Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Prof.Dr.H.Asuman TOKULLUGİL
NÜKLEİK ASİTLER Prof.Dr.H.Asuman TOKULLUGİL

2
1.RNA: Ribonükleik asitler 2.DNA: Deoksiribonükleik asitler
Genetik bilginin depolanmasında → DNA Genetik bilginin transferinde → RNA rol oynar.

3
Azotlu bir baz + şeker nükleotid + (monomer) fosforik asit
nükleotidler=N.A oluşturur (polinükleotid) monomer polimer mononükleotid= N.A yapıtaşı,yapı birimi
 monomer polimer. mononükleotid= N.A yapıtaşı,yapı birimi.")
4
Nükleotid Yapı Taşları
1.Azotlu baz: Pürin veya pirimidin 2.Şeker : D-riboz veya 2 deoksi D- riboz 3.Fosforik asittir

5
I.AZOTLU BAZLAR A-PURİNLER a.PURİNLER

6
B.PİRİMİDİNLER

7
Pseudoüridin : (pirimidin)
İnozin : (pürin) Pseudoüridin : (pirimidin) → t.RNA yapısında metilli türevlerdir.
 Pseudoüridin : (pirimidin) → t.RNA yapısında metilli. türevlerdir.")
8
Molekülün farklı bölgeleri arasında proton
TAUTOMERİZASYON Molekülün farklı bölgeleri arasında proton alış-verişi oksopurin, keto → enol dengeli oksopirimidinlerde C=0 C -OH

9
Keto-enol izomerizasyonu
Fizyolojik şartlarda keto formu

10
Nükleozid= baz + şeker Nükleotid= baz + şeker+ fosforik asit Nükleik asit= poli nükletid

11
b-N GLİKOZİD BAĞI A) PURİNLERDE
 PURİNLERDE")
12
B)PİRİMİDİNLERDE
PİRİMİDİNLERDE")
13
NÜKLEOTİDLERİN FONKSİYONLARI
1.ATP: Evrensel kimyasal enerji taşıyıcısı ATP ADP + Pi → 7.3 kcal/mol enerji ATP AMP + PPi PPi → 2Pi pirofosfataz

14
2.Biosentez reaksiyonlarında
TAŞIYICI ve AKTİVATÖR a)S-adenozil methionin: Transmetilasyon reak.da
S-adenozil methionin: Transmetilasyon reak.da.")
15
b)AMP 3 fosfoadenozin 5’fosfo sülfat =PAPS yapısında bulunur Kondroitin sülfatın sülfatlanması
AMP 3 fosfoadenozin 5’fosfo sülfat =PAPS yapısında bulunur Kondroitin sülfatın sülfatlanması")
16
c)ADP : Koenzim A’nın bir parçası
ADP : Koenzim A’nın bir parçası")
17
d)UTP: Uridin trifosfat
1- Glikojen sentezi UDP-glukoz Galaktoz metab UDP-galaktoz Amino-şeker metab UDP-glukuronik asit Uronik asit yolu 5- Bilirubin metab

18
CDP-Kolin (=sitidin difosfat kolin)
e)CTP:Sitidin trifosfat CDP-Kolin → fosfolipid sentezi CDP-digliserit → trigiserid sentezi CDP-Kolin (=sitidin difosfat kolin)
CTP:Sitidin trifosfat. CDP-Kolin → fosfolipid sentezi. CDP-digliserit → trigiserid sentezi. CDP-Kolin (=sitidin difosfat kolin)")
19
3-Nükleozid trifosfatlar (nükleotid), biyosentez reaksiyonunda gerekli fosfat ve pirofosfatı sağlarlar: Ör:ATP a)Fosforilasyon Reaksiyonu Heksokinaz X +NTP X-P + NDP Glukokinaz Fruktokinaz Glukoz + ATP G.6.P +ADP Proteinkinaz kinaz Mg++ glukokinaz
Fosforilasyon Reaksiyonu. Heksokinaz X +NTP X-P + NDP. Glukokinaz. Fruktokinaz Glukoz + ATP G.6.P +ADP. Proteinkinaz. kinaz. Mg++ glukokinaz.")
20
B) Pirofosforilasyon Reaksiyonu
Ör: Nükleotid sentezinde kullanılan ribozun sentezi Riboz-1-P + ATP PRPP + AMP ( PRPP = 1 Fosforibozil-5-pirofosfat )
")
21
4-Elektron transfer reaksiyonuna katılan koenzimler
NAD (nikotinamid adenin dinükleotid) NADP (nikotinamid adenin dinükleotid fosfat) FMN (Flavin adenin mononükleotid) FAD (Flavin adenin dinükleotid) FAD D-riboz yerine D-ribitol (şeker alkolü) FMN Flavin azotlu baz ama N.A yapısında bulunmaz
 NADP (nikotinamid adenin dinükleotid fosfat) FMN (Flavin adenin mononükleotid) FAD (Flavin adenin dinükleotid) FAD 1-D-riboz yerine D-ribitol (şeker alkolü) FMN 2-Flavin azotlu baz ama N.A yapısında. bulunmaz.")
22
ATP CTP RNA polinükleotidlerinin prekürsörü GTP URASİL RİBOZ UTP dATP dCTP DNA polinükleotidlerinin prekürsörü dGTP TİMİN DEOKSİRİBOZ dTTP

23
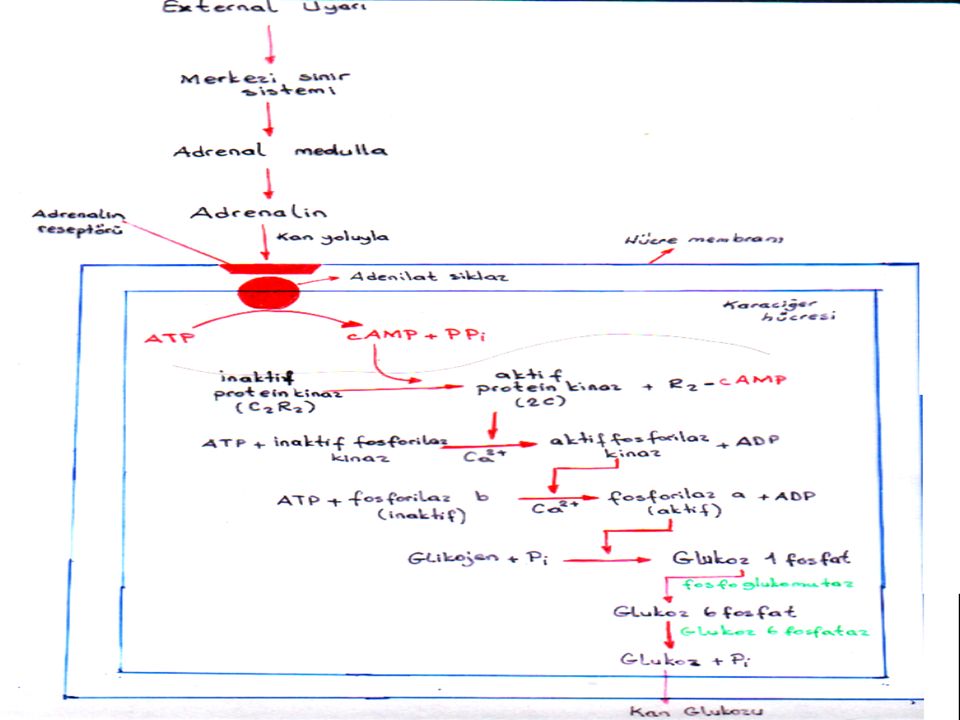
=Secondary messenger= Hücre içi habercisi Ör: c.AMP (çembersel AMP)
5)İkincil haberci =Secondary messenger= Hücre içi habercisi Ör: c.AMP (çembersel AMP) ATP ’5’ cAMP +PPi Adenilat siklaz
İkincil haberci. =Secondary messenger= Hücre içi habercisi. Ör: c.AMP (çembersel AMP) ATP 3’5’ cAMP +PPi. Adenilat. siklaz.")
25
DNA’nın YAPISI ve ÖZELLİKLERİ
1) 3’ 5’ fosfodiester bağı Adenin Urasil (DNA’da timin) Guanin Sitozin
 3’ 5’ fosfodiester bağı. Adenin. Urasil (DNA’da timin) Guanin. Sitozin.")
26
3’ ucunda : serbest OH 2) Polinükleotid zincirinin
5’ ucunda : 5’ trifosfatlar bulunur (öncül molekül) 5’ → 3’
 5’ → 3’")
27
3)RNA ile DNA’nın farkları
Baz :RNA ‘da Urasil DNA’ da Timin Şeker : RNA’ da riboz 2’de OH grubu DNA’da deoksiriboz 2’de H grubu

28
HÜCRELER EUKARYOTİK PROKARYOTİK

29
I-Prokaryotik Hücrelerin Özellikleri
E.koli gibi bakteriler Riketsiya Küçük ilkel Spiroket canlılar 1) Hücreler küçük 2) Sitoplazma: -depo granülleri -nükleer zon (çekirdeğimsi bölge) -sitoplazma zarı (tek zar) 3)Ribozomlar: Protein sentez yeri Ultrasantrifüjde 70 s çökme hızı 30 s , 50 s’lik iki alt birim 4)DNA : - Tek bir dev makromolekül - DNA-protein ilişkisi yok - Küçük sitoplazmik - DNA plazmid,epizom denir
 Hücreler küçük. 2) Sitoplazma: -depo granülleri. -nükleer zon (çekirdeğimsi bölge) -sitoplazma zarı (tek zar) 3)Ribozomlar: Protein sentez yeri. Ultrasantrifüjde 70 s çökme hızı 30 s , 50 s’lik iki alt birim. 4)DNA : - Tek bir dev makromolekül. - DNA-protein ilişkisi yok. - Küçük sitoplazmik - DNA. plazmid,epizom denir.")
30
II-Eukaryotik Hücrelerin Özellikleri:
Yüksek bitki, hayvan, insan hücreleri 1)Hücreler kat büyük 2)Sitoplazma: Zarla çevrili, sınırlı, şekilli organeller -mitokondri, kloroplast -Golgi cisimcikleri -Düz ve kaba EPR -lizozom -çekirdek 3)Ribozomlar: daha büyük 80 s de çöker : 40 s ve 60 s lik 2 alt birim 4)DNA:Kromozomlar halinde dağılmış durumda Drosofilia 8 kromozom İnsan 46 kromozom Tavuk 78 kromozom
Hücreler kat büyük. 2)Sitoplazma: Zarla çevrili, sınırlı, şekilli organeller. -mitokondri, kloroplast. -Golgi cisimcikleri. -Düz ve kaba EPR. -lizozom. -çekirdek. 3)Ribozomlar: daha büyük. 80 s de çöker : 40 s ve 60 s lik 2 alt birim. 4)DNA:Kromozomlar halinde dağılmış durumda. Drosofilia 8 kromozom. İnsan 46 kromozom. Tavuk 78 kromozom.")
31
Kromozom = 1 veya daha fazla sayıda DNA molekülü içerir
Kromatin= DNA + bazik protein(histonlar) = (nükleoprotein) NÜKLEOLUS= Çekirdekçik:Ribozom sentez bölgesi -% 0.1, 0.2 DNA mitokondri veya kloroplastlar içinde yer alır
 = (nükleoprotein) NÜKLEOLUS= Çekirdekçik:Ribozom sentez. bölgesi. -% 0.1, 0.2 DNA mitokondri veya kloroplastlar içinde yer alır.")
32
DNA’nın Kimyasal Analiz Sonuçları
1)Adenin = Timin A=T A/T= 1 Guanin= Sitozin G=C G/C=1 2)Pürin nükleotid sayısı=Pirimidin nükleotid sayısı (A+G=T+C) 3) 4 ve 6. C da (NH2) grubu içeren bazların toplamı = 4 ve 6. C da (=O) grubu içeren bazların toplamı (A+C=G+T) (NH2) (=O) 4)Dissimetri oranı = A+T/G+C Belirli bir tür için sabit ve karakteristik, türler arası değişim gösterir.
Adenin = Timin A=T A/T= 1. Guanin= Sitozin G=C G/C=1. 2)Pürin nükleotid sayısı=Pirimidin nükleotid sayısı. (A+G=T+C) 3) 4 ve 6. C da (NH2) grubu içeren bazların toplamı = 4 ve 6. C da (=O) grubu içeren bazların toplamı. (A+C=G+T) (NH2) (=O) 4)Dissimetri oranı = A+T/G+C. Belirli bir tür için sabit ve karakteristik, türler arası değişim gösterir.")
33
DNA X- Işını Kırınımı Bulguları
1953’te Watson ve Crick DNA çift sarmal yapısı

34
Watson-Crick DNA Modeli
1)Bazlar sarmal eksenine dik düzlem yapar 2)İki baz düzlemi arası arası =0.34 nm 3)Sarmalın bir tam dönüşü= 10 nükleotid=3.4 nm 4) Her 2 zincir birbirine komplementer A=T G=C
Bazlar sarmal eksenine dik. düzlem yapar. 2)İki baz düzlemi arası arası. =0.34 nm. 3)Sarmalın bir tam dönüşü= 10 nükleotid=3.4 nm. 4) Her 2 zincir birbirine. komplementer. A=T G=C.")
35
Hidrojen Bağları

36
5) Fosfodiester iskeleti
2 nükleotid birbirine fosfat grubu aracılığı ile 3’, 5’ grubları vasıtasıyla bağlanır. 5’ 3’ ne doğru uzar.

37
3’, 5’ fosfodiester bağı Fosfoester iskeleti

38
1-Hidrojen bağları 6)DNA Sarmalının Stabilitesi
2- Bazlar arası hidrofobik etkileşimler 7) pH= 7’ de fosfat grubları (-) yüklü. Bu nedenle asidik özellik Bu nedenle N.A denir Hidrofilik Hidrofobik
 pH= 7’ de fosfat grubları (-) yüklü. Bu nedenle asidik özellik. Bu nedenle N.A denir. Hidrofilik. Hidrofobik.")
39
Denaturasyon, Renaturasyon:
İzole edilmiş DNA çözeltisi Oda sıcaklığında pH 7 de viskoz çözelti Aşırı pH viskozite ↓ Isı olunca DNA da fiziksel değişim H bağları Hidrofob etkileşimler bozulur ↓ karşıt zincirler kısmen veya tamamen açılır DNA denaturasyonu= DNA erimesi Tersinir olaydır

40
Hibrit DNA’ların Oluşumu
DNA’lar izole edilir sıçan insan Ayrı ayrı denatüre edilir İnsan sıçan Hibrid DNA 65°C birkaç saat bekletilir Karıştırılır

41
İki tür birbirine ne kadar yakınsa
-Hibridleşme -DNA da sarmal yapı oluşturma oran olur Ör: İnsan –sıçan hibritleşmesi İnsan – maya hibritleşmesi

42
Hibritleşme deneyleri genetik biyokimyada;
1)Akrabalık derecesinin tayini, 2)DNA-RNA hibridleşmesi → DNA-RNA ilişkisi 3)Genlerin izolasyonu için kullanılır
Akrabalık derecesinin tayini, 2)DNA-RNA hibridleşmesi → DNA-RNA ilişkisi. 3)Genlerin izolasyonu için kullanılır.")
43
Prokaryotik Hücrelerin DNA’ları
Prokaryotik hücre → E.coli (bakteri) 200 misli DNA DNA virusu → Lamda faj.(bakteri virusu) E.coli’de DNA -Tek ve çok büyük molekül -Çift sarmal yapısında , halkasal -4 milyon baz çiftinden m.g. -DNA’ nın uzunluğu, hücrenin uzunluğundan 700 kat -Süpercoiling ( DNA fonks. için gerekli) -Nükleer zon (=Çekirdeğimsi bölge) -Topoizomeraz(DNA giraz): Supercoiling yapan ya da açan enzimler -Halkasal DNA :plazmid- sitoplazmada
 200 misli DNA. DNA virusu → Lamda faj.(bakteri virusu) E.coli’de DNA. -Tek ve çok büyük molekül. -Çift sarmal yapısında , halkasal. -4 milyon baz çiftinden m.g. -DNA’ nın uzunluğu, hücrenin uzunluğundan 700 kat -Süpercoiling ( DNA fonks. için gerekli) -Nükleer zon (=Çekirdeğimsi bölge) -Topoizomeraz(DNA giraz): Supercoiling yapan ya da. açan enzimler. -Halkasal DNA :plazmid- sitoplazmada.")
44
Nükleer DNA: Bir kaç bin gen
Küçük halkasal DNA: Bir kaç gen GEN: Tek bir protein veya enzim kodlamak için gerekli DNA parçası (nükleotid dizisi)
")
45
Eukaryotik Hücre DNA’ları
-E.coli’ ye göre : Drosofilia ‘da 25 misli İnsanda 600 misli DNA -E.coli DNA sı=1.4 mm -İnsan hücre DNA sı = 2m

46
KROMOZOM: -nükleoprotein kümeleri
-genetik materyal kromozomlara bölünmüş -kromozom organizmanın türüne özgü sayıda -kromozomların DNA içeriği ve hacmi farklıdır

47
Her KROMOZOMDA 1) 1 DNA sarmalı 2) Proteinler (histonlar) 3) E.coli DNA’ sının katı kadar nükleotid içerir (E.coli DNA’sı= 4 milyon baz çifti) -Eukoryotik hücre DNA’ları doğrusal yapıda
 -Eukoryotik hücre DNA’ları doğrusal yapıda.")
48
GENOM: Bir hücredeki genlerin hepsi
İnsan KC hücresi Hücre çapı = 25 μmetre Çekirdek çapı= 5 μmetre 46 kromozom DNA’ların toplam uzunluğu = 2 metre (=2.106 μ m)
")
49
KROMATİN -Kromozom materyali -Dağınık, koyu boyanan, ağımsı %60 protein %35 DNA dan oluşur %5 RNA

50
NÜKLEOZOMLAR DNA sarmalı Nükleozom çekirdeği H1 histon 10 nm
Ayırıcı DNA NÜKLEOZOMLAR

51
KROMATİNDE: DNA + Histonlar = NÜKLEOZOM Histonlar:
-Bazik proteinlerdir. -Lizin,arjinin -MA: -Prokaryot hüc.de bulunmazlar.

52
NÜKLEOZOM: -10-11 nm çapında
-Her nükleozomda 2’şer tane H2A, H2B, H3 ,H4 proteinleri bulunur (toplam 8 histon prot.) -DNA sarmalı nükleozomun çevresine 2 kez sarılır -Bir nükleozomda 200 baz çifti bulunur nükleotidlik AYIRICI DNA SARMALI -H1 prot. = Ayırıcı bölgede
 -DNA sarmalı nükleozomun çevresine 2 kez sarılır. -Bir nükleozomda 200 baz çifti bulunur nükleotidlik AYIRICI DNA SARMALI. -H1 prot. = Ayırıcı bölgede.")
53
ll ll ll DNA REPLİKASYONU Watson-Crick Hipotezi
- Yeni sentezlenen DNA zincirleri - Ebeveyn DNA’lar kalıp rolü oynar Ebeveyn Yavru sarmallar

54
Messelson ve Stahl Deneyi(1957)
1) E.Coli NH4Cl, (15N)’li besi yerinde üretiliyor Cecium Cl içinde ultrasantrifüj i)Normal E.coli -14N içeren ii)15N ‘li besi yerinde üretilen E.coli Hafif DNA(14N) Ağır DNA(15N)
 E.Coli NH4Cl, (15N)’li besi yerinde üretiliyor. Cecium Cl içinde ultrasantrifüj. i)Normal E.coli -14N. içeren. ii)15N ‘li besi yerinde. üretilen E.coli. Hafif DNA(14N) Ağır DNA(15N)")
55
14N 15N 2)15N içeren E.coli ler 14N LÜ ortama alınıyor 1 nesil sonra
Orta noktaya çöker Hibrit DNA

56
DNA replikasyonunda ; yavru DNA’ nın bir zinciri
3) 2 nesil sonra 14N N 14N 15N Hafif DNA(14N) Hibrit DNA (14N 15N) DNA replikasyonunda ; yavru DNA’ nın bir zinciri ebeveynden diğer zinciri yeni sentezleniyor
 2 nesil sonra. 14N 14N. 14N. 15N. Hafif DNA(14N) Hibrit DNA. (14N 15N) DNA replikasyonunda ; yavru DNA’ nın bir zinciri. ebeveynden diğer zinciri yeni sentezleniyor.")
57
Semikonservatif replikasyon

58
Halkasal DNA’ nın Replikasyonu

59
Halkasal DNA’nın replikasyonu:
- Replikasyon yönünde DNA’nın açılması - Çift yönlü - Origin:Başlangıç noktası nükleotidlik bir bölge, özel bir protein tarafından tanınır

60
E.Coli’de DNA replikasyon çatalı oluştuktan sonra ;
→ 37 C de nükleotid/dakikada ilerler (replike olur) → 1 DNA sarmalı = 10 nükleotidde tam dönüş Bu nedenle ters yönde dönmesi gerekir 4500 devir / dk ters yönde döner ve DNA sarmalı açılır (Bu hız = 70 mil/saat)
 → 1 DNA sarmalı = 10 nükleotidde tam dönüş. Bu nedenle ters yönde dönmesi gerekir devir / dk ters yönde döner ve. DNA sarmalı açılır. (Bu hız = 70 mil/saat)")
61
Eukaryotik DNA’ların Replikasyonu
-Birçok origin mevcut -Çift yönlü -Hızı prokaryotların 10 da biri kadar Tek origin olsa → 2 ayda replikasyon (Binlerce origin → binlerce replikasyon çatalı Bu nedenle → replikasyon hızlı olur )
")
62
Çift yönlü replikasyon sonucu
Kabarcıklar Çift yönlü replikasyon sonucu oluşur REPLİKASYON

63
DNA POLİMERAZ I ENZİMİ

64
Deoksiribonükleozid 5’trifosfat = NTP (dNMP)n + dNTP DNA
(dNMP) n+1 + PPi Uzamış DNA
 n+1 + PPi. Uzamış DNA.")
65
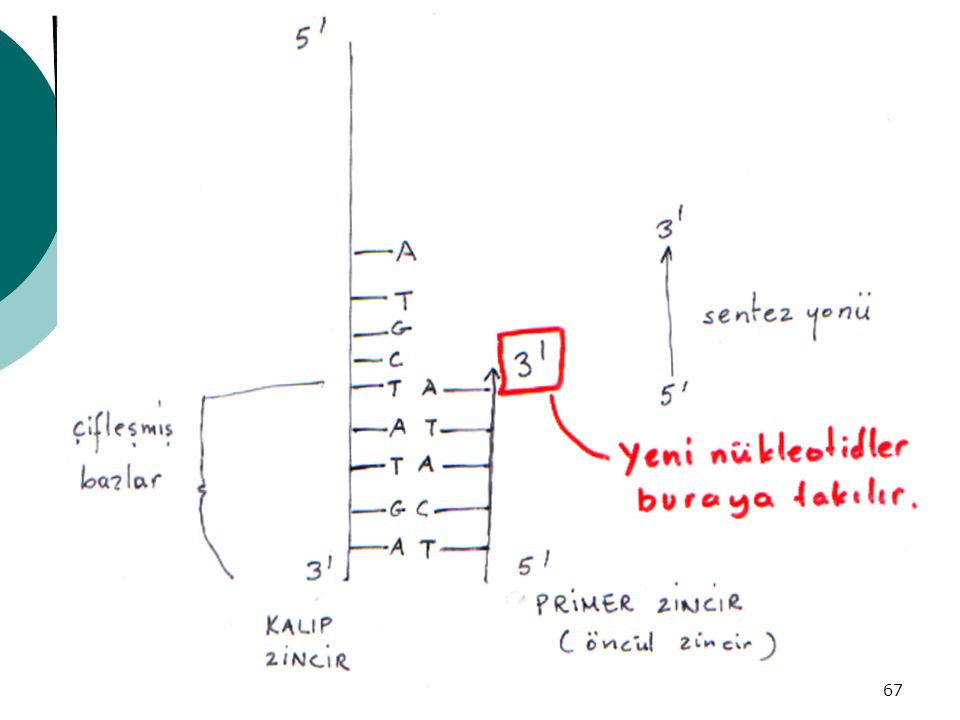
DNA POLİMERAZ I ENZİMİ Substratları : dATP, dTTP, dCTP, dGTP, bir DNA çifti sarmalı Kofaktörleri : Mg++ ve Zn++ iyonları DNA çift sarmalı = başlangıç ve kalıp Mevcut DNA zinciri kalıp kabul edilir Buna KOMPLEMENTER= KARŞIT zincir sentezlenir Sentez (zincirin uzaması) 5’ → 3’ yönünde olur
 5’ → 3’ yönünde olur.")
66
Uzayan DNA zinciri Zincire yeni girecek dNTP dGTP

68
REPLİKASYON OLAYI 1)Başlama noktasının tanınması(origin)
2)Ebeveyn sarmalın dönerek açılması 3)Yavru komplementer zincirlerin oluşumu 4)Zincirin uzaması 5)Zincirin sarmal şeklini alması 6)Replikasyonun sonlanması 20 veya enzim = DNA replikaz sisitemi (REPLİZOM)
Ebeveyn sarmalın dönerek açılması. 3)Yavru komplementer zincirlerin oluşumu. 4)Zincirin uzaması. 5)Zincirin sarmal şeklini alması. 6)Replikasyonun sonlanması. 20 veya enzim = DNA replikaz sisitemi. (REPLİZOM)")
69
E.coli bakterisinde : DNA polimeraz I : Hücre içinde en fazla DNA poimeraz II : İşlevi ? DNA polimeraz III: DNA sarmalının uzamasından esas sorumlu enzim DNA polimeraz III holoenzim (subuniteleri) M.a ‘da -Zn++ iyonları mevcut, aktivite için Mg++ gerekli
 M.a ‘da. -Zn++ iyonları mevcut, aktivite için Mg++ gerekli.")
70
DNA polimeraz I ve III a)Endonükleaz aktivitesi: 5’ → 3’ ucuna doğru yeni nükleotidler takarak ilerler b)Her iki enzimde - DNA kalıp zincirine ve buna sarmal olarak sarılmış PRIMER zincire gerek duyar alt birimi = Ebeveyn DNA ‘daki primer zinciri tanıyarak ona bağlanır c) Ekzonükleaz aktivitesi Hem 3’ → 5’, hemde 5’ → 3’ yönünde zincirden nükleotid koparma aktivitesidir
Her iki enzimde - DNA kalıp zincirine ve buna sarmal olarak sarılmış PRIMER zincire gerek duyar. alt birimi = Ebeveyn DNA ‘daki primer zinciri tanıyarak ona bağlanır. c) Ekzonükleaz aktivitesi. Hem 3’ → 5’, hemde. 5’ → 3’ yönünde. zincirden nükleotid koparma aktivitesidir.")
71
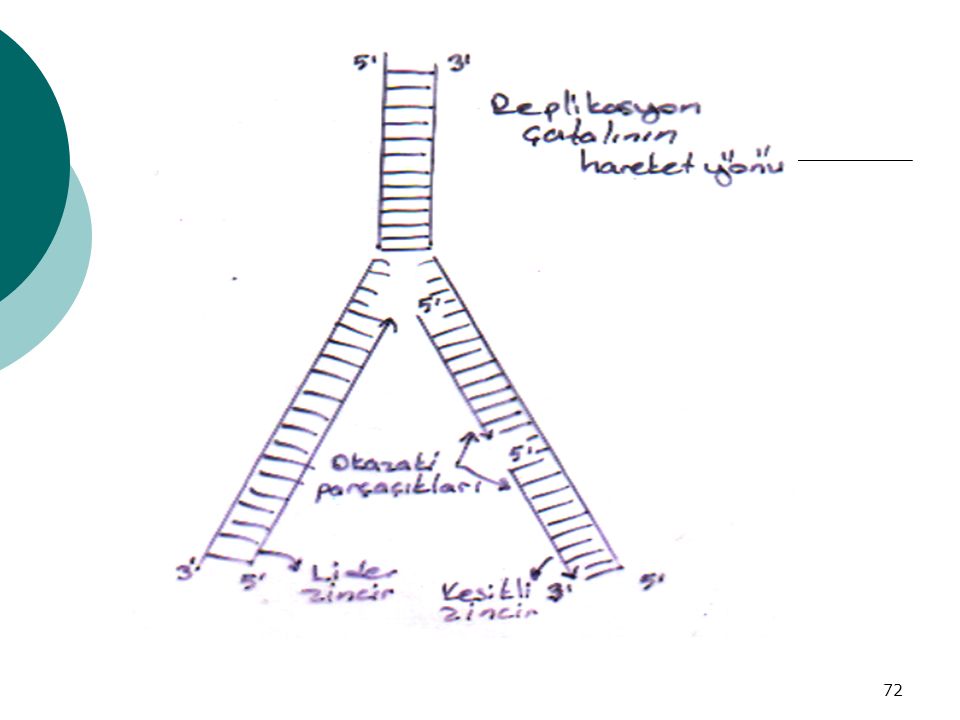
OKAZAKİ PARÇALARI: -Normalde DNA replikasyonu 5’ → 3’ yönünde
Replikasyonda karşıt zincir oluşturulur A zinciri : 5’ → 3’ yönünde normal replikasyon B zinciri : 3’ → 5’ yönünde replike olmaz.Bu nedenle 5’ → 3’ yönünde okazaki parçaları ile replike olur

73
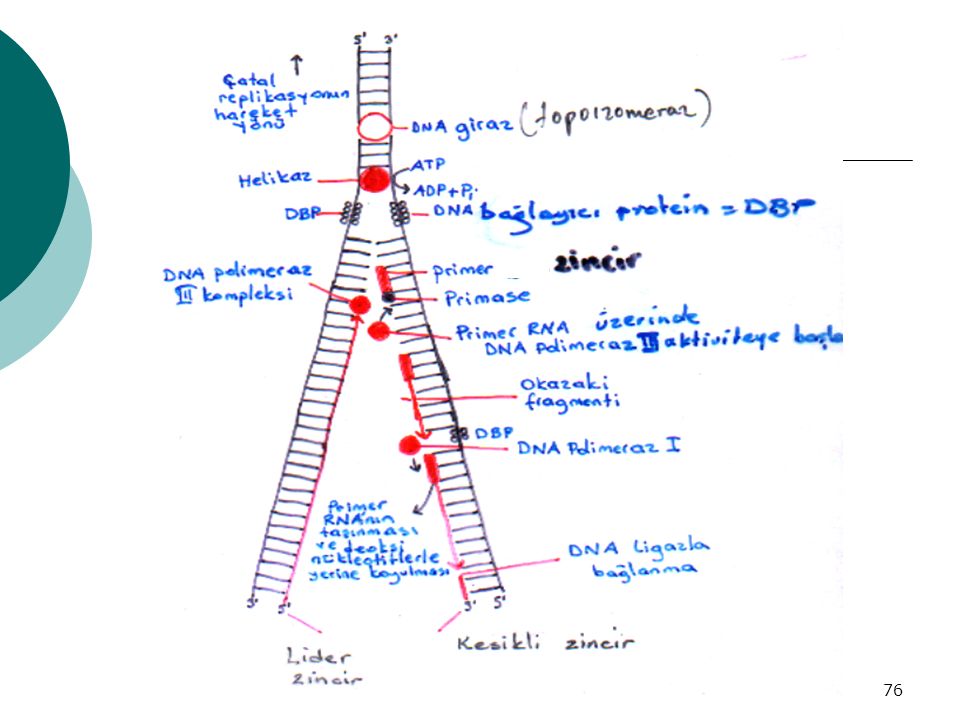
Replikasyondaki enzimler ve işlevleri
1)PRİMAZ Enzimi -Okazaki parçalarınn oluşumu için gerekli -Açılan replikasyon çatalının ucuna birkaç tane ribonükleotid takarak → primer sarmal m.g (öncül) 2)DNA polimeraz III : Primer sarmala → deoksiribonükleotidleri takar, zincir uzar 3)DNA polimeraz I : a) Ekzonükleotidaz aktivitesi ile ribonükleotidleri çıkarır sonra b) Aynı yere : kalıba uyan deoksiribonükleotidleri takar -Okazaki parçaları meydana gelmiş olur
PRİMAZ Enzimi. -Okazaki parçalarınn oluşumu için gerekli. -Açılan replikasyon çatalının ucuna birkaç tane. ribonükleotid takarak → primer sarmal m.g. (öncül) 2)DNA polimeraz III : Primer sarmala → deoksiribonükleotidleri takar, zincir uzar. 3)DNA polimeraz I : a) Ekzonükleotidaz aktivitesi ile. ribonükleotidleri çıkarır sonra. b) Aynı yere : kalıba uyan. deoksiribonükleotidleri takar. -Okazaki parçaları meydana gelmiş olur.")
74
4)DNA ligaz Okazaki parçalarını birleştirir 5)Helikaz enzimi -DNA sarmalını ters yönde döndürerek, zinciri DÜZLEŞTİREN ve AÇAN enzim -Çatalın hemen önünde bulunur 6)DNA bağlayıcı protein(DBP) -Düzleşip açılan DNA’nın tekrar kapanmasını önler 7)Topoizomerazlar -Sarmal 4500 devir/dak hızla düzelir -Dengeleyici oynak nokta olmasa → bu hızda replikasyon çatalının önündeki kromozom ters yönde dönebilirdi
DNA bağlayıcı protein(DBP) -Düzleşip açılan DNA’nın tekrar kapanmasını önler. 7)Topoizomerazlar. -Sarmal 4500 devir/dak hızla düzelir. -Dengeleyici oynak nokta olmasa → bu hızda replikasyon çatalının önündeki kromozom ters yönde dönebilirdi.")
75
Topoizomerazlar a)Dengeleyici oynak nokta: Helikazın önüne oturur.
Helikazla aynı hızda, helikazla kendi arasındaki DNA segmentini 180ºlik dönmelere tabi tutar. DNA düzleşmesine yardımcı olur b)Superkoling olayı: Topoizomeraz sarmalın düzleştirilmesine yardımcı olur Helikaz sarmalı açar (Prokaryotlarda :Topoizomeraz=DNA giraz) Topoizomeraz Helikaz
Superkoling olayı: Topoizomeraz sarmalın düzleştirilmesine yardımcı olur. Helikaz sarmalı açar. (Prokaryotlarda :Topoizomeraz=DNA giraz) Topoizomeraz. Helikaz.")
77
HİSTONLARIN REPLİKASYONU
Kesikli DNA zincirinin bulunduğu yavru sarmalda yeni histonlar yeni nükleozomları oluştururlar. Replikasyon çatalında DNA sentez yönü bir zincirde :5’→3’ , diğer zincirde ise 3’→5’ dir. 3’→5’ yönünde sentez kesikli zincir şeklindedir. 5’→3’ yönünde lider zincir sentezlenir.

78
Eski ve yeni histonlar nasıl dağılım gösterir ?
In vitro DNA sentezi yapılıyor Bir protein sentez inhibitörü olan sikloheksimid ortama ekleniyor (Böylece yeni histon sentezi engelleniyor) Bu şartlar altında DNA sentezi 15 dk devam ediyor Yeni sentezlenen DNA’ nın yarısı DNAaz I’le tamamen yıkılıyor Diğer yarısı ise 200 baz çifti içeren parçalara ayrılıyor.
 Bu şartlar altında DNA sentezi 15 dk devam ediyor. Yeni sentezlenen DNA’ nın yarısı. DNAaz I’le tamamen yıkılıyor. Diğer yarısı ise 200 baz çifti içeren parçalara ayrılıyor.")
79
Bu deney ve density-labeling çalışmaları şunu düşündürmüştür;
Ebeveynden gelen histonlar yeni DNA sarmallarından sadece birisinde bulunur Diğer yavru sarmalda ise önce histon yoktur ve çıplaktır E/M da da replikasyon çatalında bir tarafta histonlar bulunurken diğer tarafta bulunmadığı görülüyor Özellikle replikasyon esnasında ebeveynden gelen histonlar konservatif olarak ayrılı (veya tek bir yavru DNA sarmalında toplanır)
")
80
Özetle; Replikasyon esnasında
ebeveynden gelen histonlar konservatif olarak ayrılır (veya tek bir yavru DNA sarmalında toplanır)
")
81
Bu bulgular; Histonların replikasyon esnasında DNA’ dan ayrışmadığını gösterir Gerçekte, eski histonlar lider zincirin olduğu yavru sarmalda durur Buna karşın yeni sentezlenen histonlar kesikli DNA zincirinin olduğu yavru DNA sarmalında toplanır

82
Bu farkın bir olası nedeni şu olabilir :
Histonlar, tek sarmallı DNA’ya kıyasla, çift sarmallı DNA’ya çok daha kuvvetle bağlanırlar Eski histonlar olasılıkla kesikli sarmalı bırakırlar Çünkü bunda okazaki parçalarının birleşmesinden önce tek sarmallı bölgeler vardır. Bu nedenle öbür zincire geçerler.

83
TRANSKRİPSİYON DNA’ daki baz dizilişi= genetik bilgi içerir
Bazların özel dizilişi= genetik şifre DNA nın ufak bir kısmının açılması (=gen= Bir protein kodlayacak baz dizesi) Transkripsiyon RNA sentezi Translasyon Protein sentezi
 Transkripsiyon. RNA sentezi. Translasyon. Protein sentezi.")
84
TRANSKRİPSİYON → -DNA’ daki baz dizilişine göre genetik
şifrenin RNA’ya aktarılması -Komplementer olay

85
RNA’lar 1)Habercil (messenger RNA= mRNA) DNA Ribozomlara gider
Transkripsiyon 2)Taşıyıcı RNA(transfer RNA= tRNA) Her aa’e özgü bir tRNA vardır 3)Ribozomal RNA (rRNA) Hepsi nükleusta DNA’dan transkripsiyonla sentezlenirler
Taşıyıcı RNA(transfer RNA= tRNA) Her aa’e özgü bir tRNA vardır. 3)Ribozomal RNA (rRNA) Hepsi nükleusta DNA’dan transkripsiyonla. sentezlenirler.")
86
mRNA Uzunluğu değişik, tek zincir halinde molekül
MONOGENİK (MONOSİSTRONİK): Tek genin bilgisini POLİGENİK (POLİSİSTRONİK): Çok genin bilgisini taşıyorsa denir -Prokaryotlardaki mRNA’lar poligenik -Eukaryotlardaki mRNA’lar monogenik mRNA mol. uzunluğu kodladığı polipeptid zincirinin uzunluğu ile sınırlı. 3 baz → 1 aa kodlar Bu nedenle 100 aalik bir polipeptid zinciri için en az 300 bazlık RNA gereklidir
: Tek genin bilgisini. POLİGENİK (POLİSİSTRONİK): Çok genin bilgisini. taşıyorsa denir. -Prokaryotlardaki mRNA’lar poligenik. -Eukaryotlardaki mRNA’lar monogenik. mRNA mol. uzunluğu kodladığı polipeptid zincirinin uzunluğu. ile sınırlı. 3 baz → 1 aa kodlar. Bu nedenle 100 aalik bir polipeptid zinciri için en az. 300 bazlık RNA gereklidir.")
87
5’ 3’ Protein kodlamaz Protein kodlar
Prokaryot mRNA ları gen.la kodladıkları polipeptid zinciri için gerekenden daha uzun →5’ ucunda :LİDER bölge ( baz) polipeptid kodlamaz Poligenik mRNA’larda, INTERGENİK BÖLGE polipeptid kodlamaz Protein kodlamaz Protein kodlar -Bir metabolik yol enzimleri için poligenik mRNA kullanılır İntergenik bölge Gen II Lider bölge Gen I 5’ 3’
 polipeptid. kodlamaz. Poligenik mRNA’larda, INTERGENİK BÖLGE polipeptid kodlamaz. Protein kodlamaz Protein kodlar. -Bir metabolik yol enzimleri için poligenik mRNA kullanılır. İntergenik. bölge. Gen II. Lider. bölge. Gen I. 5’ 3’")
88
mRNA Sentezi: DNA’ya bağımlı RNA polimeraz enzimi
-DNA ‘nın bir zincirini kalıp gibi kullanır -Buna komplementer RNA zinciri oluşturur -Aktif merkezinde Zn++ -Mg++ ‘a gerek duyar -Substratları: ATP, GTP, UTP, CTP -5’ → 3’ yönünde zincir uzaması -3’ ucuna ribonükleotid birimlerini takar

89
RNA polimeraz reaksiyonu:
n(NMP)n + NTP (NMP) n+1 + PPi Ribonükleosid uzamış RNA ( ve 5’ trifosfat fosfat grubları) ( fosfat grupları)
n + NTP (NMP) n+1 + PPi. Ribonükleosid uzamış RNA ( ve 5’ trifosfat fosfat grubları) ( fosfat grupları)")
90
DNA kalıp görevi görür DNA’da : A T G C (deoksiribonükleotid) RNA’da : U A C G (ribonükleotidler) - Primer zincire gerek yok, ama ÖZGÜL BİR BAŞLAMA noktası gerekli -RNA polimeraz bu noktaya oturduktan sonra→ TRANSKRİPSİYON başlar

91
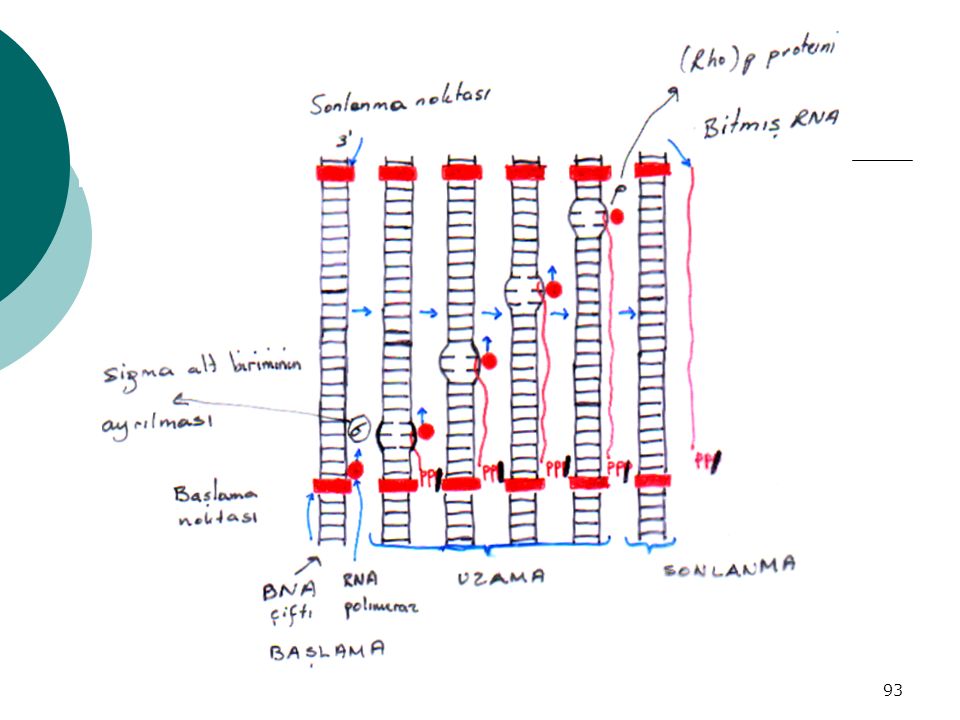
TRANSKRİPSİYON’un Safhaları
RNA polimerazın→ 1- Başlama noktasına oturması 2- Birkaç fosfodiester bağı yapması 3- Sigma birimi (enzimin bir alt birimi, holoenzimden ayrılması 4- Zincirin uzaması 5- Genin transkripsiyonunun bitme sinyali= DNA kalıbı üzerindeki DURMA DİZİSİ 6- RNA ve RNA polimerazın DNA’dan ayrılması. (Rho) P proteini
 P proteini.")
92
RNA polimeraz : -Kompleks, holoenzim
Prokaryotlarda: -M:A -Kompleks, holoenzim -5 polipeptid subuniti var (2, , ’, ) -mRNA, tRNA; rRNA sentezler Eukaryotik hücrelerde: RNA polimeraz I : Nukleolusta lokalize rRNA sentezler RNA polimeraz II : Kromatin içinde lokalize mRNA sentezler RNA polimeraz III :Kromatin içinde lokalize tRNA ve 5s’lik rRNA sentezler
 -mRNA, tRNA; rRNA sentezler. Eukaryotik hücrelerde: RNA polimeraz I : Nukleolusta lokalize rRNA sentezler. RNA polimeraz II : Kromatin içinde lokalize mRNA. sentezler. RNA polimeraz III :Kromatin içinde lokalize tRNA ve. 5s’lik rRNA sentezler.")
94
Transkripsiyonun İnhibisyonu
Aktinomisin D: Prokaryot ve eukaryotlarda ; DNA sarmalında G-C arasına oturur, transkripsiyonu kitler ve zincir uzayamaz Akridin D: Aktinomisin D gibi aktivite Rifampicin D: Prokaryotlarda RNA polimerazın bir alt birimine bağlanarak enzimi bloke eder

95
Posttranskripsiyonel İşlem
Enzimatik olarak RNA’ya bazı grubların takılması ve çıkarılması → RNA’nın AKTİFLEŞMESİ DNA RNA zinciri AKTİF RNA transkrip Posttranskripsiyonel işlem

96
Heterojen nükleer RNA =hnRNA
Eukaryotik hücrelerde önce nükleer RNA’lar sentezlenir Heterojen nükleer RNA mRNA + sRNA(small RNA)
")
97
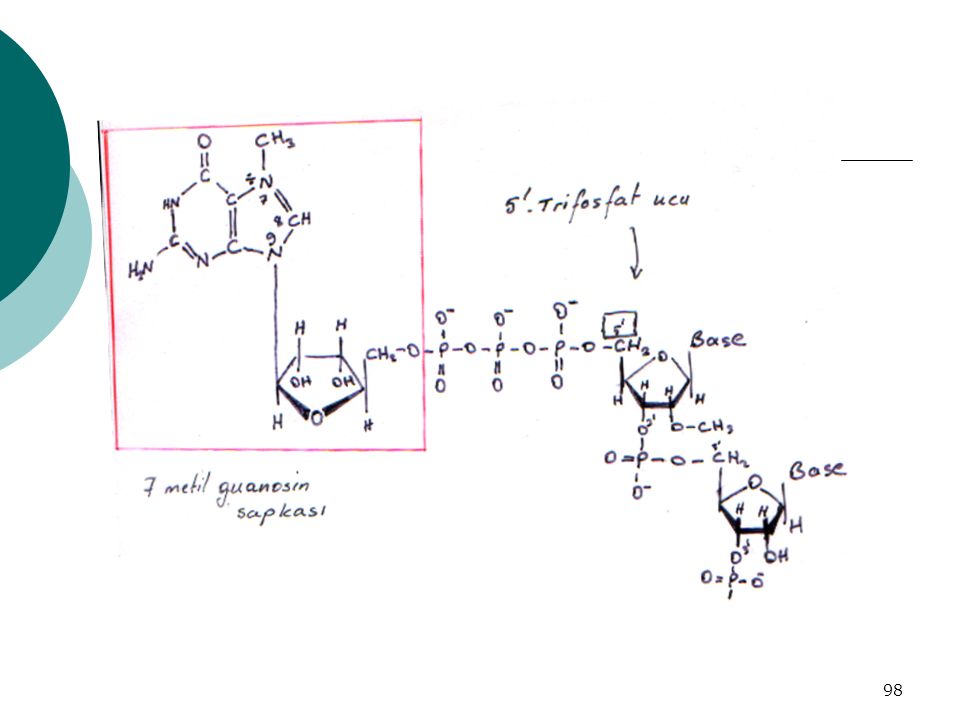
Eukaryotik mRNA’ların, Prokaryotik mRNA’lardan Farkları:
1) Eukaryotik mRNA’lar :MONOGENİK 2) Eukaryotik mRNA 3’ ucunda POLİ-A KUYRUĞU ( tane –A-A-A-) (Poliadenilat polimeraz, substrat ATP) 3) Eukaryotik mRNA 5’ ucunda → 7-METİL GUANOSİN şapkası Şapka → Translasyonu başlatmak üzere ribozoma bağlanmada yardımcı Şapka ve kuyruk → mRNA’ yı enzimatik yıkımdan korur
 Eukaryotik mRNA’lar :MONOGENİK. 2) Eukaryotik mRNA 3’ ucunda POLİ-A KUYRUĞU. ( tane –A-A-A-) (Poliadenilat polimeraz, substrat ATP) 3) Eukaryotik mRNA 5’ ucunda → 7-METİL GUANOSİN. şapkası. Şapka → Translasyonu başlatmak üzere ribozoma bağlanmada yardımcı. Şapka ve kuyruk → mRNA’ yı enzimatik yıkımdan korur.")
99
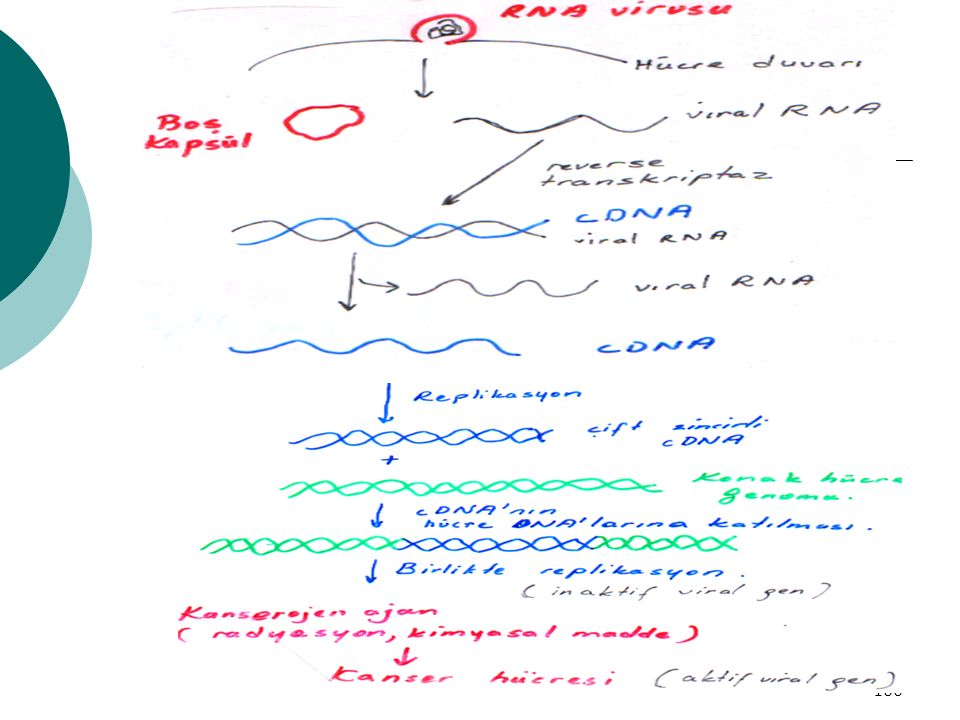
Reverse Transkriptaz (Ters transkripsiyon yapıcı) ve Kanser Oluşumu
Viral RNA Komplementer Reverse DNA (cDNA) transkriptaz Kuşlarda Rous sarkomu etkeni :RNA virusu Bunda ; RNA’ ya bağımlı DNA polimeraz (reverse transkriptaz)
 transkriptaz. Kuşlarda Rous sarkomu etkeni :RNA virusu. Bunda ; RNA’ ya bağımlı DNA polimeraz. (reverse transkriptaz)")
101
Hormon Üretimi Reverse transkriptaz sentetik gen sentezi
E.coli’ye verilir Plazmid oluşturma (cDNA) Hızla çoğalır Translasyon Sonuçta istenen protein,Ör:İNSULİN
 Hızla çoğalır Translasyon. Sonuçta istenen protein,Ör:İNSULİN.")
102
TRANSLASYON Replikasyon DNA Transkripsiyon Reverse Transkripsiyon
(R.Transkriptaz) RNA Translasyon PROTEİN
 RNA. Translasyon. PROTEİN.")
103
PROTEİN SENTEZİ (Translasyon)
a) Protein sentezi çok karmaşık bir olaydır Eukaryotik hücrelerde : 70 tane ribozomal protein 20 tane protein: aa aktivasyonu için 12 tane protein: translasyonda enzim 100 tane protein: posttranslasyonel işlemlerde 100 tane rRNA, mRNA, tRNA -Yaklaşık 300 tane makromolekül → 1 polipeptid zincirinin sentezi için gerekir b) Protein sentezi çok hızlı gerçekleşir. 100 aa’lik polipeptid 5 sn’de c) Hücre içi protein sentezi çok sıkı kontroldedir.Gerektiği kadar protein sentezlenir
 Protein sentezi çok karmaşık bir olaydır. Eukaryotik hücrelerde : 70 tane ribozomal protein. 20 tane protein: aa aktivasyonu için. 12 tane protein: translasyonda enzim. 100 tane protein: posttranslasyonel işlemlerde. 100 tane rRNA, mRNA, tRNA. -Yaklaşık 300 tane makromolekül → 1 polipeptid. zincirinin sentezi için gerekir. b) Protein sentezi çok hızlı gerçekleşir. 100 aa’lik polipeptid 5 sn’de. c) Hücre içi protein sentezi çok sıkı kontroldedir.Gerektiği. kadar protein sentezlenir.")
104
Translasyonu Evreleri
1- AA’lerin aktivasyonu 2- Polipeptid zincirinin başlaması 3- Uzama 4- Sonlanma ve ribozomdan ayrılma 5- Kıvrılma ve işlenme

105
I.evre :AA’lerin aktivasyonu
aa + tRNA + ATP aminoasil-tRNA +AMP + PPi a.asil-tRNA sentetaz Tersinmez reak Gº’ = -7.0 kcal/mol Mg 2+

106
Ör: isolösil-tRNA sentetaz =E
isolösin + ATP + E E-İsölösil-AMP + PPi E-[isolosil- AMP] +tRNA ile → İsolosil-tRNA ile + E + AMP Gº’ = -7 kcal/mol

108
T=ribotimidin =Pseudoüridin DHU=Dihidrouridin tRNA

109
Polipeptid zincirinin başlaması

110
II.evre: Polipeptid zincirinin başlaması 1) RİBOZOMLAR:
 RİBOZOMLAR:")
111
21 tane polipeptid 34 tane polipeptid Prokaryot Eukaryot

112
2) Başlangıç amino asiti
Prokaryotlarda: Peptid zincirinin aminoterminalinde : N-FORMİL METHİONİN bulunur H COO- ı l H-C-N-C-H ll l O CH2 l N-formil CH2 grubu S CH3

113
Bu aa 2 reaksiyonla oluşur
ATP AMP + PP a) Methionin + tRNA fmet methionil- tRNAfmet Mg ++ Methionil –tRNA sentetaz b)Formil grubunun methionil’in amino grubuna transferi N10- Formil tetrahidrofolat + Met-tRNAfmet tetrahidrofolat +fmet-tRNAfmet N10- FH4 transformilaz -Transformilaz serbest methionine formil bağlayamaz.Özgül substratı Met-tRNAfmet tRNAmet :Peptid zinciri içindeki methionine özgü tRNAfmet :Başlangıçtaki formillenmiş methionine özgü.Bu tRNA sadece formil grubu alabilir
 Methionin + tRNA fmet methionil- tRNAfmet. Mg ++ Methionil –tRNA sentetaz. b)Formil grubunun methionil’in amino grubuna transferi. N10- Formil tetrahidrofolat + Met-tRNAfmet tetrahidrofolat +fmet-tRNAfmet. N10- FH4 transformilaz. -Transformilaz serbest methionine formil bağlayamaz.Özgül substratı Met-tRNAfmet. tRNAmet :Peptid zinciri içindeki methionine özgü. tRNAfmet :Başlangıçtaki formillenmiş methionine özgü.Bu tRNA sadece formil. grubu alabilir.")
114
EUKARYOTLARDA: -Ekstramitokondrial ribozomlarda, methioninle sentez başlar. t-RNAmet -Mitokondri ve kloroplastlardaki ribozomlarda, N-formil Met-tRNAfmet ile başlar *Mitokondrilerin bakteriden oluşumu; simbiotik yaşam teorisini destekler

115
B)Polipeptid sentezinin Başlaması
Prokaryotlarda gerekli yapılar; 1) 30 S subuniti (16 s rRNA içerir) 2) Sentezlenilecek polipeptidi kodlayacak mRNA 3) Başlangıç aa-tRNA=N-formil methionil-tRNA fmet 4) Başlama faktörleri BF (IF-1) BF (IF-2) BF (IF-3) 5) GTP
 30 S subuniti (16 s rRNA içerir) 2) Sentezlenilecek polipeptidi kodlayacak mRNA. 3) Başlangıç aa-tRNA=N-formil methionil-tRNA fmet. 4) Başlama faktörleri BF-1 (IF-1) BF-2 (IF-2) BF-3 (IF-3) 5) GTP.")
116
Başlama kopmleksinin oluşumu
3 safhada gerçekleşir.

117
1.safha 30 s ribozom üniti + BF-3 → Bağlanır
(30 s ve 50 s‘ın birleşmeleri engellenir) mRNA + 30 s subünitine → Bağlanır Başlangıç sinyali Başlangıç kodunu 30 s deki 16 s rRNA N-fmet-tRNAfmet deki UAC ile karşıttır İle koplementer baz oluşturur 6-8 tane A ve G A U G 3’ 5’
 mRNA + 30 s subünitine → Bağlanır. Başlangıç sinyali Başlangıç kodunu. 30 s deki 16 s rRNA N-fmet-tRNAfmet deki UAC ile karşıttır. İle koplementer baz. oluşturur. 6-8 tane. A ve G. A U G. 3’ 5’")
118
İşte başlangıç sinyali sayesinde;
a) mRNA 30 s’te doğru yere oturur b) Başlangıçtaki AUG kodonu= N fmet tRNAfmet Zincir içindeki AUG kodonu= met-tRNAmet bağlar
 mRNA 30 s’te doğru yere oturur. b) Başlangıçtaki AUG kodonu= N fmet tRNAfmet. Zincir içindeki AUG kodonu= met-tRNAmet bağlar.")
119
2. safha 30 s subüniti GTP-BF-2 BF-3 + mRNA Nfmet-tRNAfmet
BÜYÜK BAŞLANGIÇ KOMPLEKSi

120
3.safha

121
1- mRNA ‘da AUG kodonu fmet-tRNAfmet’de UAC’nin anti kodonu 2- Ribozomal P noktası başlangıç aasil-tRNA doğru yere oturmasını sağlar

122
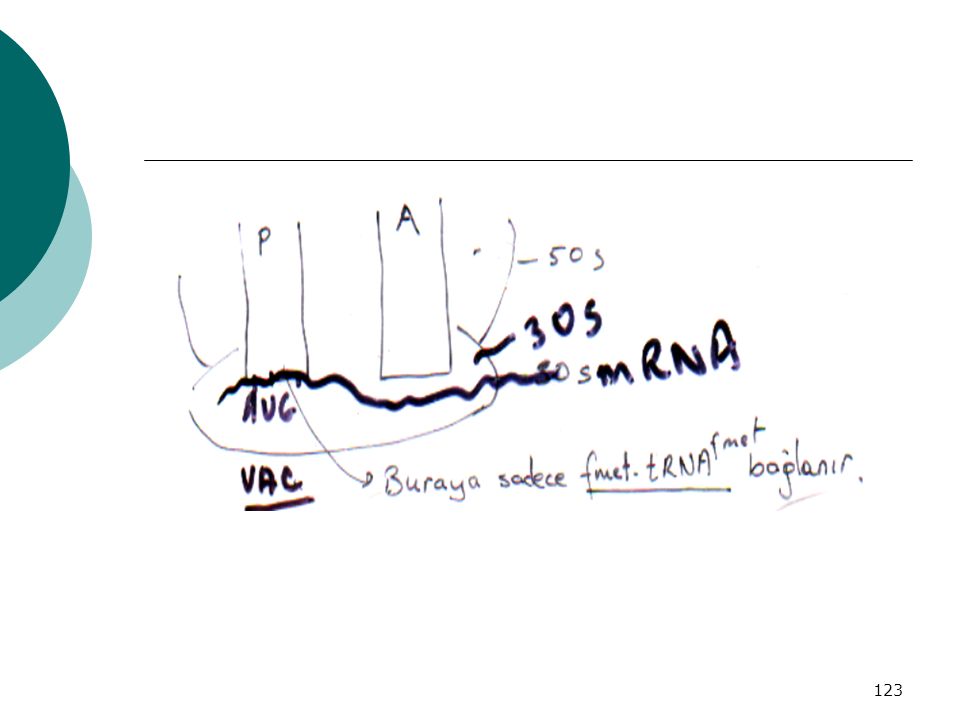
Ribozomlarda aminoasil-tRNA’ları bağlamak için 2 bölge vardır
A Bölgesi : Aminoasil kısmı P Bölgesi : Peptidil kısmı - Bu bölgeler 50 s ve 30 s subunitelerinin spesifik kısımlarından m.g -P bölgesine sadece başlangıç fmet-tRNAfmet bağlanırken A bölgesine ise diğer yeni gelen aminoasil-tRNA’ lar bağlanır

124
İkinci kodon

125
III. Evre:Uzama Gerekli yapılar 1-Başlangıç kompleksi : 70 s ribozom
mRNA fmet-tRNAfmet 2- Bağlanılacak bir sonraki aminoasil-tRNA (mRNA’ da AUG’den sonraki kodona uyan antikodonlu aasil-tRNA)
")
126
3- Uzama Faktörleri Tu Ts G 4- GTP

127
Uzamanın Safhaları 1. safha
Bir sonraki aasil-tRNA + Tu- GTP → aasil-tRNA –Tu-GTP aasil-tRNA-Tu-GTP Tu-GDP+Pi Yeni aasil-tRNA 70 s 70s başlangıç kompleksi (kompleks)
")
128
Rejenerasyon Reaksiyonu
Tu-GDP Tu-GTP GTP GDP Ts

129
Bir sonraki kodon

130
2. Safha: P ve A bölgelerinde oturan aa’ler arasında peptid bağı m.g
50 s subunitindeki; peptidil transferaz enzimi

131
3. safha: TRANSLOKASYON Ribozomun mRNA üzerinde 3’ ucuna doğru
bir kodon kaymasıdır Böylece → A bölgesindeki dipeptidil-tRNA P bölgesine P bölgesindeki- boş tRNA → sitoplazmaya

132
Translokasyonda 1)G= Translokaz (uzama faktörü)
2) GTP GDP + Pi (enerji) gerekir
 GTP GDP + Pi (enerji) gerekir.")
133
Dipeptidil t-RNA2 Translokasyon kademesi A kısmı bir sonraki Aasil t-RNA için hazır

134
IV-Sonlanma ve Ribozomdan Salınma
mRNA’daki şifreye göre son aa takıldıktan sonra SONLANMA KODONLARI; UAA, UAG, UGA ‘dır. Bunlar hiçbir aa kodlamaz Bu kodonlara gelinince 3 tane SONLANMA veya SALINIM FAKTÖRÜ işe karışır R1, R2, S → Sonlanma proteinleridir

135
SONLANMA veya SALINIM FAKTÖR’lerinin işlevleri :
1- Polipeptid zincirini son tRNA’dan ayırma ve polipeptid zincirini sitoplazmaya salma 2- P kısmında boş kalan tRNA’ yı → sitoplazmaya 3- 70 s ribozomu 30s + 50 s’li subunitelere ayırma (Böylece yeni bir polipeptid zinciri sentezlenebilir)
")
136
mRNA Aa kodlayan kodonlar
Sonlanma kodonları Başlama kodonu Başlama sinyali

137
V.Kıvrılma ve İşlenme Posttranslasyonel Modifikasyonlar
-Polipeptid zincirinin biolojik aktif hale gelmesi için geçirdiği değişimler; Sekonder, tersiyer, kuarterner yapıların oluşumu Bazı grupların takılması Bazı grupların çıkarılması

138
PROTEİN SENTEZİNİN DÜZENLENMESİ
1- Transkripsiyonel Kontrol → Bakterilerde 2-Translasyonel Kontrol → Eukaryotlarda (karmaşık, ?)
")
139
TRANSKRİPSİYONEL KONTROL
Bir hücredeki enzimler; A) YAPISAL enzimler: Her hücrenin tipine göre sabit miktarda B) UYARILABİLİR enzimler: Yapımı şartlara göre veya (uyarılabilen- baskılanabilen enzimler)
 YAPISAL enzimler: Her hücrenin tipine göre sabit miktarda. B) UYARILABİLİR enzimler: Yapımı şartlara göre veya (uyarılabilen- baskılanabilen enzimler)")
140
A) BASKILANABİLEN enzim
E.coli → tek N kaynağı NH4+ tuzları → tüm aa’leri sentezler *Ortama histidin ilavesiyle, histidin sentezleyen enzimler baskılanır (son ürün inhibisyonu)
")
141
B)UYARILABİLEN enzim Normalde mikterı az, ama bazı şartlarda yapım miktarı artan enzimler Ör: -Galaktozidaz Laktoz → D-Glukoz + D-Galaktoz E.coli’de - -Galaktozidaz normalde 5-6 tane. - Ortamda glukoz varsa bu enzim hiç kullanılmaz - Tek C kaynağı laktozlu besi yerinde -1-2 dk’da 1000 tane - Galaktozidaz sentezi

142
OPERON: Birbiriyle fonksiyonel olarak ilişkili ve şartlara göre açılıp, kapatılabilen genler grubudur

143
LAC OPERONU Yapısal genler : z → -Galaktozidaz y → permeaz
a → A proteini Düzenleyici gen : i → represör protein Kontrol genleri : p → promoter 0 → operator İndükleyici : allolaktose (Laktozun izomeri)
")
144
DNA’ da Lak operon’unun genetik yapısı

Benzer bir sunumlar

>")


 ZM106 Biyokimya 11. Hafta.>")




>")



 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")
>")





 PC KOPAT>")