Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
DNA ile etkileşen proteinler ve enzimler
BİYOFİZİK A.B.D. Araştırma Gör. Ayhan ÜNLÜ

2
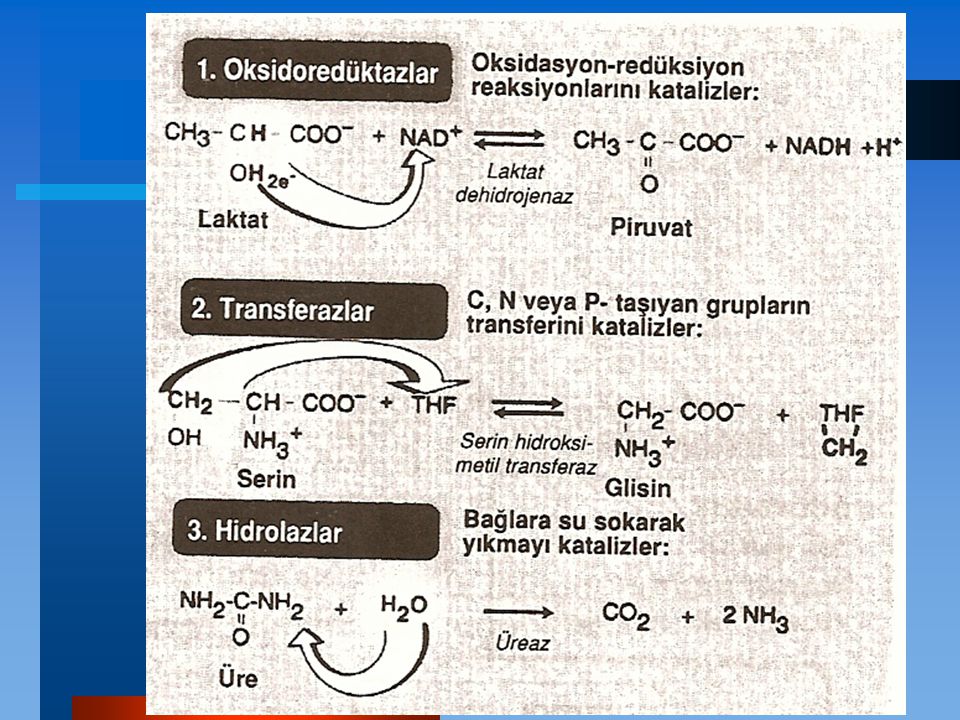
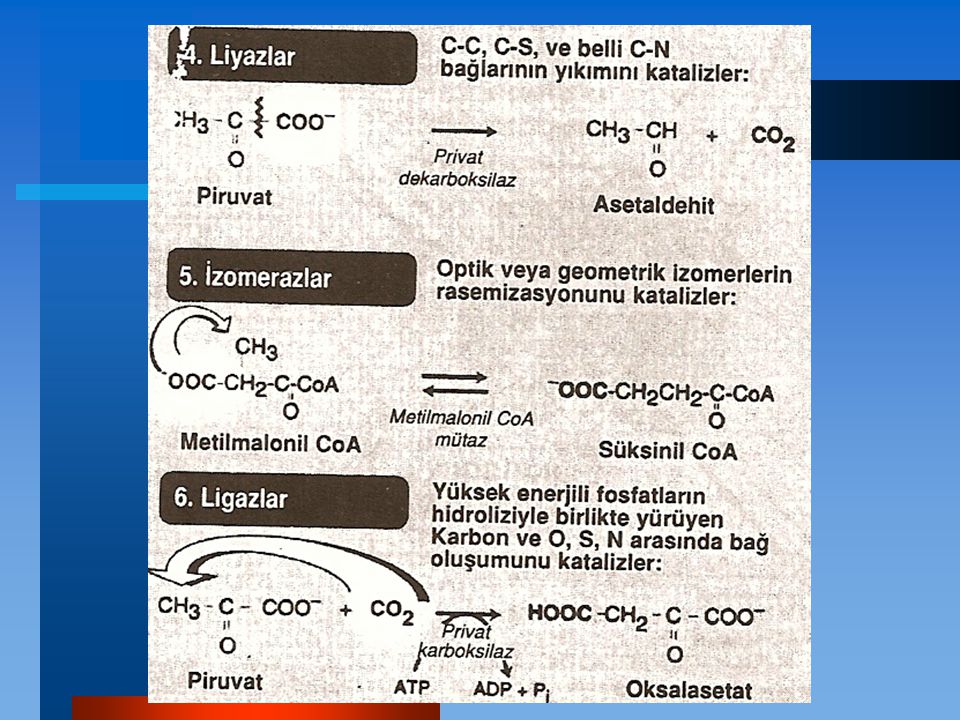
ENZİMLER Vücuttaki tüm reaksiyonlar, enzimler (tüm işlem sonunda kendileri değişmeden reakisyonların hızını arttıran protein katalizörler) tarafından yürütülürler. Enerji açısından mümkün olan bir çok biyolojik reakisyon arasında enzimler reaktanları (substratlar diye adlandırırlar) kullanışlı yollara seçici olarak kanalize ederler. Böylece enzimler bütün metabolik olayları yönlendirirler.
 tarafından yürütülürler. Enerji açısından mümkün olan bir çok biyolojik reakisyon arasında enzimler reaktanları (substratlar diye adlandırırlar) kullanışlı yollara seçici olarak kanalize ederler. Böylece enzimler bütün metabolik olayları yönlendirirler.")
5
Enzimler bir kimyasal reaksiyonun hızını arttıran ve katalizledikleri reaksiyon sırasında tüketilmeyen protein katalizörlerdir.

6
Aktif Bölgeler Enzim moleküllerinde aktif bölge denilen özel bir cep ya da yuva bulunur. Aktif bölge substrata komplemanter olan üç-boyutlu bir yüzey yaratan amino asit yan zincirleri içerir. Aktif bölge substratı bağlayarak bir enzim-substrat (ES) kompleksi" meydana getirir. ES, sonradan enzim ve ürüne parçalanan enzimürün (EP)'e dönüşür.
 kompleksi meydana getirir. ES, sonradan enzim ve ürüne parçalanan enzimürün (EP) e dönüşür.")
7
Katalitik Etkinlik Enzimle katalizlenen reaksiyonların çoğu katalizlenmeyen reaksiyonlara göre 103 ile 108 kere daha hızlı olarak oldukça etkindir. Tipik olarak, her enzim molekülü saniyede 100 ila 1000 substrat molekülünü ürüne çevirme yeteneğine sahiptir. Enzim başına düşen ürüne çevrilmiş substrat molekülü sayısına turnover sayısı denir.

8
Spesifiklik Enzimler bir veya birkaç belirli substratla etkileşerek ve sadece tek tip kimyasal reaksiyon katalizleyerek oldukça spesifiktir.

9
Kofaktörler Bazı enzimler, enzimatik reaksiyon için gerekli olan bir nonprotein kofaktörle birleşirler. Sıklıkla karşılaşılan kofaktörler arasında metal iyonlar (örneğin, Zn2+ Fe2+) ve koenzim olarak bilinen, genellikle vitamin türevi olan (NAD+, FAD, koenzim A) organik moleküller yer alır. Holoenzim, kofaktörüyle birlikte enzimi ifade eder. Apoenzim, holoenzimin protein kısmını ifade eder. Uygun kofaktör yoksa, apoenzim biyolojik aktivite gösteremez.
 ve koenzim olarak bilinen, genellikle vitamin türevi olan (NAD+, FAD, koenzim A) organik moleküller yer alır. Holoenzim, kofaktörüyle birlikte enzimi ifade eder. Apoenzim, holoenzimin protein kısmını ifade eder. Uygun kofaktör yoksa, apoenzim biyolojik aktivite gösteremez.")
10
Düzenleme Enzim aktivitesi düzenlenebilir -yani, enzimler ürün oluşum hızı ve hücrenin ihtiyacını karşılayacak şekilde aktive veya inhibe edilebilirler.

11
Hücre içindeki konumları
Bir çok enzim hücre içindeki spesifik organellerde lokalizedir

12
ENZiMLER NASIL ÇALIŞIR
Enzimlerin etki mekanizmasına iki ayrı açıdan bakılabilir. Birincide, kataliz reaksiyon sırasında meydana gelen enerji değişimi açısından değerlendirilir; enzimler katalizlenmemiş reaksiyondan farklı, enerji açısından tercih edilen alternatif bir reaksiyon yolu sağlarlar. ikincide ise aktif bölgenin kimyasal olarak katalizi nasıl hızlandırdığı tarif edilir.

13
Reaksiyon sırasında meydana gelen enerji değişimleri
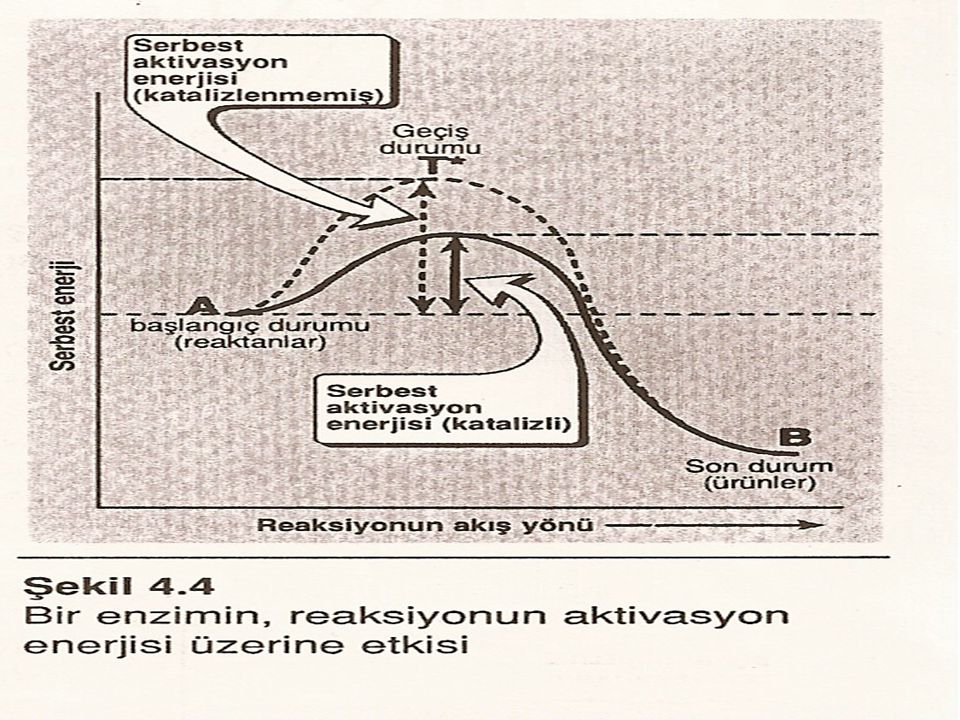
Bütün kimyasal reaksiyonlarda reaktanları ve ürünleri ayıran bir enerji bariyeri vardır. Serbest aktivasyon enerjisi denilen bu bariyer, reaktanlar ile ürün oluşumu sırasında meydana gelen yüksek enerjili ara-ürün arasındaki enerji farkıdır.

15
Serbest aktivasyon enerjisi:
Bu enerji piki reaktanın ürüne dönüşümü sırasında yüksek-enerjili bir araürünün meydana geldiği geçiş durumunu gösterir. Yüksek aktivasyon enerjisinden dolayı katalizlenmeyen reaksiyonların hızları genellikle yavaştır.

16
Reaksiyon hızı: Moleküllerin reaksiyona girmeleri için geçiş durumunun enerji bariyerini aşmaya yetecek kadar enerjiye sahip olmaları gerekir. Reaksiyonun hızı böyle enerjili moleküllerin sayısı ile belirlenir. Genel olarak, serbest aktivasyon enerjisi ne kadar düşükse, o kadar molekül geçiş durumunu aşmaya yeterli enerjiye sahiptir ve böylece reaksiyonun hızı o kadar fazladır.

17
Alternatif reaksiyon yolu:
Bir enzim daha düşük bir serbest aktivasyon enerjisi olan alternatif bir reaksiyon yolu sağlayarak bir reaksiyonun hücre içindeki şartlarda daha hızlı yürümesine olanak verir.

18
Aktif bölgenin kimyası
Aktif bölge substratın bağlanacağı basit bir yuva değildir, tam tersine.substatın ürüne dönüşümünü hızlandıracak bir çok mekanizmayı yürüten karmaşık bir moleküler makinadır.

19
Geçiş durumu stabilizasyonu:
Aktif bölge substratı yapısal olarak molekülün aktive olmuş geçiş durumuna benzeyen bir geometride bağlayan esnek bir moleküler kalıp gibi davranır. Enzim subtratı geçiş durumunda stabilize ederek, ürüne dönüşebilecek reaktif ara ürün konsantrasyonunu büyük miktarlarda arttırır ve böylece reaksiyonu hızlandırır.

20
Diğer faktörler: Aktif bölge geçiş durumunun oluşumunu sağlayan katalitik grupları içerir. Bazı enzimlerde bu gruplar amino asit kalıntılarının proton alıp verdiği genel asit-baz katalizine katılabilir. Bazı enzimlerde ise kataliz geçişi bir kovalent enzimsubstrat kompleksinin meydana gelmesini sağlar.

21
REAKSiYON HIZINI ETKiLEYEN FAKTÖRLER
Enzimler hücrelerden izole edilerek özellikleri test tüpü içinde (in vitro) araştırabilir. Farklı enzimler substrat konsantrasyonu, ısı ve pH'daki değişikliklere farklı olarak yanıt verirler.
 araştırabilir. Farklı enzimler substrat konsantrasyonu, ısı ve pH daki değişikliklere farklı olarak yanıt verirler.")
22
Substrat konsantrasyonu
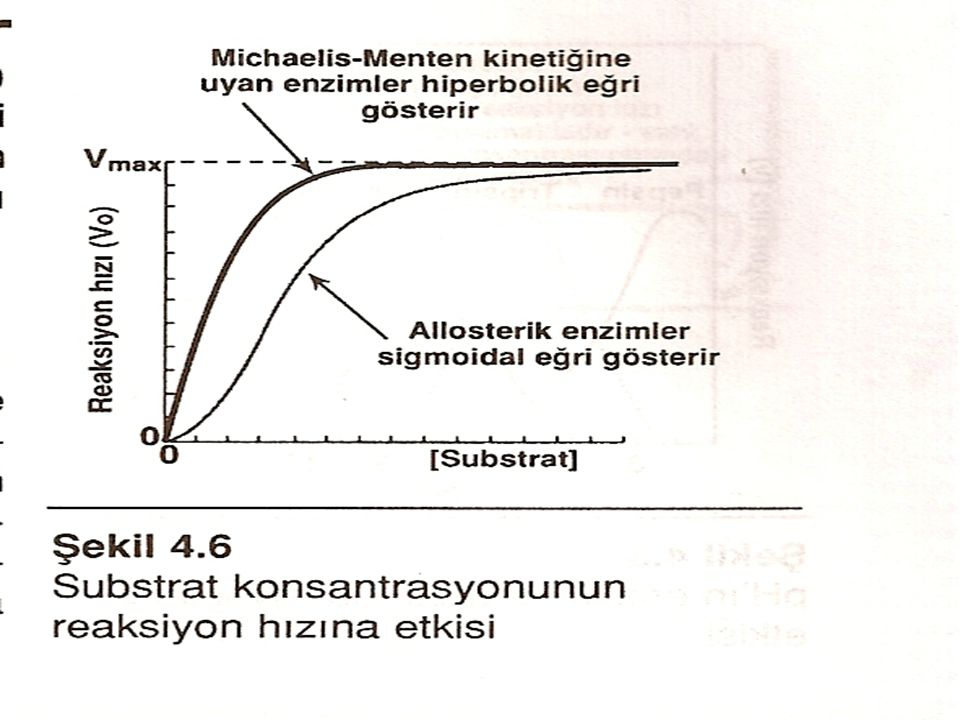
Maksimal hız: Bir reaksiyonun hızı (v) birim zamanda ürüne çevrilen substrat molekülü sayısıdır ve genellikle dakikada oluşan mol ürün olarak ifade edilir. Enzim-katalizli bir reaksiyonun hızı bir maksimum hıza (V max) ulaşana kadar substrat konsantrasyonuyla artar. Reaksiyon hızının dengeye ulaşması enzimin bütün uygun bölgelerinin substratla doyduğuna işaret eder.
 birim zamanda ürüne çevrilen substrat molekülü sayısıdır ve genellikle dakikada oluşan mol ürün olarak ifade edilir. Enzim-katalizli bir reaksiyonun hızı bir maksimum hıza (V max) ulaşana kadar substrat konsantrasyonuyla artar. Reaksiyon hızının dengeye ulaşması enzimin bütün uygun bölgelerinin substratla doyduğuna işaret eder.")
24
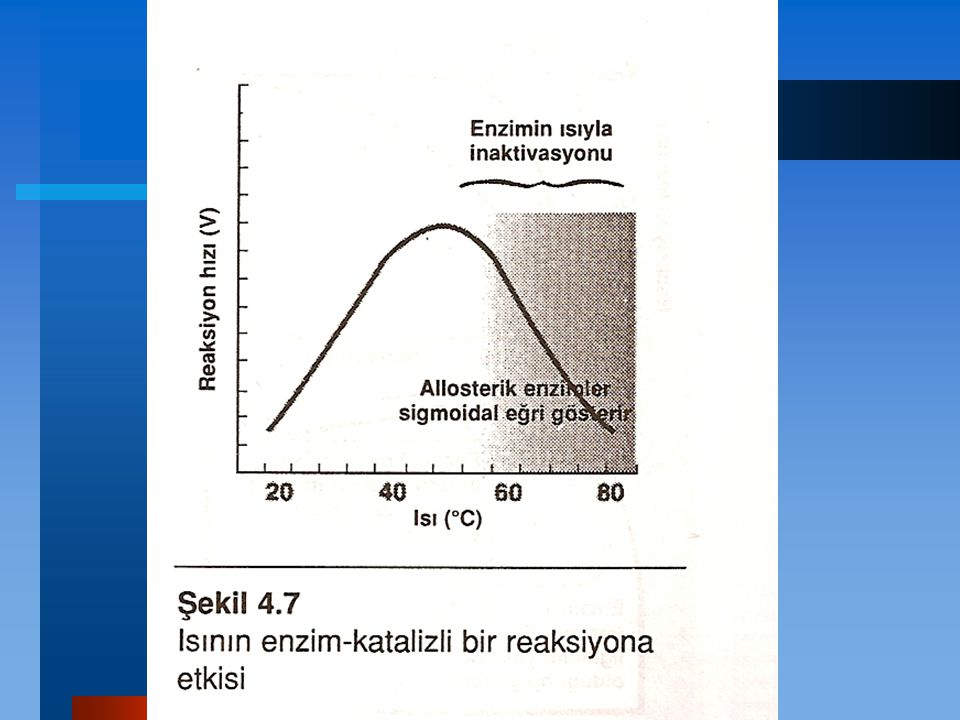
Isı Hızın ısıyla artışı: Reaksiyon hızı maksimum hıza erişilinceye kadar ısıyla artar. Bu artış enerji bariyerini geçip reaksiyon ürünlerini oluşturmaya yetecek enerjiye sahip olan molekül sayısının artmasına bağlıdır. Hızın ısıyla azalması: Isının daha da arttırılması sonucunda enzimin ısıyla denatürasyonuna bağlı olarak reaksiyon hızında bir azalma meydana gelir.

26
pH pH'ın aktif bölgenin iyonizasyonu üzerine etkisi: H+ konsantrasyonu reaksiyon hızını çeşitli yönlerden etkiler. ilk olarak, katalitik işlem jçin genellikle enzim ve substratın spesifik gruplarının reaksiyona girebilmeleri için iyonize veya iyonize olmamış durumda bulunmaları gerekir. Örneğin, katalitik aktivite için enzimin bir amino grubunun protonlanmış durumda (-NH3+) olması gerekebilir. Alkali pH'da bu grup deprotone olur ve bu yüzden reaksiyonun hızı düşer.
 olması gerekebilir. Alkali pH da bu grup deprotone olur ve bu yüzden reaksiyonun hızı düşer.")
27
pH pH'in enzim denatürasyonu üzerine etkisi: Katalitik olarak aktif olan protein molekülünün yapısı amino asit yan zincirlerinin karakterine bağlı olduğu için, pHdaki aşırılıklar da enzimin denatüre olmasına neden olur.

28
MICHAELIS-MENTEN DENKLEMİ
Michaelis ve Menten enzimle katalizlenen reaksiyonların özelliklerinin çoğunu kapsayan basit bir model öne sürmüşlerdir. Bu modelde,' enzim substratıyla geridönüşümlü olarak bağlanarak sonradan enzimi serbest olarak bırakarak ürüne yıkılan bir ES kompleksi meydana getirir. E + S ES E+P

29
Michaelis-Menten denklemi reaksiyon hızının substrat konsantrasyonuyla nasıl değiştiğini göstermektedir:

30
Michaelis-Menten kinetiği hakkında önemli sonuçlar
Km'in özellikleri: Michaleis sabiti bir enzime ve belli bir substrata özeldir ve o enzimin söz konusu substrata olan ilgisini yansıtır. Km sayısal olarak, reaksiyon hızının 1/2 Vmax'a eşit olduğu esnadaki substrat konsantrasyonudur. Km enzim konsantrasyonuyla değişmez.

31
Küçük Km: Sayısal olarak küçük (düşük) Km enzimin substrata ilgisinin yüksek olduğunu gösterir çünkü, enzimi yarı yarıya doyurmak, -yani, 1/2 V max hızına erişmek için, düşük konsantrasyonda substrat yeterli olmaktadır Büyük Km: Sayısal olarak büyük (yüksek) Km enzimin substrata ilgisinin az olduğunu gösterir çünkü, enzimi yarı yarıya doyurmak için yüksek konsantrasyonda substrat gerekmektedir.
 Km enzimin substrata ilgisinin az olduğunu gösterir çünkü, enzimi yarı yarıya doyurmak için yüksek konsantrasyonda substrat gerekmektedir.")
33
GEN AKTİVİTESİNİN DÜZENLENMESİ
Enzim olarak görev yapan bazı proteinlerin yıkım hızı hücrede enzim aktivitesinin düzenlenmesi bakımından çok önemlidir. Bunların anlaşılması mikroorganizmalarla yapılan deneyler sonucu elde edilmiştir. Genlerin yapısı ve etkileri bir prokaryot olan Escherichia coli bakterisinde incelenmiştir.

34
E.coli'nin enzim sistemine ait bilgiler gen düzenlenmesi olayının anlaşılmasına yardımcı olmuştur. E.coIi'nin enzim sentezi, enzim üretilişleri yönünden iki grupta incelenebilir: Düzenlenmemiş Enzim Üretimi Düzenlenmiş Enzim Üretimi Organizmadaki bazı enzimler enzimin etkileyeceği substratın bulunup bulunmadığına bakılmadan sürekli olarak sentezlenir, bu enzimlere yapısal enzimler denir.

35
Düzenlenmiş Enzim Üretimi
Dürtülenebilen (Inducible) Enzimler Baskılanabilen (REpressible) Enzimler Bu enzimler ortamda ancak kendi substratları (yani onları uyaran) olduğu zaman sentezlenir. Bu tip enzimlerin büyük bölümü kendi aktiviteleri bakımından katabolik (yıkıcı) olup dürtülenebilen enzimler adını alırlar.
 Enzimler. Baskılanabilen (REpressible) Enzimler. Bu enzimler ortamda ancak kendi substratları (yani onları uyaran) olduğu zaman sentezlenir. Bu tip enzimlerin büyük bölümü kendi aktiviteleri bakımından katabolik (yıkıcı) olup dürtülenebilen enzimler adını alırlar.")
36
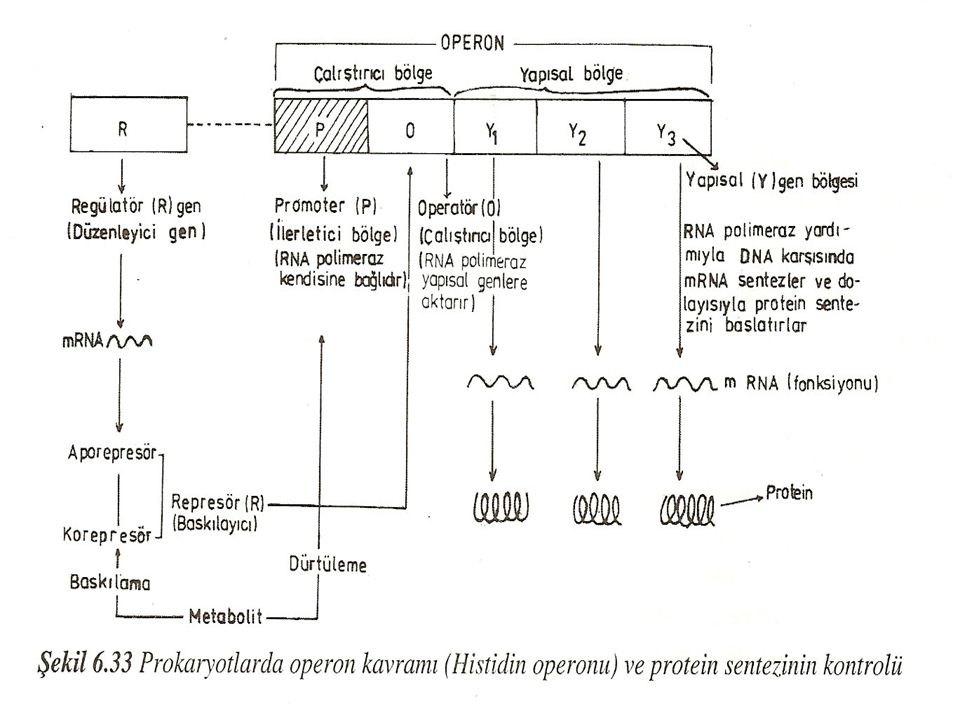
Baskılayıcı (Repressible) Enzimler
Bunlar hücrelerde normal olarak bulunur. Fakat bu enzimlerin reaksiyon sonucu oluşturduğu son ürünlerin konsantrasyonu artınca sentez işlemine son verilir. Son ürünlere korepresör denir. Bir de düzenleyici genler tarafından oluşturulan aporepresör maddesi vardır. Aporepresör, korepressör ile birleşerek fonksiyonel baskılayıcı represör molekülü ortaya çıkarır. Represör moleküller, gerekli enzimleri sentezleten genlerden mRNA kopyasının çıkartılmasını önler ve sonuçta protein sentezi durur. Baskılanabilen enzimler genellikle sentez (anabolik) reaksiyonlarda görev yapar
 reaksiyonlarda görev yapar.")
38
RNA Polimeraz Enzimleri:
Ökaryotik hücrelerde üç tür nüklear RNA polimeraz enzimi vardır. Bunların her biri farklı tipteki genlerin transkripsiyonunu yapar. RNA polimeraz I büyük ribozamal RNA'ların, RNA polimeraz II mRNA'ların, RNA polimeraz III ise küçük 5SrRNA ile tRNA'ların transkripsiyonunda rol alır.

39
DNA’nın Replikasyonu DNA'nın hücredeki önemli görevlerinden biri, hücre bölünmesi esnasında zorunlu olarak kendisini eşleyerek (ki buna replikasyon denir) eşit miktarda iki hücreye dağılmasıdır. Bu olayda DNA polimeraz ve DNA ligaz gibi enzimler rol oynar. Her hücre bölünmesinde DNA replikasyonu olur. Ökaryot kromozomunda replikasyon bir çok noktada başlayarak (100 kadar) DNA'nın iki kolu birbirinden ayrılır. Bunun için DNA çift zincirinde karşılıklı gelen nukleotidlerin bazları arasındaki zayıf hidrojen bağları koparak zincir o noktada ikiye ayrılır.
 eşit miktarda iki hücreye dağılmasıdır. Bu olayda DNA polimeraz ve DNA ligaz gibi enzimler rol oynar. Her hücre bölünmesinde DNA replikasyonu olur. Ökaryot kromozomunda replikasyon bir çok noktada başlayarak (100 kadar) DNA nın iki kolu birbirinden ayrılır. Bunun için DNA çift zincirinde karşılıklı gelen nukleotidlerin bazları arasındaki zayıf hidrojen bağları koparak zincir o noktada ikiye ayrılır.")
40
Ayrılma tamamlanınca bu dallar kalıp görevi yapar
Ayrılma tamamlanınca bu dallar kalıp görevi yapar. Nükleusta mevcut olan nükleotidler kalıp dalların karşısına, uygun nukleotidler halinde DNA polimeraz III enzimi yardımıyla aralarında hidrojen bağlan oluşarak, sıralanırlar. Böylece DNA çift sarmalından birbirinin tamamen aynı olan iki yavru zincir (sarmal) meydana gelir
 meydana gelir.")
41
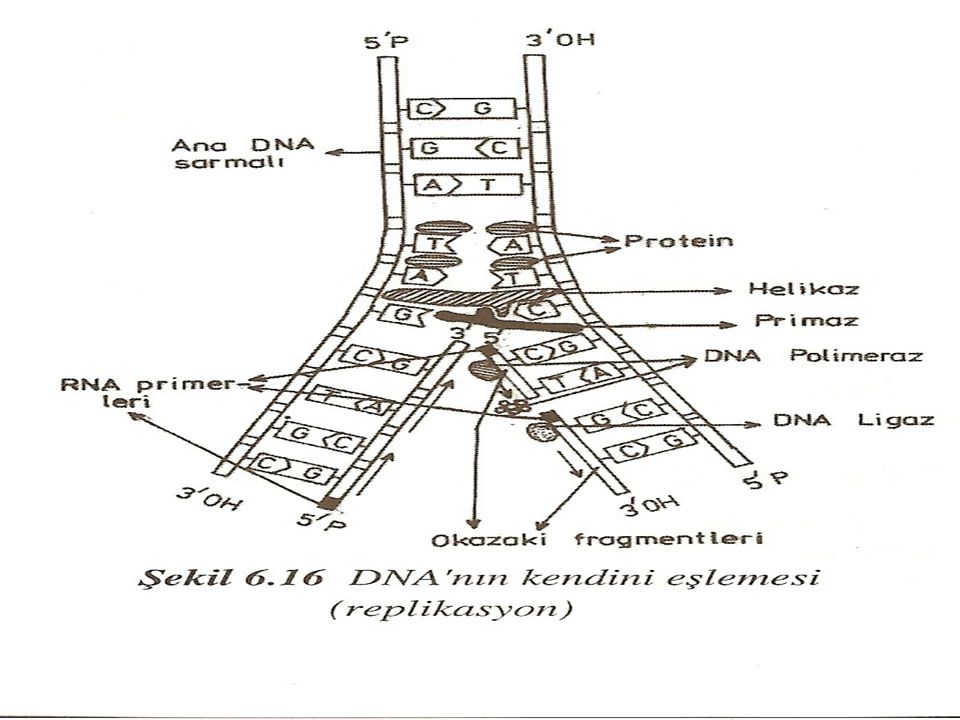
DNA sarmalında yer alan elemanların fonksiyonları
Primaz: Bir çeşit RNA polimeraz enzimi olup RNA primerlerini sentezletir. RNA primeri: Ökaryotlarda DNA primaz tarafından sentezlenen 10 kadar RNA nükleotidinden ibaret olup, DNA replikasyonunda, 3'-OH 5'-P DNA kolunun karşısına, 5' - P 3' -OH istikametinde uygun nükleotidlerin gelmesini sağlar. Kesikli olarak sentezlenen dalda her kesinti noktasında RNA primeri bulunur. Görevi biten RNA primeri ayrılır ve kesinti bölgesi, DNA polimerazın yeni nükleotidleri eklemesiyle doldurulur. Son olarak kesikli noktalardaki 3'OH ile 5'P uçlan DNA ligaz yardımıyla birleştirilir.

42
DNA sarmalında yer alan elemanların fonksiyonları
Helikaz: DNA kollarının açılmasını sağlayan bir enzimdir. Protein molekülü (=Tek dala bağlanan protein) (Single Stranded DNA Binding Protein): Açılan DNA kollarının birleşmesini önler. Böylece tek dalın kopyalanmaya hazırlanmasını sağlar.
 (Single Stranded DNA Binding Protein): Açılan DNA kollarının birleşmesini önler. Böylece tek dalın kopyalanmaya hazırlanmasını sağlar.")
43
DNA sarmalında yer alan elemanların fonksiyonları
DNA polimeraz: Nükleotidlerin karşılıklı bağlanarak DNA sentezini sağlar.

44
DNA sarmalında yer alan elemanların fonksiyonları
DNA ligaz: Tek polipeptid zincirinden ibarettir. Prokaryot ve ökaryotlarda bulunur. Sağ kalıp kolda 5'-P 3'-OH istikametinde kesikli olarak sentezlenen nükleotidleri (Okazaki fragmentleri) birleştirir. Daha genel anlamda, DNA ligaz; DNA'nın herhangi bir kırık bölgesinde bulunan serbest 3'-OH grubu ile aynı zincirin diğer ucunda bulunan 5'- fosfat grubu arasında bir fosfodiester bağı yaparak kırığı kapatır. Bu görevini ATP veya diğer bir enerji kullanarak yapar. DNA ligaz, tek zincirli DNA moleküllerini birbirine bağlamaz. Aksine çift zincirli DNA'da, kırılan tek zincirin iki ucunu birbirine bağlar.
 birleştirir. Daha genel anlamda, DNA ligaz; DNA nın herhangi bir kırık bölgesinde bulunan serbest 3 -OH grubu ile aynı zincirin diğer ucunda bulunan 5 - fosfat grubu arasında bir fosfodiester bağı yaparak kırığı kapatır. Bu görevini ATP veya diğer bir enerji kullanarak yapar. DNA ligaz, tek zincirli DNA moleküllerini birbirine bağlamaz. Aksine çift zincirli DNA da, kırılan tek zincirin iki ucunu birbirine bağlar.")
46
DNA Polimeraz I 1957'de Arthur Kornberg ve arkadaşları, E. coli'den hücreden-arı (in vitro) sistemde DNA sentezini yönlendiren bir enzimİ ayrıştırmıştır. Benzer enzimlerin ilki olan bu enzim bugün, DNA Polimeraz I olarak bilinmektedir.
 sistemde DNA sentezini yönlendiren bir enzimİ ayrıştırmıştır. Benzer enzimlerin ilki olan bu enzim bugün, DNA Polimeraz I olarak bilinmektedir.")
47
Kornberg, DNA polimeraz I'in varlığında in vitro DNA sentezi için başlıca iki gereksinim olduğunu saptamıştır: Dört tip deoksiribonükleozit trifosfat (dATP, dCTP, dGTP, dTTP = dNTP) DNA kalıbı
 DNA kalıbı.")
48
Kornberg'in enzimi ile yönlendirilen sentez, çoğunlukla yarı-saklı replikasyonda olması gereken biçimde cereyan etmiştir. Daha sonraki çalışmalar ile enzimin 928 amino asit içeren tek bir polipeptit zincirinden oluştuğu gösterilmiştir.

50
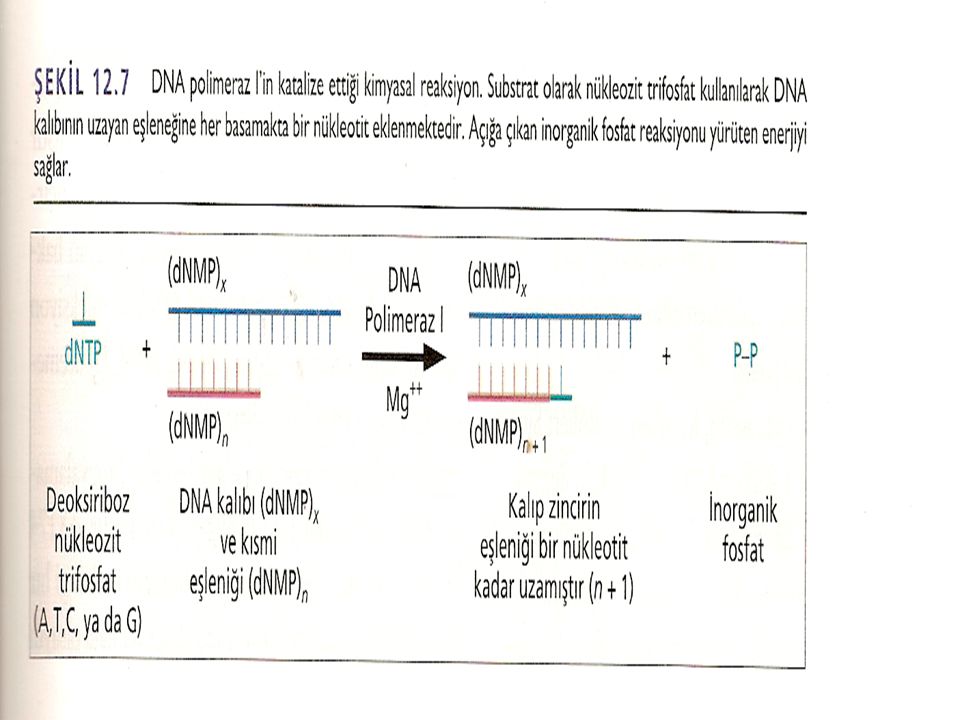
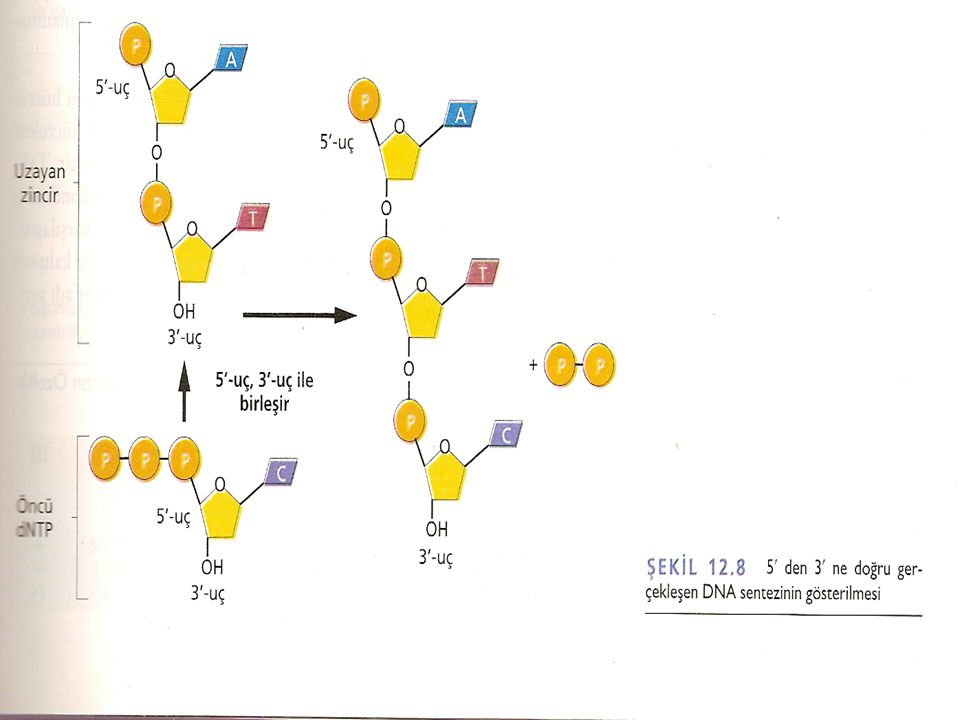
Uzayan DNA zincirine her bir nükleotitin katılım şekli DNA polimeraz I'in özgüllüğüne bağlıdır. Öncü dNTP'de, d-ribozun 5'karbonuna üç adet fosfat grubu bağlıdır. Sentez esnasında, uçta ki iki fosfat grubu koparken, 5' -karbona bağlı olan fosfat grubu, ilave edileceği d-ribozun 3'OH'ına kovalent bağla bağlanır. Böylece zincir uzaması, uzayan zincirin 3' ucuna her seferinde bir nükleotit ilavesi ile 5'3' yönünde devam eder. DNA sentezi devam ettikçe, her basamakta açığa çıkan yeni 3'-OH grubu, DNA sentezi ilerlerken sonraki nükleotitin zincire eklenmesini sağlar.

52
1967' de, Mehran Goulian, Kornberg ve Robert Sinsheimer, küçük bir bakteriyofaj olan X174 ile deneyler yapmıştır. X174 fajı tek zincirli, halkasal ve küçük bir genetik materyale (5386 nükleotit) sahip olduğu için, ideal bir deneysel sistemdir. Molekül kapalı halkasal yapıda olduğundan, replikasyondan sonra sentezlenmiş olan doğrusal DNA zincirinin iki ucunun DNA ligaz (polinükleotitleri birleştiren enzim olarak da bilinir) adı verilen başka bir enzimle birleştirilmesi gerekir.
 sahip olduğu için, ideal bir deneysel sistemdir. Molekül kapalı halkasal yapıda olduğundan, replikasyondan sonra sentezlenmiş olan doğrusal DNA zincirinin iki ucunun DNA ligaz (polinükleotitleri birleştiren enzim olarak da bilinir) adı verilen başka bir enzimle birleştirilmesi gerekir..")
53
DNA Polimeraz II ve III Polimeraz I'in sentezini yönlendirdiği DNA'nın biyolojik aktivitesi saptanmış olsa da, 1969'da enzimin gerçek biyolojik rolü hakkında ciddi şüpheler oluşmuştur. Peter DeLucia ve John Cairns, DNA polimeraz I aktivitesine sahip olmayan bir mutant Ecoli suşu bulmuştur. Bu mutasyon polA1 olarak adlandırılmıştır. İşlevsel enzime sahip olmayan bu mutant E coli suşu DNA'sını kopyalayıp üremeyi başarmıştır, ancak hücreler DNA "onarım" yeteneği bakımından oldukça yetersiz kalmıştır. Örneğin, mutant suş DNA hasarı yapan ve mutajenik olan ultraviyole ışığına (UV) ya da radyasyona son derece hassastır. Mutant olmayan bakteriler, W-hasarını önemli miktarda tamir edebilmektedirler.
 ya da radyasyona son derece hassastır. Mutant olmayan bakteriler, W-hasarını önemli miktarda tamir edebilmektedirler.")
54
Bu gözlemlerden iki sonuç ortaya çıkmışır:
E coli'de, in vivo DNA replikasyonu yapabilen başka bir enzim bulunmalıdır. DNA polimeraz I'in in vivo koşullarda ikincil bir işlevi olabilir. Bu gözlemler, araştırıcıların, DNA polimeraz I'in DNA sentezinin doğru yapılmasından sorumlu olduğuna, ancak tamamlayıcı zinciri sentezleyen gerçek enzim olmadığına inanmalarına neden olınuştur.

55
Bugüne kadar, polimeraz i aktivitesi olmayan hücrelerden ve polimeraz I aktivitesi içeren normal hücrelerden, iki özgün DNA polimeraz daha elde edilmiştir.

56
Bu üç enzimin hiçbiri, bir kalıptan DNA sentezini başlatamaz, ancak üçü de primer adı verilen, var olan bir DNA zincirini kalıp boyunca uzatabilir. DNA sentezinin başlangıcında primer olarak RNA kullanılmaktadır

57
DNA polimeraz enzimlerinin hepsi molekül ağırlığı 100
DNA polimeraz enzimlerinin hepsi molekül ağırlığı daltonun üzerinde olan büyük protein kompleksleridir. Her üçünün de 3'- 5' ekzonükleaz aktivitesi bulunur. Bu özellik, enzimlerin polimerizasyonu tek yönde gerçekleştirme, bir an duraksayıp, geri dönerek ilave edilen nükleotideri çıkarabilme kapasitelerini ifade etmektedir. Bu aktivite enzime yeni sentezlenen DNA'da hata olup olmadığını kontrol etme (proofread) ve yanlış girmiş nükleotideri doğrusu ile değiştirebilme yeteneğini vermektedir. Hücrede, polimeraz I, polimeraz III'e göre daha fazla miktarda bulunur ve daha dayanıklıdır.
 ve yanlış girmiş nükleotideri doğrusu ile değiştirebilme yeteneğini vermektedir. Hücrede, polimeraz I, polimeraz III e göre daha fazla miktarda bulunur ve daha dayanıklıdır.")
58
Üç polimerazın in vivo rolleri
Polimeraz I, primeri uzaklaştırır ve primerler uzaklaştıkça doğal olarak oluşan boşluklarda DNA sentezleyerek bu bölgeleri doldurur. Ekzonükleaz aktivitesi ile bu işlem esnasında oluşabilecek hataları da onarır. Polimeraz II, ultraviyole gibi dış etmenler sonucu hasar gören DNA'nın onarımında rol almaktadır. Enzim, replikasyon çatalındaki DNA sentezi bozulduğunda aktive olan bir gen tarafından şifrelenmektedir. Replikasyonda gerekli olan ve polimerizasyondan sorumlu asıl enzim olarak Polimeraz III görülmektedir. Enzimin 3'-5'· ekzonükleaz aktivitesi, sentez sırasında hata onarımı işlevini görmesini sağlamaktadır.

59
DNA Polimeraz III Enzimin haloenzim olarak adlandırılan aktif formu, iki takım, 10 farklı polipeptit zincirinden meydana gelmiş bir dimerdir. Molekül ağırlığı daltondan fazladır. Molekül ağırlığı dalton olan en büyük alt birim olan , ve alt birimleri ile beraber holoenzimin polimerizasyon aktivitesi gösteren "çekirdek" (core) enzim kısmını oluşturur. Kalıp zincirin nükleotit polimerizasyonundan alt birimi sorumludur. Çekirdek enzimin alt birimi, 3'-5' ekzonükleaz aktivitesi gösterir.
 enzim kısmını oluşturur. Kalıp zincirin nükleotit polimerizasyonundan alt birimi sorumludur. Çekirdek enzimin alt birimi, 3 -5 ekzonükleaz aktivitesi gösterir.")
61
DNA Sarmalının Açılması
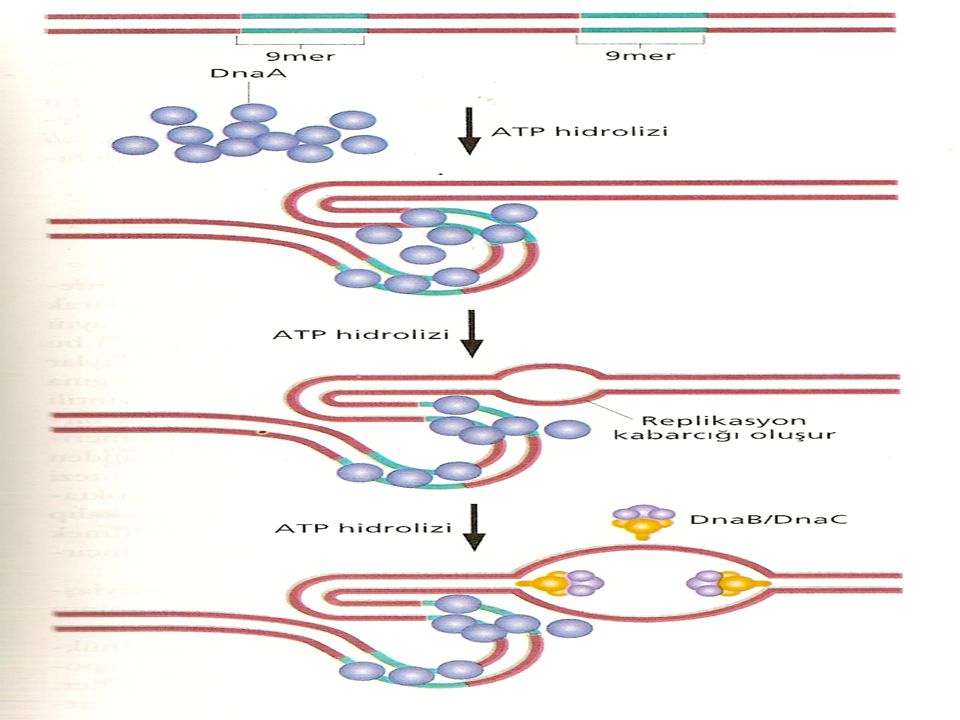
Çoğu bakteri ve virüslerin halkasal kromozomlarında, DNA sentezinin başladığı bir orijin noktası bulunur. Replikasyon orijini olan oriC, 9 ve 13 bazdan oluşan (9mer ve 13mer olarak adlandırılır) tekrar dizilerinin bulunduğu 245 baz çifti içerir. DnaA denen özgül bir protein (dnaA geni tarafından şifrelenir) ilk basamakta sarmalın açılmasından sorumludur. DnaA proteininin bazı alt birimleri birçok 9mer dizisine bağlanır. Bu bağlanma, sarmalın daha fazla açılmasında ve kararlılığında rol alan DnaB ve DnaC proteinlerinin bağlanmasını kolaylaştırır. Hidrojen bağlarını kırıp ikili sarmalı denatüre etmek için normalde ATP hidrolizi ile sağlanan enerjiye gereksinim duyan bu tip proteinler helikazlar olarak adlandırılır. Tek zincire bağlanan proteinler (singlestranded binding proteins, SSBP) olarak bilinen diğer bazı proteinler bu konformasyonu daha da kararlı kılar.
 tekrar dizilerinin bulunduğu 245 baz çifti içerir. DnaA denen özgül bir protein (dnaA geni tarafından şifrelenir) ilk basamakta sarmalın açılmasından sorumludur. DnaA proteininin bazı alt birimleri birçok 9mer dizisine bağlanır. Bu bağlanma, sarmalın daha fazla açılmasında ve kararlılığında rol alan DnaB ve DnaC proteinlerinin bağlanmasını kolaylaştırır. Hidrojen bağlarını kırıp ikili sarmalı denatüre etmek için normalde ATP hidrolizi ile sağlanan enerjiye gereksinim duyan bu tip proteinler helikazlar olarak adlandırılır. Tek zincire bağlanan proteinler (singlestranded binding proteins, SSBP) olarak bilinen diğer bazı proteinler bu konformasyonu daha da kararlı kılar.")
63
Sarmalın açılması devam ettikçe, replikasyon çatalının önünde oluşan sarılma gerilimi çoğu kez üstün kıvrılma (supercoiling) meydana getirir. Halkasal moleküllerde üstün kıvrılmalar, DNA'daki ek bükülmeler ve dönüşler sonucu oluşturulur. Bu durum aynen bir lastiğin uzatılıp, bir ucundan büküldüğünde ortaya çıkan sarmal yapıya benzer. DNA topoizomerazlar olarak adlandırılan geniş bir enzim ailesinin üyesi olan DNA giraz (DNA gyrase) enzimi, bu tip üstün kıvrılmaları gevşetir. Giraz enzimi tek zincirde ya da her iki zincirde "kırıklar" oluşturur, aynı zamanda üstün kıvrılma oluşumu sırasında meydana gelen bükülmeleri ve düğümleri "açma" hareketlerini de katalizler. Oluşan kırıklar sonra tekrar birleştirilir. Bu çeşit reaksiyonlarda ATP hidrolizinden açığa çıkan enerji kullanılır.
 enzimi, bu tip üstün kıvrılmaları gevşetir. Giraz enzimi tek zincirde ya da her iki zincirde kırıklar oluşturur, aynı zamanda üstün kıvrılma oluşumu sırasında meydana gelen bükülmeleri ve düğümleri açma hareketlerini de katalizler. Oluşan kırıklar sonra tekrar birleştirilir. Bu çeşit reaksiyonlarda ATP hidrolizinden açığa çıkan enerji kullanılır..")
64
PCR’da kullanılan enzimler

65
Termostabil (sıcaklığa dayanıklı) DNA polimeraz enziminin PCR'de kullanılmaya başlanması araştırma ve klinik laboratuvarıarında rutin olarak yapılan deneylere teknolojik olarak büyük bir avantaj sağlamıştır. Önceleri kullanılan Escherichia coli'nin DNA polimeraz I enziminin Klenow fragmenti sıcaklığa dayanıklı olmadığı için, her PCR döngüsünde denatürasyon aşamasından sonra yeniden enzim ekleme zorunluluğu duyulmaktaydı. Ancak günümüzde kullanılan termostabil DNA polimerazlarla bu sorun ortadan kalktığından enzimin amplifikasyonun başlangıcında tüplere konulması yeterli olmaktadır.

67
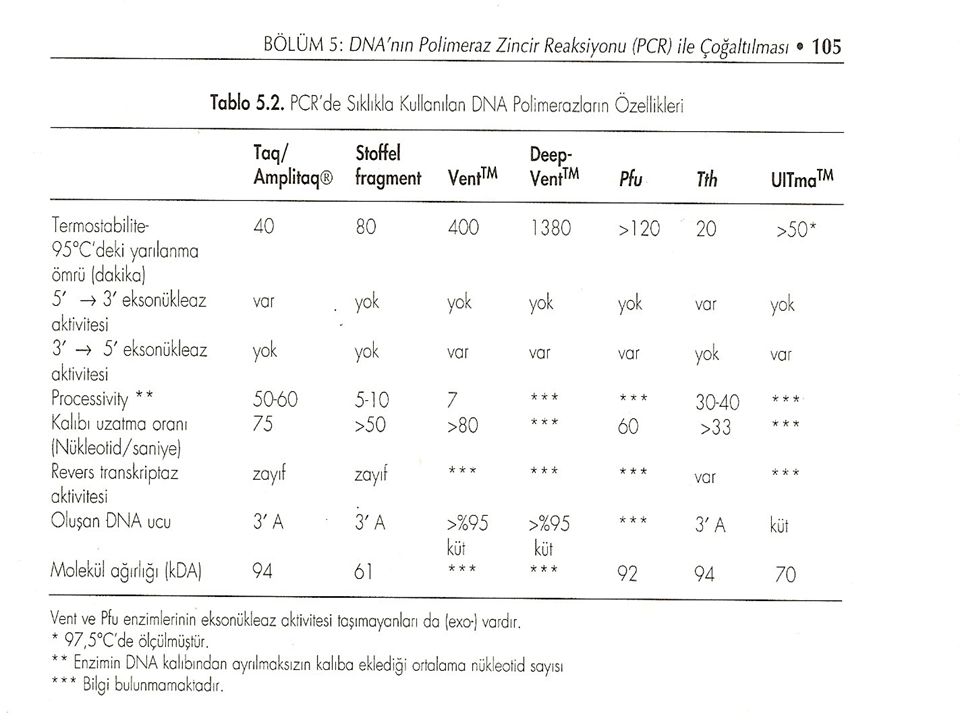
Termostabii DNA polimerazlardan PCR'de en yaygın olarak kullanılanı Thermus aquaticus'dan elde edilen Taq DNA polimerazdır. Taq DNA polimeraz'ın polimerizasyon oranı (nükleotid/saniye) enzim için en uygun sıcaklık olan 70-80°C'da 'dür. Bu enzimin kalıptan ayrılmaksızın kalıba ilave ettiği ortalama nükleotid sayısı (processivity) da yüksektir. Taq DNA polimeraz ve Amplitaq® zincirin uzayan ucundan nükleotidleri uzaklaştıran 5' 3' eksonükleaz aktivitesine sahiptir. Amplitaq®, Taq DNA polimerazın E. coli'de klonlanmış ve modifiye edilmiş rekombinant formudur. Amplitaq®'ın özellikleri Taq DNA polimerazınki ile aynıdır, ama Amplitaq rekombinant olduğundan saflığı Taq DNA polimerazdan daha. fazladır ve PCR'de tercih edilir. Ancak bakteriyal DNA'ların çoğaltılmasında Amplitaq yerine Taq DNA polimeraz kullanımı daha güvenlidir. Çünkü Amplitaq® E. colide klonlandığından bakteriye ait kontaminant DNA taşıyabilir. Bununla birlikte özellikle bakteri DNA'larının çoğaltılması amacıyla Perkin-Elmer Cetus firması düşük DNA içerikli özel bir Amplitaq®,LD üretmiştir.
 enzim için en uygun sıcaklık olan 70-80°C da dür. Bu enzimin kalıptan ayrılmaksızın kalıba ilave ettiği ortalama nükleotid sayısı (processivity) da yüksektir. Taq DNA polimeraz ve Amplitaq® zincirin uzayan ucundan nükleotidleri uzaklaştıran 5 3 eksonükleaz aktivitesine sahiptir. Amplitaq®, Taq DNA polimerazın E. coli de klonlanmış ve modifiye edilmiş rekombinant formudur. Amplitaq® ın özellikleri Taq DNA polimerazınki ile aynıdır, ama Amplitaq rekombinant olduğundan saflığı Taq DNA polimerazdan daha. fazladır ve PCR de tercih edilir. Ancak bakteriyal DNA ların çoğaltılmasında Amplitaq yerine Taq DNA polimeraz kullanımı daha güvenlidir. Çünkü Amplitaq® E. colide klonlandığından bakteriye ait kontaminant DNA taşıyabilir. Bununla birlikte özellikle bakteri DNA larının çoğaltılması amacıyla Perkin-Elmer Cetus firması düşük DNA içerikli özel bir Amplitaq®,LD üretmiştir..")
Benzer bir sunumlar












>")



>")
>")
 Nedir? Lipitlerdir.>")



 PC KOPAT>")