Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
BİYOKİMYA I 6.DERS PROTEİNLER I

2
Nükleik asitlerde gizli şifre fenotipte PROTEİNLER şeklinde kendini gösterir. Proteinlerin çok çeşitli işlevleri vardır:

3
Küçük moleküllerin taşınması

4
Küçük moleküllerin depolanması
Ferritin, 24 özdeş alt birimden oluşan ve demir depolanmasında iş gören bir protein

5
Hücre ve dokuların yapıtaşları olarak bulunması

6
Kas kasılması

7
İmmün cevabın oluşturulması

8
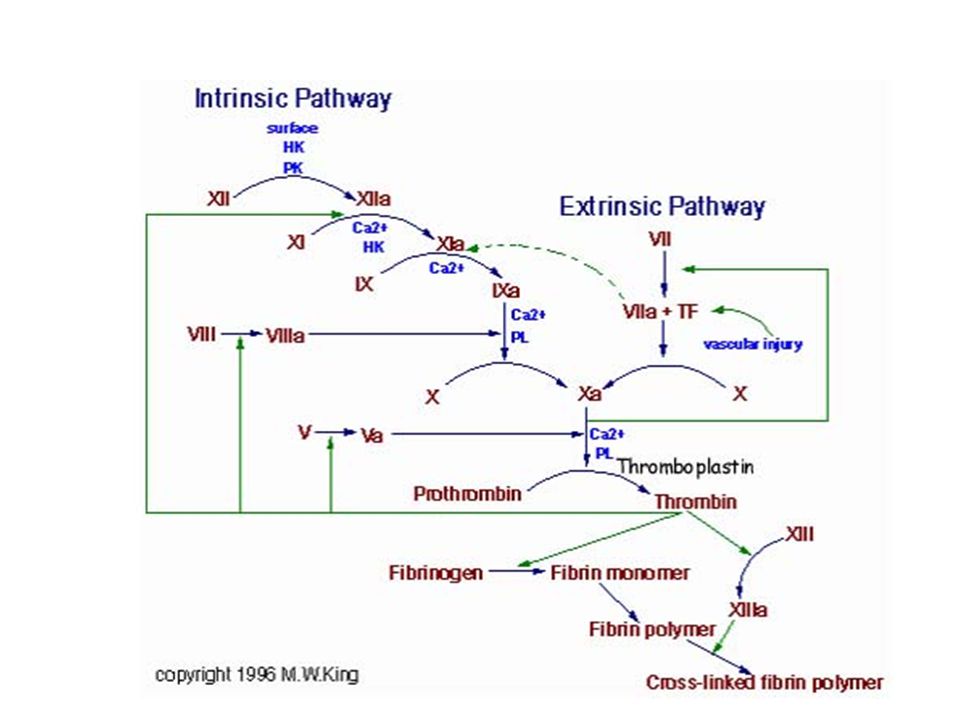
Kanın pıhtılaşması

10
ENZİMLER !!!!

11
Her hücrede binlerce farklı türde protein molekülü bulunur
Her hücrede binlerce farklı türde protein molekülü bulunur. Üç boyutlu yapıları oldukça karmaşıktır:

12
Üç boyutlu yapısı x ışını kırınımı yöntemiyle aydınlatılmış (Kendrew, Geis & ark., 1950ler) ilk protein: Miyoglobin
 ilk protein: Miyoglobin")
13
Her proteinin hücredeki yerleşimi, yapısı ve diğer moleküllerle etkileşimi kendisine özgüdür.

15
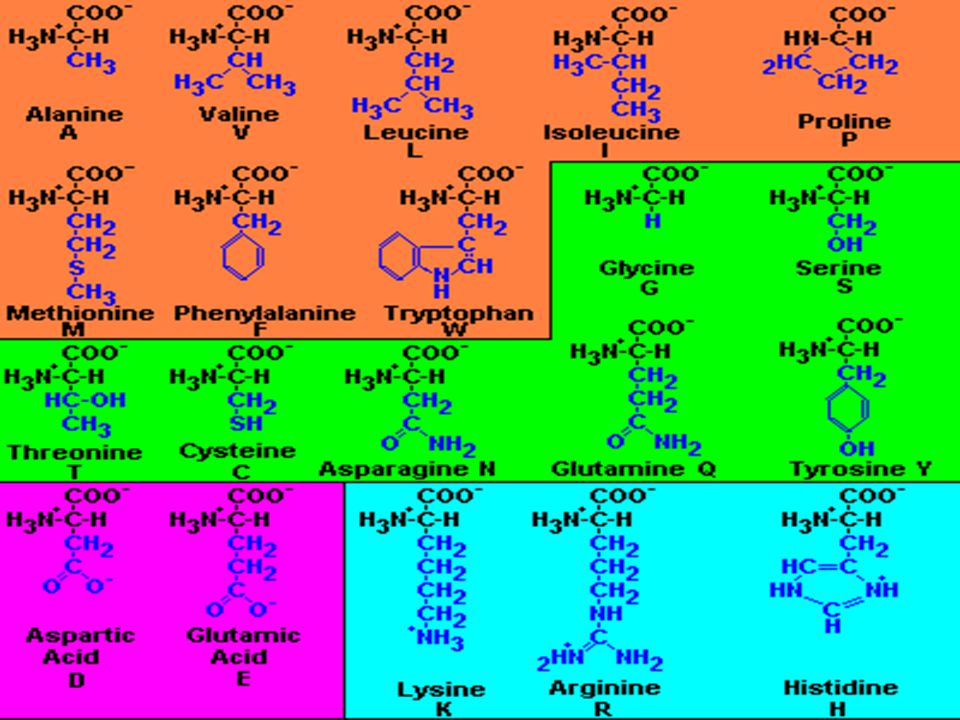
Tüm proteinlerde kimyasal yapıyı oluşturan en küçük birimler -AMİNO ASİTLERdir.

16
Amino grubu, karboksile komşu C atomuna, yani -C atomuna bağlıdır
Amino grubu, karboksile komşu C atomuna, yani -C atomuna bağlıdır. Her amino asidin -C atomunda daima bir hidrojen atomu ve bir yan zincir (R) de bulunur.
 de bulunur.")
17
Örneğin, valin amino asidi:
İyonik olmayan form Bu gösteriliş kimyasal olarak doğru. Ancak in vivo (fizyolojik pH) söz konusu olduğunda COOH grubunun pKa’sı 2, NH2 grubunun pKa’sı 10 Bu nedenle karboksil grubu protonunu kaybeder, amino grubu ise bir proton (H) kazanır, yani molekül ZWİTTERİYON şeklindedir.
 söz konusu. olduğunda. COOH grubunun pKa’sı 2, NH2 grubunun pKa’sı 10. Bu nedenle karboksil grubu protonunu kaybeder, amino grubu ise bir proton (H) kazanır, yani molekül ZWİTTERİYON şeklindedir.")
18
-Karbon atomuna bağlı R yan zincirindeki C atomları Yunan alfabesindeki harflerle simgelenir:

19
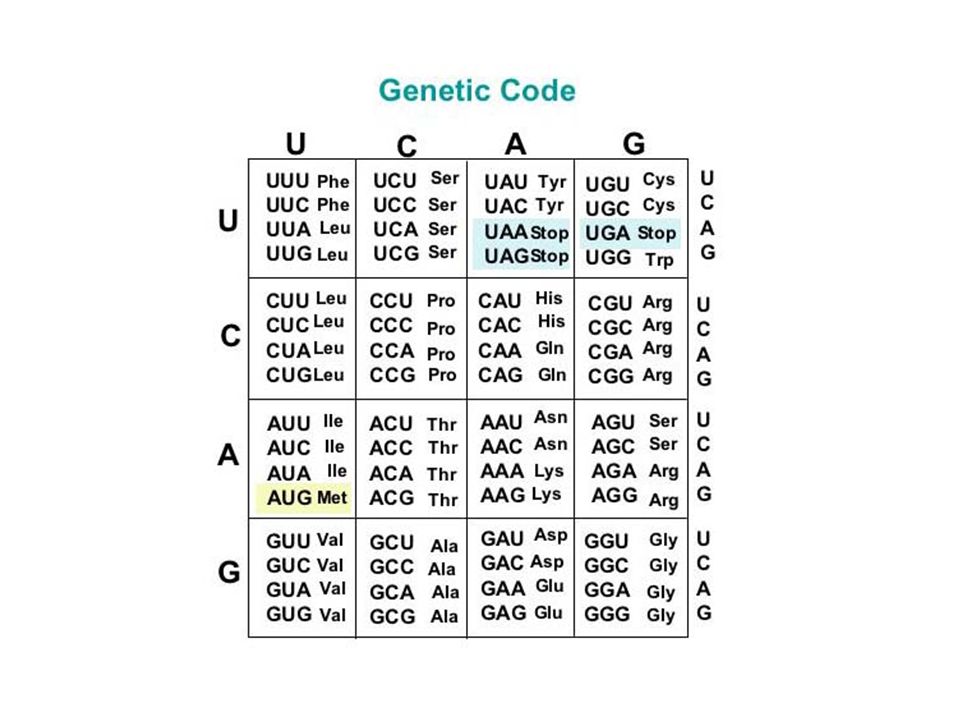
Tüm organizmalarda genler proteinlerin yapısına giren en az 20 çeşit amino asidi şifreler.

21
Ayrıca: 21. ve 22. amino asitler de artık bilinmektedir.

22
Selenosistein az sayıda proteinin yapısına UGA kodonunu kullanarak giren bir amino asittir. Selenosistein amino asidi, glutatyon peroksidaz, thioredoksin redüktaz gibi enzimlerin yapısında yer alır. Selenosistein içeren proteinlere SELENOPROTEİNLER denir.

23
UGA normalde bir bitim (stop) kodonudur, UGA’nın selenosistein için kullanıldığı proteinlerin mRNA’sı farklıdır. mRNA’da SECIS (SElenoCysteine Insertion Sequence) elementi varsa ribozom UGA’yı bitim kodonu olarak algılamaz, bunun yerine amino açil transfer RNA sentetaz tarafından selenosisteinil-tRNA oluşturulur.
 elementi varsa ribozom UGA’yı bitim kodonu olarak algılamaz, bunun yerine amino açil transfer RNA sentetaz tarafından selenosisteinil-tRNA oluşturulur..")
24
SECIS Karakteristik bir nükleotit dizisi ve bu dizideki baz eşleşmeleri nedeniyle ortaya çıkan özel bir sekonder yapı. Bakterilerde, arkelerde ve ökaryotlarda tanımlanmıştır. Bakterilerde hemen yanında bulunan UGA kodonuna etkir. Diğer organizmalarda mRNA’nın 3’ ucuna yakındır ve birden fazla UGA kodonunu yönetir. Hücre ortamında Selenyum YOKSA, selenoprotein sentezi UGA kodonunda biter. Fonksiyonunu yerine getiremeyen bir protein oluşur.

25
Identification of the 22nd genetically-encoded amino acid in a methanogen methyltransferase
Mikrobiyolog Biyokimyacı

26
Metan üreticisi bir bakterinin (Methanosarcina barkeri ) UAG’nin karşılığı bir tRNA ile UAG’nin translasyonuna yardımcı bir enzim (lizil-tRNA sentetaz) içerdiğini buldular. Bu enzim UAG’yi önce lizin amino asidi olarak protein yapısına sokmakta, sonra da lizini enzimatik olarak pirolizine çevirmektedir. PUBLICATION 1. Hao, B., W. Gong, T. Ferguson, C. James, J. Krzycki, and M. Chan. “A New UAG-Encoded Residue in the Structure of a Methanogen Methyltransferase”, Science, 2002, 296, 2. Srinivasan, G., C. James, and J. Krzycki “Pyrrolysine Encoded by UAG in Archaea: Charging of a UAG-Decoding Specialized tRNA”, Science, 2002,

28
Bitim Kodonları: UAA UAG ve UGA
???? Selenosistein Pirolizin

29
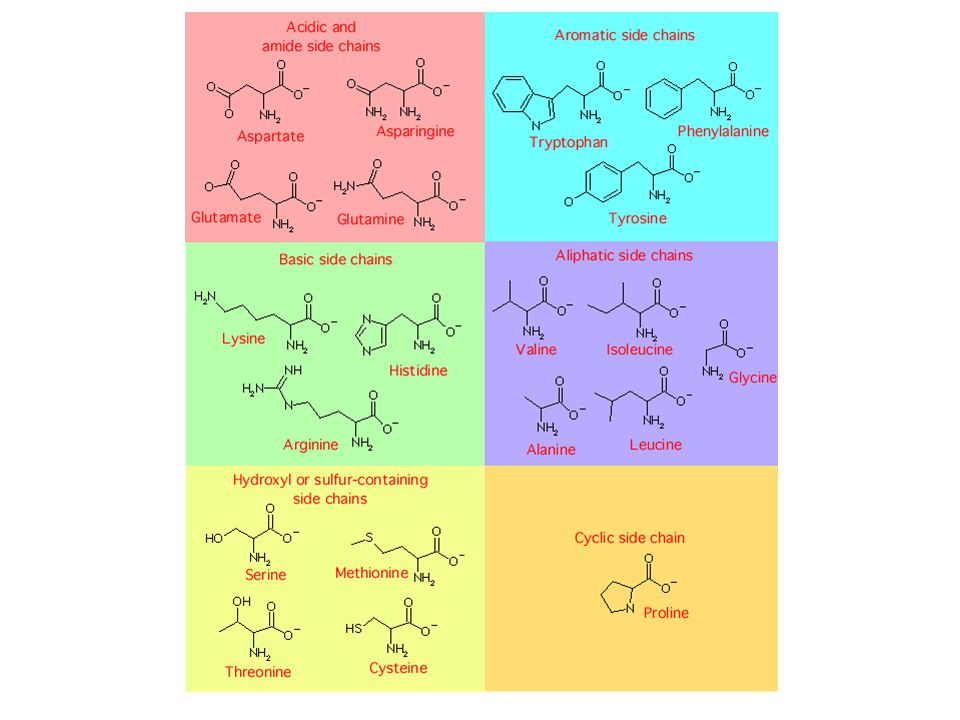
Amino asitleri çeşitli kriterlere göre sınıflandırmak olasıdır, ancak hiçbir sınıflandırma tam olarak ayırt edici değildir!

30
* Farklı amino asitlerde farklı yan zincirler yer alır:

32
FARKLI YAPIDA YAN ZİNCİRLERE SAHİP AMİNO ASİTLERİN KİMYASAL ÖZELLİKLERİ DE FARKLIDIR:
Bazıları hidrofobik veya hidrofilik polar veya apolar iyonlaşabilen veya iyonlaşamayan karakterdedir.

33
ALİFATİK YAN ZİNCİRLERE SAHİP AMİNO ASİTLER:
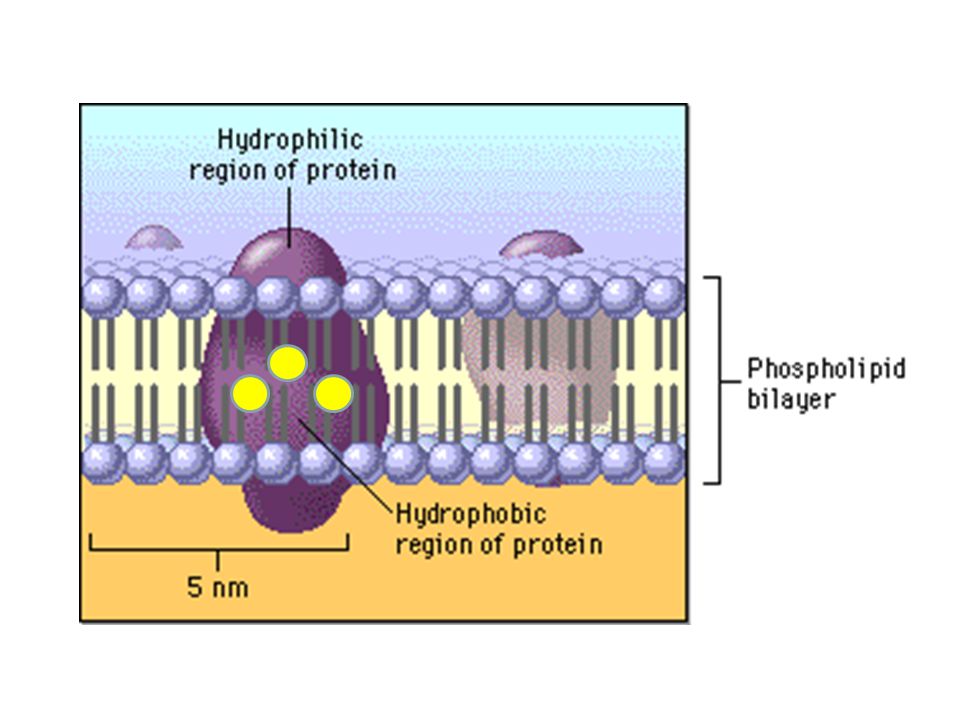
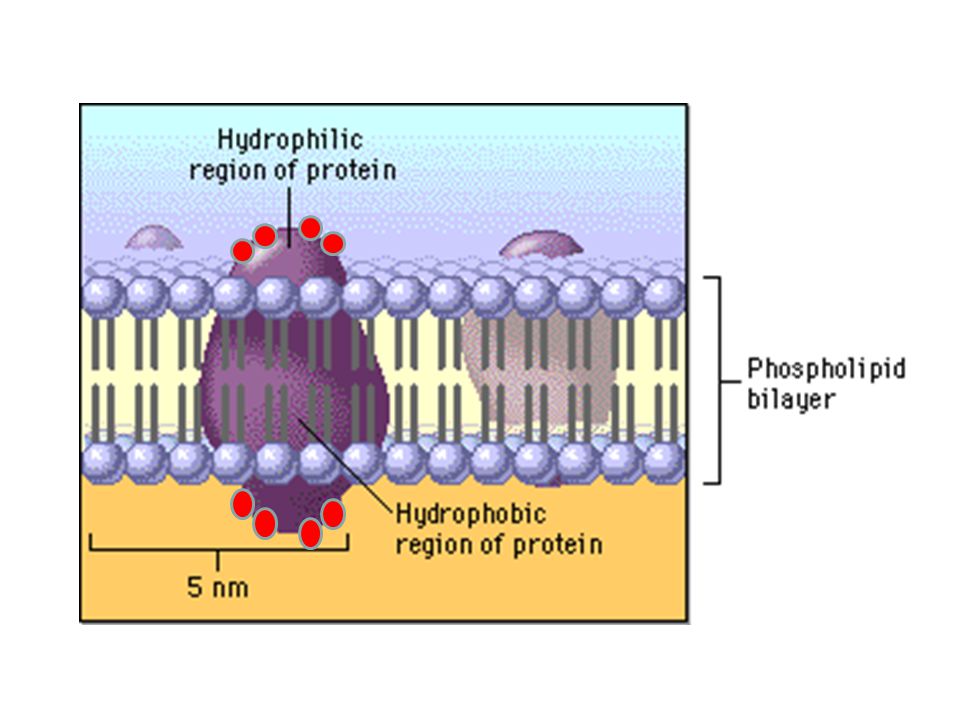
Hidrofobiklik artar İzolösin gibi HİDROFOBİK amino asitler, daha çok proteinin iç kısımlarında bulunur.

35
SİKLİK AMİNO ASİT: Prolin pek çok özellik bakımından alifatik amino asitlere benzer; yan zinciri siklik yapıda olmasına rağmen alifatik karakterdedir. Bununla beraber halkasal yapının esnek olmaması nedeniyle, prolin kalıntılarının proteinin kıvrılıp katlanmış yapısı içinde yer alması zordur.

36
HİDROKSİL VEYA SÜLFİDRİL (TİOL) İÇEREN AMİNO ASİTLER:
Zayıf polar yan zincirlere sahip olmaları nedeniyle (metionin hariç) zayıf hidrofilik karakter taşırlar.
 zayıf hidrofilik karakter taşırlar.")
37
SİSTEİN iki açıdan FARKLIDIR:
1) Nispeten yüksek pH’da İYONİZE OLABİLİR.
 Nispeten yüksek pH’da İYONİZE OLABİLİR.")
38
Bu özellik proteinlerin yapısı açısından önem taşır.
2) İki sistein arasındaki oksidasyon reaksiyonu sonucunda yan zincirler arasında disülfit bağı kurulur ve bir SİSTİN oluşur: Bu özellik proteinlerin yapısı açısından önem taşır.
 İki sistein arasındaki oksidasyon reaksiyonu sonucunda yan zincirler arasında disülfit bağı kurulur ve bir SİSTİN oluşur: Bu özellik proteinlerin yapısı açısından önem taşır.")
39
AROMATİK AMİNO ASİTLER:
Valin, lösin ve izolösin gibi en hidrofobik amino asitlerdendir. Proteinin sudan uzak iç kısımlarında bulunur. Ayrıca alkaloit (bitkilerde bulunan morfin, kolşisin vb. azotlu bileşikler) sentezinde görev alır. Tirozindeki hidroksil grubu bazı proteinler için bir fosforillenme hedefidir. Bu amino asit yüksek pH’da iyonlaşabilir. Triptofan, azot metabolizmasında önemli rolü olan glutamin sentetaz enziminin inhibitörüdür. Polar gruplar da içerdiklerinden zayıf hidrofobik karakter taşırlar.
 sentezinde görev alır. Tirozindeki hidroksil grubu bazı proteinler için bir fosforillenme hedefidir. Bu amino asit yüksek pH’da iyonlaşabilir. Triptofan, azot metabolizmasında önemli rolü olan glutamin sentetaz enziminin inhibitörüdür. Polar gruplar da içerdiklerinden zayıf hidrofobik karakter taşırlar.")
40
Aromatik amino asitlerin (benzen halkasından kaynaklanan)
yakın UV’deki absorpsiyon spektrumları From D. Wetlaufer, Adv. Protein Chem. (1962) 17: © 1962 Academic Press.
 17: © 1962 Academic Press.")
41
Fizyolojik pH’da daima (+) yüklüdürler.
BAZİK AMİNO ASİTLER: Histidin bazik karakteri en zayıf olan amino asittir. Yan zincirdeki imidazol halkası yaklaşık pH 6’da protonunu kaybeder. Histidin proteinin yapısına girdiğinde pKa değeri 7’ye çıkar. Histidin proton alışverişini fizyolojik pH’da yapabildiğinden, proton transferi gerektiren enzimatik reaksiyonlarda önemli rol oynar. Fizyolojik pH’da daima (+) yüklüdürler. Baziklik artar
 yüklüdürler. Baziklik artar.")
42
Bazik amino asitlerin polaritesi yüksektir
Bazik amino asitlerin polaritesi yüksektir. Bu nedenle genellikle proteinlerin dış yüzeylerinde (sulu ortamla ilişkide olacakları yerde) bulunurlar.
 bulunurlar.")
44
ASİDİK AMİNO ASİTLER VE AMİTLERİ:
pH 7’de (-) yük taşıyan amino asitler Bu amino asitlerin pKa değeri o kadar düşüktür ki, protein yapısına katıldıklarında bile, fizyolojik pH’da (-) yük korunur. Bu yüzden bunlara aspartat ve glutamat denir. Yüksüz yan zincirlere sahiptirler, ancak zayıf polar özelliktedirler.
 yük taşıyan amino asitler. Bu amino asitlerin pKa değeri o kadar düşüktür ki, protein yapısına katıldıklarında bile, fizyolojik pH’da (-) yük korunur. Bu yüzden bunlara aspartat ve glutamat denir. Yüksüz yan zincirlere sahiptirler, ancak zayıf polar özelliktedirler.")
45
Bazı amino asitlerin protein yapısına katılmanın ötesinde işlevleri vardır:
Tirozin tiroid hormonlarının yapısını oluşturur. Glutamat Nörotransmitter Triptofan Nörotransmitter serotoninin öncülü Glisin Hem gibi porfirinlerin öncülü Arjinin Nitrik oksidin öncülü

46
Amino Asitlerin Beslenme Açısından Önemi
20 standart amino asitten 8 tanesi insan vücudunda yeterince sentezlenemez ve beslenme ile alınmalıdır. Bunlara ESANSİYEL AMİNO ASİTLERdenir. Sistein, Tirozin, histidin ve arjinin ÇOCUKLARDA Yarı esansiyel olarak kabul edilir, çünkü metabolik yollar tam olarak gelişmemiştir. Essential Nonessential Isoleucine Alanine Leucine Asparagine Lysine Aspartate Methionine Cysteine* Phenylalanine Glutamate Threonine Glutamine* Tryptophan Glycine* Valine Proline* Arginine* Serine* Histidine* Tyrosine* (*) Essential only in certain cases.
 Essential only in certain cases.")
47
PVT TIM HALL ("Private Tim Hall")
Phe Val Thr Trp Ile Met His Arg Lys Leu These ten valuable amino acids have long preserved life in man

48
Electrically Charged (negative and hydrophilic)
List of Amino AcidsAnd Their Abbreviations Nonpolar Amino Acids (hydrophobic) amino acid three letter code single letter code glycine Gly G alanine Ala A valine Val V leucine Leu L isoleucine Ile I methionine Met M phenylalanine Phe F tryptophan Trp W proline Pro P Polar (hydrophilic) serine Ser S threonine Thr T cysteine Cys C tyrosine Tyr Y asparagine Asn N glutamine Gln Q Electrically Charged (negative and hydrophilic) aspartic acid Asp D glutamic acid Glu E Electrically Charged (positive and hydrophilic) lysine Lys K arginine Arg R histidine His H Yellow amino acids contain sulfur. Blue amino acids can be phosphorylated.
 amino acid. three letter code. single letter code. glycine. Gly. G. alanine. Ala. A. valine. Val. V. leucine. Leu. L. isoleucine. Ile. I. methionine. Met. M. phenylalanine. Phe. F. tryptophan. Trp. W. proline. Pro. P. Polar (hydrophilic) serine. Ser. S. threonine. Thr. T. cysteine. Cys. C. tyrosine. Tyr. Y. asparagine. Asn. N. glutamine. Gln. Q. Electrically Charged (negative and hydrophilic) aspartic acid. Asp. D. glutamic acid. Glu. E. Electrically Charged (positive and hydrophilic) lysine. Lys. K. arginine. Arg. R. histidine. His. H. Yellow amino acids contain sulfur. Blue amino acids can be phosphorylated.")
49
Bazı amino asitler proteinlerin yapısına katıldıktan sonra modifiye edilirler. Bu modifikasyonlar sonucunda proteinlerin yapısında aşağıdaki gibi kalıntılar yer alabilir: Hidroksiprolin, 5-Hidroksilizin, 6-N-Metillizin, Fosfotreonin, Fosfoserin, Tirozinin iyotlanmış türevleri [Diiyodotirozin (DIT), Triiyodotirozin (T3), Tetraiyodotironin (Tiroksin,T4)] -Karboksiglutamik asit
, Triiyodotirozin (T3), Tetraiyodotironin (Tiroksin,T4)] -Karboksiglutamik asit.")
50
MODİFİYE AMİNO ASİTLER
Yapısal bir protein olan kollajende prolinin modifiye formu olan 4-hidroksiprolin (hidroksiprolin de denir) oldukça yaygındır. OH grupları protein yapısının dayanıklı olmasında etkilidir. 3 4 2 1
 oldukça yaygındır. OH grupları protein yapısının dayanıklı olmasında etkilidir")
51
Kollajende bazı lizin kalıntıları da hidroksillenmiştir (4-hidroksilizin). Ancak bu modifikasyona daha az rastlanır. Bu kalıntının rolü farklıdır: polisakkaritler için tutunma yerleri sağlar.

52
6-N-Metillizin: Miyozinin yapısında yer alır (kas kasılması)
")
53
O-Fosfoserin proteinlerde bulunan serinin bir protein kinaz (proteinlere fosfat grubu bağlayan enzim) tarafından fosforillenmiş şeklidir. Özellikle bazı enzimlerin yapısında yer alan serin ve treonin kalıntılarına fosfat grubu bağlandığında ya da bu grup ayrıldığında enzim aktivitesi değişir.(Enzim aktivitesinin kontrolü)
.")
54
Fosforillenebilen amino asitler

55
Tiroksin ve triiyodotironin tiroid hormonları olup, tiroglobulin’deki tirozin kalıntılarının modifikasyonu sonucunda oluşurlar.

56
Tiroid hormonlarının biyosentezi

57
-Karboksiglutamik asit, glutamatın K vitamini tarafından uyarılan bir reaksiyonla modifiye edilmesi sonucu oluşur. Bu modifikasyon proteinin kalsiyum bağlamasını sağlar. Bu olay kan pıhtılaşmasında en temel olaylardan biridir.

58
PROTEİNLERİN YAPISINDA YER ALMAYAN BAZI AMİNO ASİTLER
S-adenozilmetionin (AdoMet) metil grubu transferleri için gerekli, metioninin aktive edilmiş bir şeklidir.
 metil grubu transferleri için gerekli, metioninin aktive edilmiş bir şeklidir.")
59
Bir metil grubunun hedef moleküle aktarılması, AdoMet’i S-Adenozilhomosistein’e (AdoHcy) dönüştürür.
 dönüştürür.")
60
Ornitin ve Sitrulin üre döngüsünde yer alırlar
Ornitin ve Sitrulin üre döngüsünde yer alırlar. Bu amino asitler olmazsa NH3 dışarı atılamaz.

61
Azaserin pürin nükleotitlerinin sentezini inhibe eden potansiyel bir antibiyotiktir. Yapısı glutamine benzer, bu nedenle glutamin amidotransferazları (glutaminin amido grubundaki azotu alıcı bir moleküle transfer eden ATP-bağımlı enzimler) geri dönüşümsüz biçimde inhibe eder.
 geri dönüşümsüz biçimde inhibe eder..")
62
Homosistein sistein ve metioninin biyosentezinde ve metioninin yıkımında bir ara üründür. Son yıllarda yapılan çalışmalar dolaşım sistemi ile ilgili hastalıklarda homosistein düzeyinin düştüğünü ortaya koymaktadır. Beslenme katkısı olarak folik asit, kobalamin ve piridoksin alınması, kandaki homosistein düzeyinin düşmesine karşı koruyucu etki ortaya koymaktadır.

63
-Alanin : Koenzim A’nın bir bileşeni olan pantotenik asitin öncülü

64
- Amino butirik asit (GABA): Merkezi sinir sisteminde inhibitör etkisi olan bir nörotransmitter (sinir uyarısı iletici)
: Merkezi sinir sisteminde inhibitör etkisi olan bir nörotransmitter (sinir uyarısı iletici)")
65
Bu tip amino asitlerin diğer örnekleri de aşağıdaki tabloda görülmektedir:

66
Amino asitlerin stereokimyası Düzlemsel formüller amino asit yapısının bazı önemli özellikleri olduğunu göstermektedir.

67
-C atomu TETRAHEDRALdir. Molekülün 3 boyutlu yapısı:
şeklindedir. Düz çizgiler baktığımız yönden görünen, Kesikli çizgiler arkada kalan bağlar Daha basit bir gösterimle:

68
Tetraedron: Üçü aynı köşede birleşen 4 üçgenden oluşan geometrik şekil

70
C atomuna 4 FARKLI grup bağlı ise ASİMETRİK KARBON olarak adlandırılır.
Bu C atomuna KİRAL KARBON da (ya da kiralite merkezi, stereomerkez de) denir.
 denir.")
71
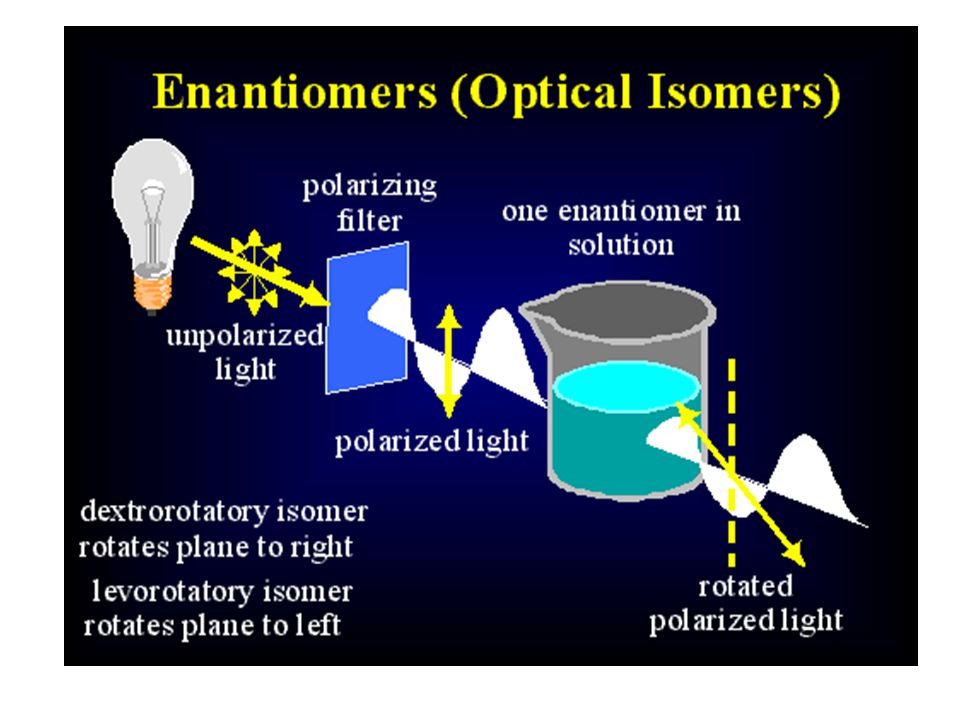
Bir molekül böyle asimetrik bir karbon atomu içeriyorsa, bu molekülün birbirinden farklı iki STEREOİZOMERi (ENANTİOMERi) vardır:
 vardır:")
72
-Amino asitlerin stereoizomerleri

73
Modern organik kimyada enantiomerleri farklandırmak için iki farklı sistem kullanılır: D-L sistemi ve daha yeni olan R-S sistemi. İleride göreceğiz.

74
Enantiomerler deneysel olarak birbirlerinden ayrılabilir
Enantiomerler deneysel olarak birbirlerinden ayrılabilir. Çünkü bunların çözeltileri polarize ışığın düzlemini zıt yönlere çevirir. Bu nedenle enantiomerlere OPTİK İZOMERLER de denir.

76
D- ve L- izomerlerini eşit miktarda içeren karışıma RASEMİK KARIŞIM denir. Böyle bir çözelti optik olarak inaktiftir, çünkü birbirine zıt yönde optik aktivite gösteren formlar birbirlerinin aktivitesini ortadan kaldırır.

77
Glisin hariç tüm amino asitlerin D- ve L-formları vardır
Glisin hariç tüm amino asitlerin D- ve L-formları vardır. Ribozomlardaki protein senteziyle yapıya giren hemen tüm amino asitler L- formundadır.

79
S.aureus’da peptidoglikanın yapısı

80
Grampozitif bir bakteride yer alan peptidoglikan tabakası (D-amino asitler içerir.)
Örneğin, D-Glutamik asit Bakteriyi peptidazların saldırısına karşı koruyabilir.

81
PEPTİTLER VE PEPTİT BAĞLARI
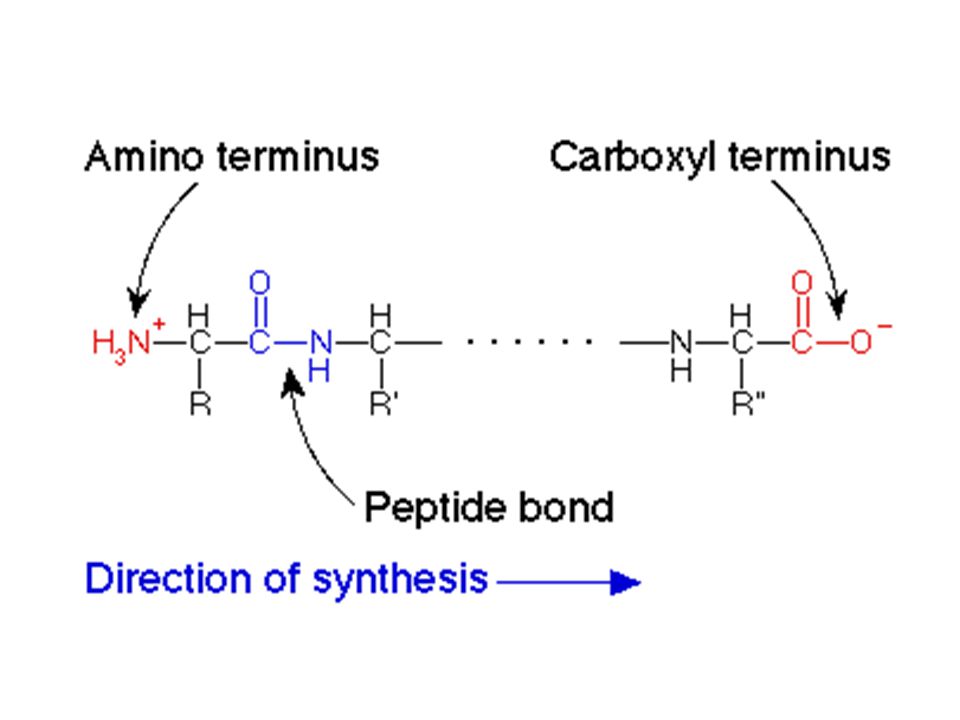
Amino asitler, bir amino asidin -karboksil grubu ile diğer bir amino asidin -amino grubu arasında kurulan bir amit bağı ile, birbirlerine kovalent olarak bağlanabilirler. Bu bağ çoğunlukla peptit bağı olarak adlandırılır. ÜRÜN: PEPTİTLER (burada bir DİPEPTİT)’dir.
’dir.")
82
Glisilalanindeki serbest amino ve karboksil gruplarına glutamik asit ve lizin katılırsa bir TETRAPEPTİT oluşur.

83
Proteinin yapısında yer alan her bir amino aside amino asit kalıntısı amino asit bakiyesi (“residue”) denir. N C H CH3 O

84
Birkaç amino asit kalıntısından oluşan zincirlere OLİGOPEPTİTler denir
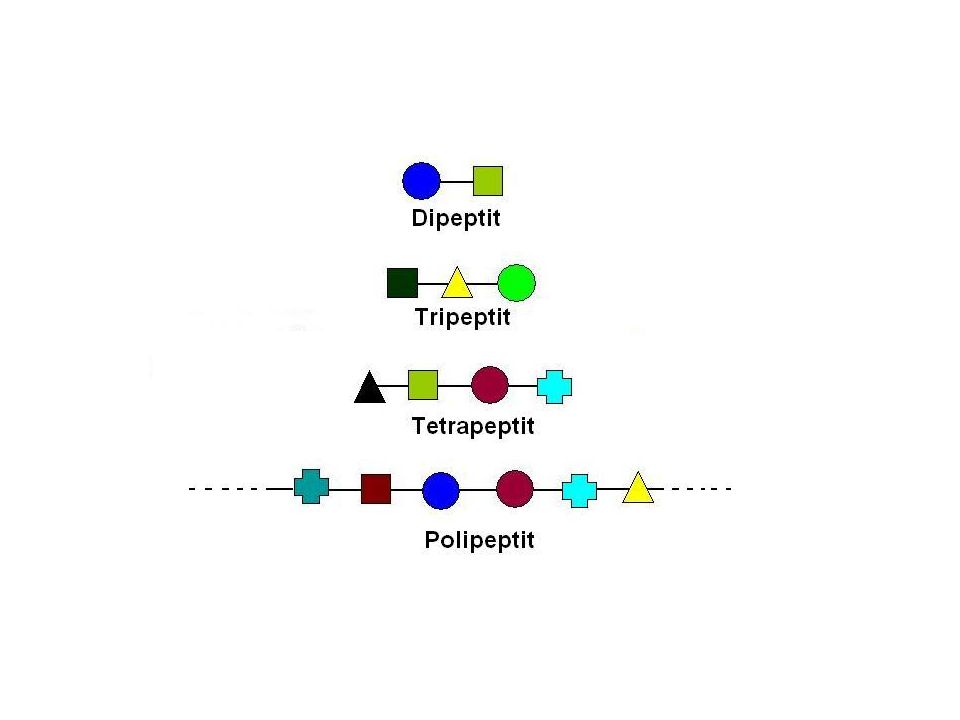
Birkaç amino asit kalıntısından oluşan zincirlere OLİGOPEPTİTler denir. Zincir çok uzunsa POLİPEPTİT olarak adlandırılır.

87
Oligopeptit ve polipeptitlerin büyük bir kısmında bir uçta reaksiyona girmemiş bir amino grubu (amino ucu, N-ucu), diğer uçta ise reaksiyona girmemiş bir karboksil ucu (karboksil ucu, C-ucu) bulunur.
, diğer uçta ise reaksiyona girmemiş bir karboksil ucu (karboksil ucu, C-ucu) bulunur.")
88
Bunun dışında kalan durumlar:
Bazı küçük siklik oligopeptitlerde N-ucu ile C-ucu birbirine bağlanmıştır. N-ucu N-formil veya N-asetil gruplarıyla bloke edilmiş olabilir. C-ucu amit şeklindedir.

89
PEPTİTLERİN KISA YAZILIŞI VE ADLANDIRILMASI:
Üç harfli kısaltma ile (Glu-Gly-Ala-Lys) Tek harfli kısaltma ile (EGAK) Daima amino ucundan başlayarak yazılır ve okunur. Amino asit kalıntıları en sondaki hariç –il eki alır. (Glutamilglisilalanillizin)
 Tek harfli kısaltma ile (EGAK) Daima amino ucundan başlayarak yazılır ve okunur. Amino asit kalıntıları en sondaki hariç –il eki alır. (Glutamilglisilalanillizin)")
90
H CH3 O N C C

91
(Glutamilglisilalanillizin)
Glisilalanindeki serbest amino ve karboksil gruplarına glutamik asit ve lizin katılırsa bir TETRAPEPTİT oluşur. (Glutamilglisilalanillizin)
")
92
Amino asitlerin sırası
GENETİK ŞİFRE’ye göre belirlenir:

93
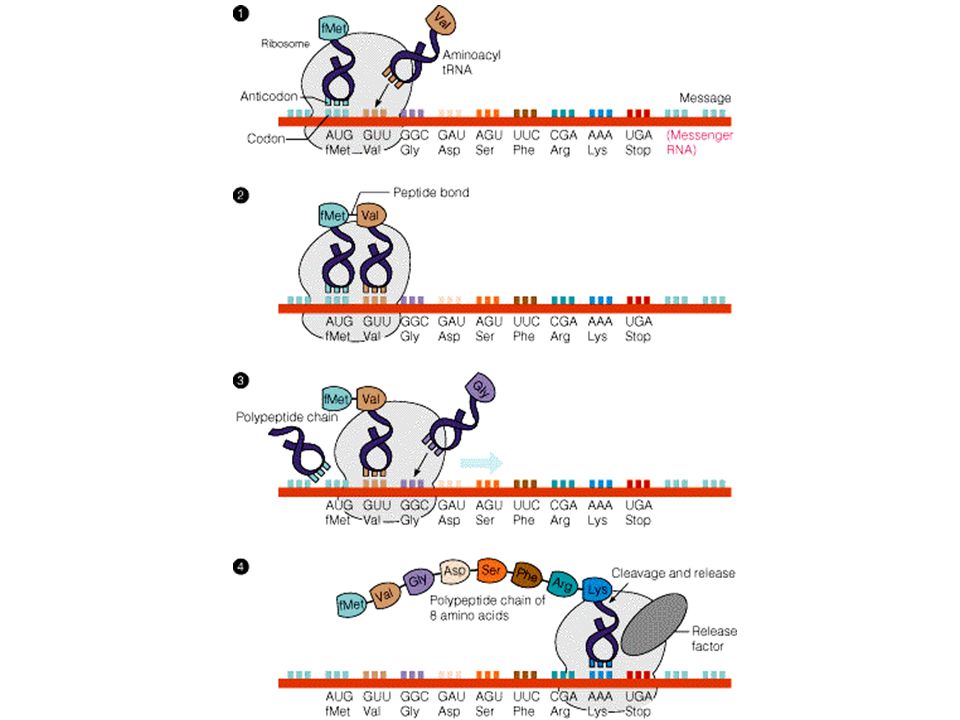
Peptit bağının oluşumu enerji gerektirdiğinden yapıya katılacak her amino asit önce aktive edilir:

95
AUG (başlangıç kodonu) Protein sentezi ökaryotlarda metionin, prokaryotlarda N-formil metionin amino asidi ile başlar. N-formil metionin

96
Bu başlangıç amino asidi ve baştan birkaçı daha translasyon sırasında veya hemen sonra spesifik proteazlar tarafından kesilir.

100
Bazı peptitler sitoplazmada ENZİMATİK olarak sentezlenir: NON-RİBOZOMAL PROTEİN SENTEZİ Örnek: GLUTATYON

101
Glutatyon glutamat, sistein ve glisin içeren bir tripeptittir
Glutatyon glutamat, sistein ve glisin içeren bir tripeptittir. Bu amino asitlerin bağlanma sırası ve şekli aşağıdaki gibidir: Glutamat sisteine -karboksil grubundan bağlıdır. Yani molekül bir -glutamilsisteinilglisindir.

Benzer bir sunumlar




>")





>")




 Nedir? Lipitlerdir.>")







 PC KOPAT>")