi buradan ayırt etme yoluna gidebilir.Bu sistem bakterilerde çok iyi tanımlanmıştır Bilindiği gibi bakteri DNAsı metillenir ve bu metilasyon replikasyondan sonra gerçekleşir bu özellik de onarım sistemine bir avantaj sağlar GATC dizisinde adenin metilaz her zincirde metilli adenin oluşturur (6-metiladenin).Onarım enzim kompleksi hangi zincirin yeni hangisinin eski olduğunu bu metil gruplarından ayırır.metillenmemiş zincir kesilir ve diğer zincire bakılarak doğru baz takılır.
.Onarım enzim kompleksi hangi zincirin yeni hangisinin eski olduğunu bu metil gruplarından ayırır.metillenmemiş zincir kesilir ve diğer zincire bakılarak doğru baz takılır.")
17
İnsanda mismatch onarım sistemi
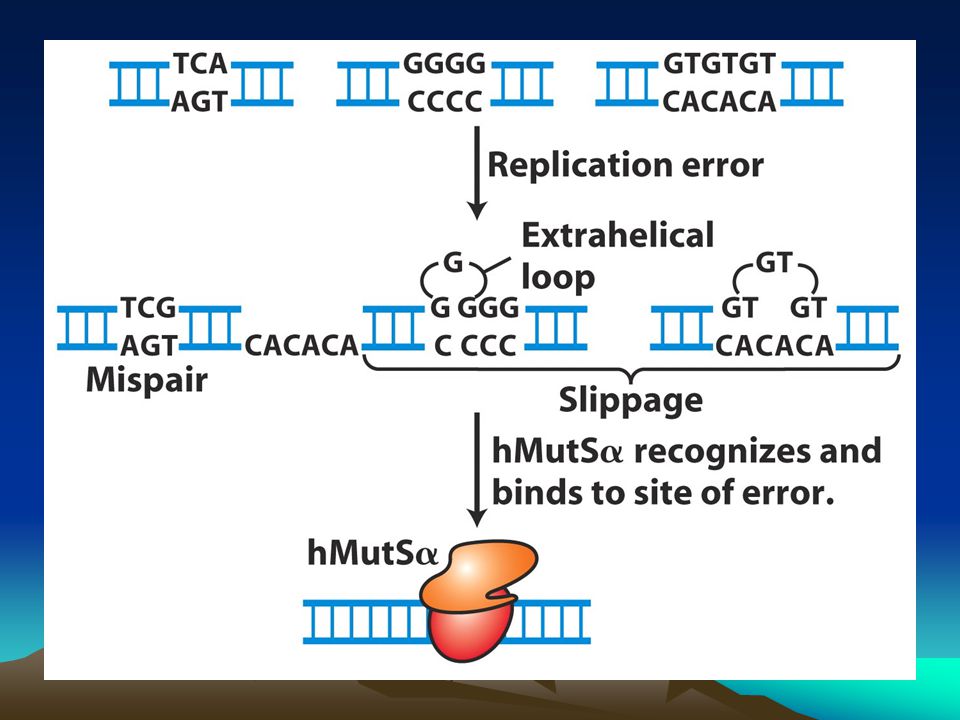
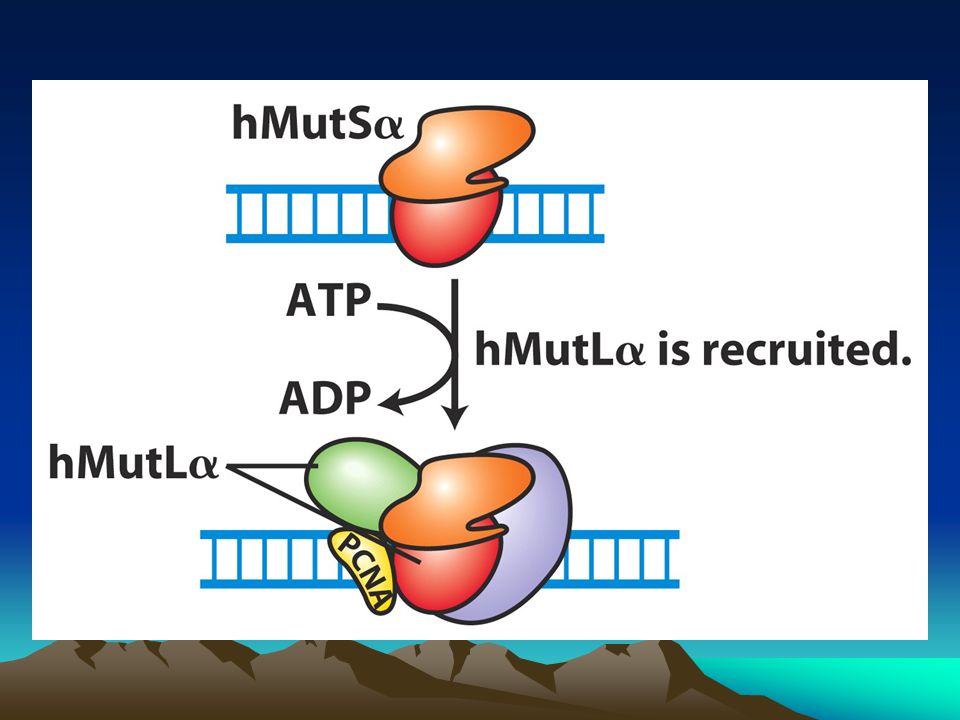
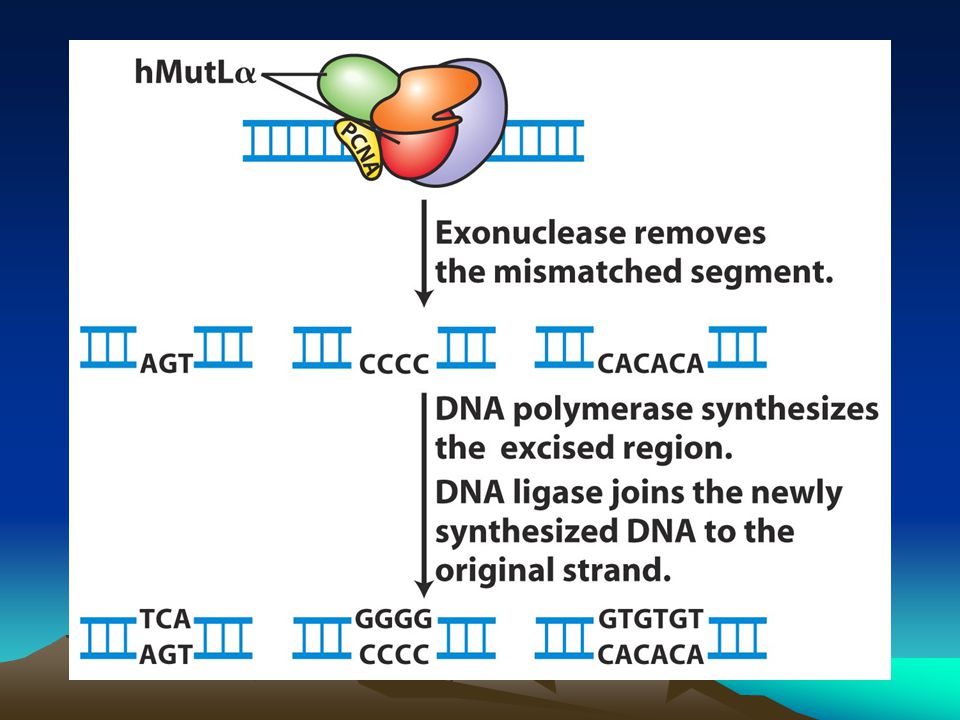
İnsandaki bu sistemin hedeflerinden biri replikasyon sırasında replikasyon kayması nedeniyle oluşan kısa dizilerin eklenmesi yada delesyonuyla sonuçlanan kısa tekrar dizileridir.Bu tür mutasyonlar kansere varan hastalıklara neden olurlar. Bu kısa tekrarlara mikrosatellitler denir ve genom boyunca binlercesi dağılmış durumdadır.Bunların çoğu kodlanmayan bölgelerde bulunmasına karşın birkaç tanesi de ekzonlarda bulunur ve canlının büyüme ve gelişmesini etkiler.

21
Çift zincir kırıklarının onarımı
Çift zincir kırıkları düzeltilmediğinde kromozom aberasyonlarına ve kanser öncesi evreye geçişe neden olabilir.Çift zincir kırıklarının bir özelliği DNA yeniden düzenlenmelerinde rol oynamasıdır.örneğin memeli hücrelerindeki antikor çeşitliliği bu yolla sağlanır.Rekombinasyon da çift zincir kırıklarını kullanarak genetik çeşitlilik sağlar. Çift zincir kırıkları spontan oluşur ve genellikle hücrenin reaktif oksijene yanıtı olarak ortaya çıkar ve iki ayrı mekanizma bu potansiyel ölüm lezyonlarını düzeltir Nonhomolog uç birleşmesi ve homolog rekombinasyon

22
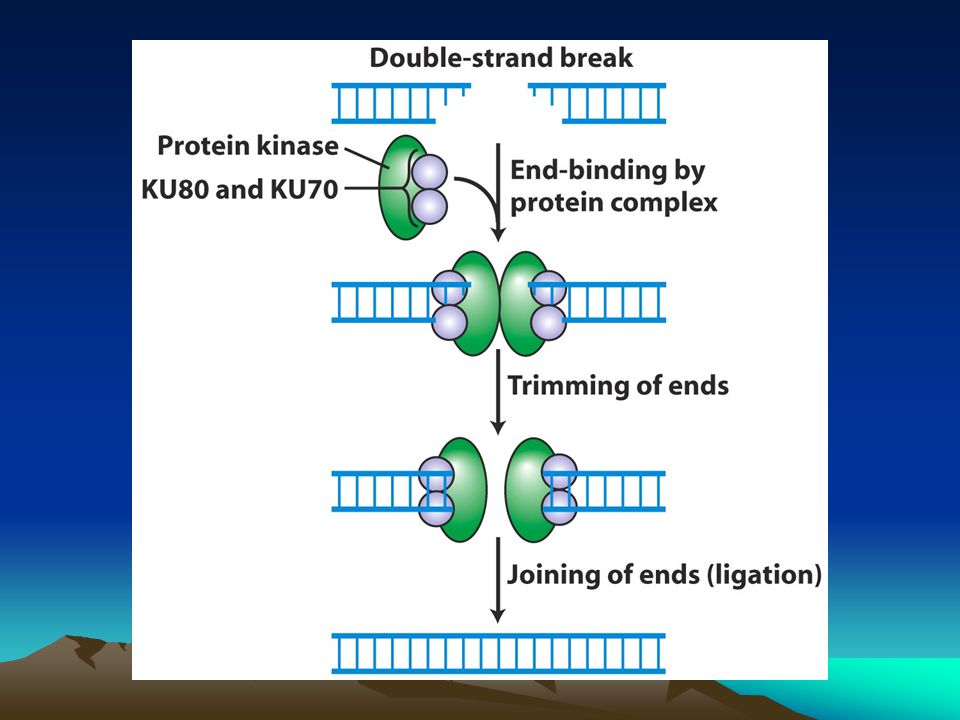
Nonhomolog uç birleşmesi
Eğer bölünmeyen bir hücrede çift zincir kırıkları oluşmuşsa kardeş kromatidler de kalıp olarak kullanılamaz ve bu kırıklar hiç tamir edilmemektense hatalı da olsa hatta bazı baz dizileri kayıp bile olsa tamir edilirler.Bu tür kırıklar eskiden olduğu gibi birbirine yapıştırılırlar.Bu tür onarıma nonhomolog uç birleşmesi denir ve üç aşamada gerçekleşir. 1.Ku70,Ku80 ve DNA-bağımlı protein kinaz kırık uçlara bağlanır ve uçlar kısaltılır daha sonrada bu kısaltılan uçlar yapıştırılır. Ancak bu sırada hücre hatadan kurtulmaz.

24
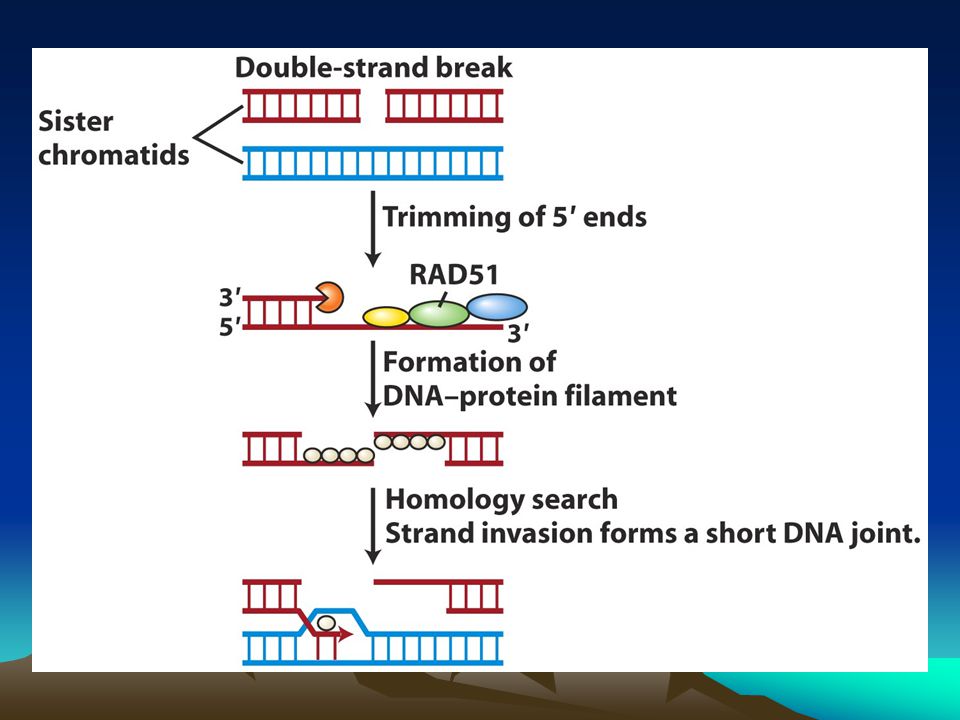
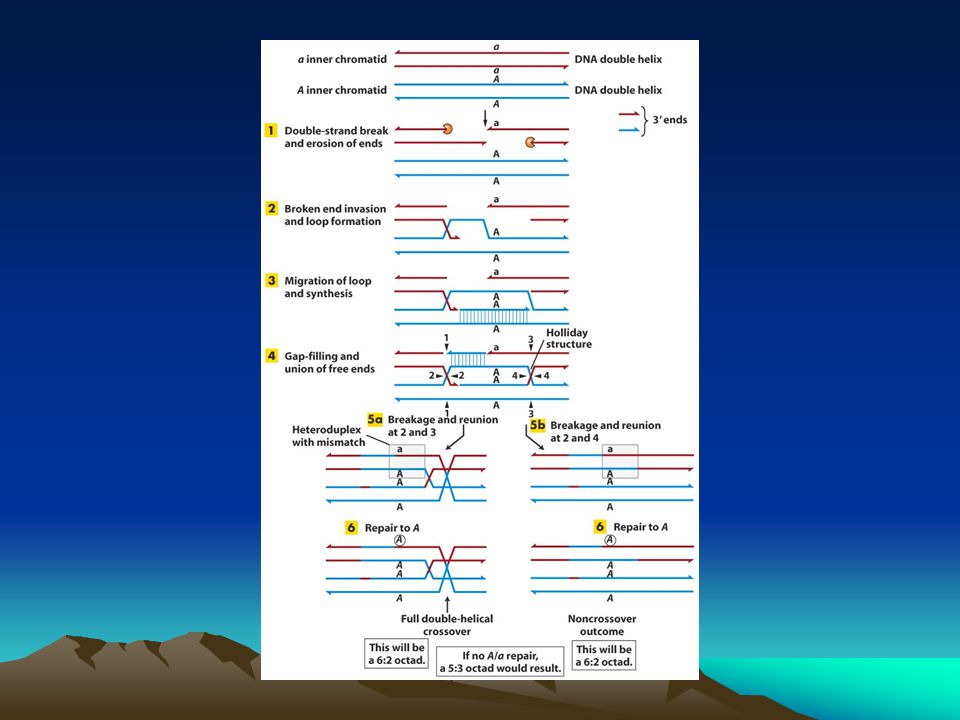
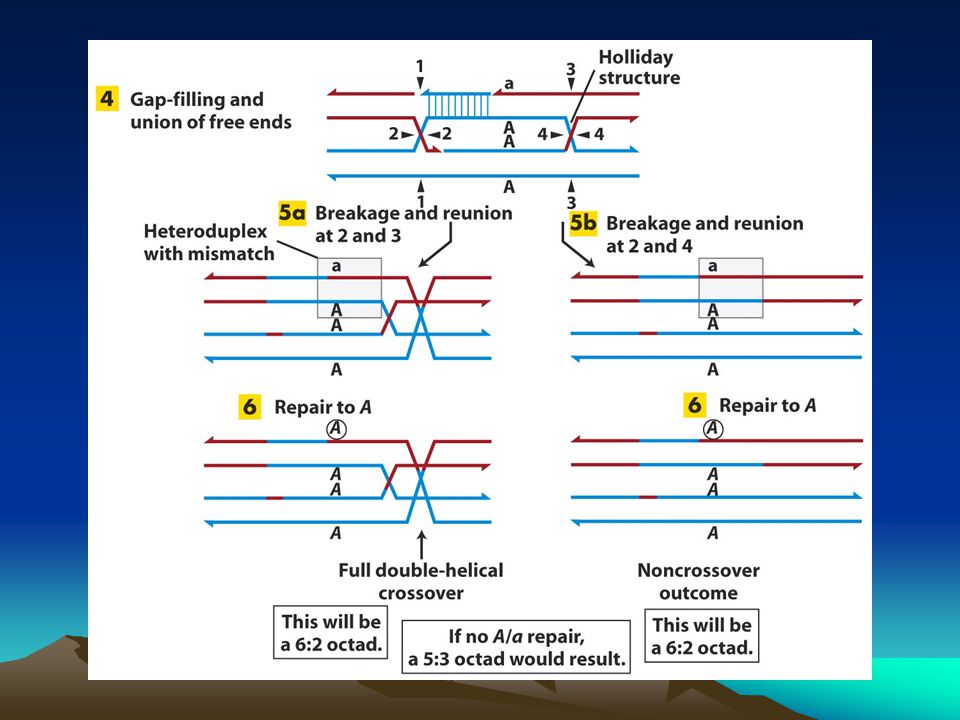
homolog rekombinasyon
Homolog rekombinasyon kardeş kromatidleri kullanarak çift zincir kırıklarını onarır.Bu nedenle onarım hatadan arınmış bir onarımdır. Kırık uçlara özel proteinler bağlanır ve bu uçları kısaltarak tek zincirli bölge yaratırlar sonrada buralara bağlanarak bu kısımları örterler. Bu protein RecA homologu olan Rad51 proteinidir.SOS yanıtı sırasında RecA monomerleri birbiriyle etkileşerek tek zincirli DNA’ya bağlanır ve uzun helikal bir filament oluşturur. Benzer biçimde Rad51 de açığa çıkan tek zincirli DNA’ya bağlanarak uzun bir filament oluşturur.Rad51-DNA filamenti hasar görmemiş kardeş bir kromatid dizisi aramaya başlar ki bu dizi bağlandığı diziye komplenterlik göstermelidir.Bu komplementer dizi onarım reaksiyonunda kalıp olarak kullanılacaktır. Komplementer dizi bulunduktan sonra homolog hasarlı ve hasarsız DNA arasında birleşik bir molekül yapısı oluşur.Hasarlı dizideki kaybolan dizi kardeş kromatiddeki diziden kopyalanır.

Benzer bir sunumlar















>")


 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")

>")




