Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
PROTEİNLERİN YAPILARINDAKİ BAĞLAR:
Proteinlerin yapılarında kovalent bağlar ve kovalent olmayan bağlar vardır. Proteinlerin yapılarındaki kovalent bağlar: peptit bağları ve disülfid bağlarıdır; Proteinlerin yapılarındaki kovalent olmayan bağlar: hidrojen bağları, iyon bağları ve hidrofob bağlar (apolar bağlar)’dır.
’dır.")
2
1) Peptit bağları: Bir amino asidin α-karboksil karbonu ile bir başka amino asidin α-amino azotu arasında oluşan C-N bağlarıdır: Bir C−N tek bağının uzunluğu 1,49 Ao (149 pm) ve bir C=N çift bağının uzunluğu 1,27 Ao(127 pm) olduğu halde peptit bağının uzunluğu 1,32 Ao (132 pm) kadardır. Peptit bağının uzunluğunun C=N çift bağının uzunluğundan büyük C−N tek bağının uzunluğundan küçük olması nedeniyle peptit bağının kısmen çift bağ olduğu kabul edilir.
 ve bir C=N çift bağının uzunluğu 1,27 Ao(127 pm) olduğu halde peptit bağının uzunluğu 1,32 Ao (132 pm) kadardır. Peptit bağının uzunluğunun C=N çift bağının uzunluğundan büyük C−N tek bağının uzunluğundan küçük olması nedeniyle peptit bağının kısmen çift bağ olduğu kabul edilir.")
3
Bunun da nedeni, peptit bağlarında rezonans veya mezomeri denen durum olmasıdır:
Peptit bağlarındaki rezonans veya mezomeri nedeniyle C- N bağı %50 çift bağ niteliği kazanmıştır. Peptit bağındaki altı atom düzlemseldir. Karbonil grubunun oksijen atomu ve amit grubundaki hidrojen atomu birbirlerine trans pozisyonda olacak şekilde yerleşmişlerdir.

4
Çift bağların eksen etrafında dönmeleri sınırlı olduğundan, peptit bağı oluşumuna katılan grupların atomları (3C, O, N ve H atomları) bir düzlemde bulunurlar; peptit bağı, rijit ve düzlemseldir: Peptit bağının iki yanındaki α-karbon atomlarının tek bağ etrafında dönüş yapmalarının sonucu olarak polipeptit zincirde art arda gelen küçük peptit düzlemleri aynı düzlemde bulunmazlar:

5
2) Disülfid bağları: İki sistein kalıntısı arasında, tiyol, -SH gruplarının H kaybetmeleri sonucu oluşan S-S bağlarıdır. Cysteine-SH + HS-Cysteine < > Cysteine-S-S-Cysteine (Sistin) Disülfid bağlarının bir protein molekülünün şeklinin oluşmasında ve korunmasında önemli etkisi vardır. Disülfid bağları, bir polipeptit zinciri içerisinde kurulabilir veya çeşitli polipeptit zincirlerinin birbirine bağlanmasını sağlayabilir. Disülfid bağları, ribonükleaz, oksitosin ve vazopressinde aynı polipeptit zincirinde bulunur; insülinde ise iki ayrı polipeptit zincirini birbirine bağlar.
 Disülfid bağlarının bir protein molekülünün şeklinin oluşmasında ve korunmasında önemli etkisi vardır. Disülfid bağları, bir polipeptit zinciri içerisinde kurulabilir veya çeşitli polipeptit zincirlerinin birbirine bağlanmasını sağlayabilir. Disülfid bağları, ribonükleaz, oksitosin ve vazopressinde aynı polipeptit zincirinde bulunur; insülinde ise iki ayrı polipeptit zincirini birbirine bağlar.")
6
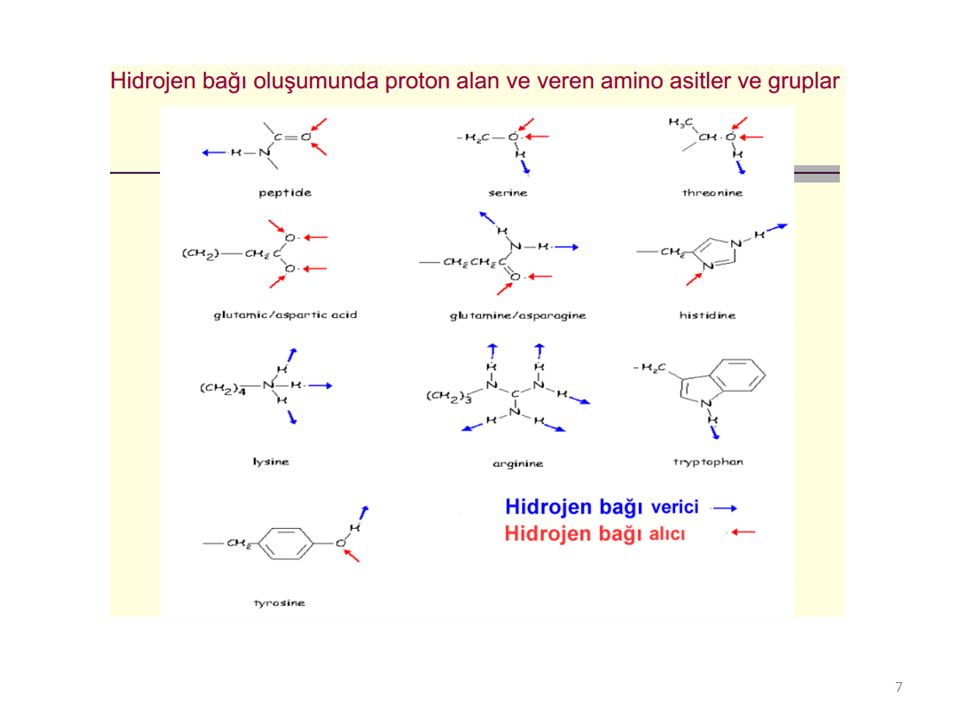
3) Hidrojen bağları: Polipeptit zinciri oluşturan peptit bağlarındaki rezonans veya mezomeri durumundan dolayı, oksijenlerin bilinen keto gruplarından daha negatif, azotların ise pozitif özellik taşımasının sonucu olarak, bir polipeptit zincirdeki bir peptit düzleminde bulunan oksijen atomu ile bir başka peptit bağı veya düzlemindeki azot H atomu arasında, aradaki uzaklık yaklaşık 2,7 Ao olduğunda, hidrojen köprüsü şeklinde (C=O⋅⋅⋅H⋅⋅⋅N) oluşan bağlardır.
 Hidrojen bağları: Polipeptit zinciri oluşturan peptit bağlarındaki rezonans veya mezomeri durumundan dolayı, oksijenlerin bilinen keto gruplarından daha negatif, azotların ise pozitif özellik taşımasının sonucu olarak, bir polipeptit zincirdeki bir peptit düzleminde bulunan oksijen atomu ile bir başka peptit bağı veya düzlemindeki azot H atomu arasında, aradaki uzaklık yaklaşık 2,7 Ao olduğunda, hidrojen köprüsü şeklinde (C=O⋅⋅⋅H⋅⋅⋅N) oluşan bağlardır.")
8
4) İyon bağları: Polipeptit zincirlerindeki asidik ve bazik amino asit kalıntılarının fonksiyonel gruplarının fizyolojik pH’da tamamen veya kısmen iyonlaşmış halde bulunmalarının sonucu olarak, elektrostatik çekim kuvveti ile (COO−⋅⋅⋅⋅⋅⋅H3N+) oluşan bağlardır. Tuz Köprüsü

9
5) Apolar bağlar (hidrofob bağlar): Polipeptit zincirindeki amino asit kalıntılarının metil grubu, alifatik grup, siklik grup gibi apolar kısımlarının birbirlerine yeter derecede yakın olmaları halinde geçici bir polarite göstermelerinin sonucu ortaya çıkan ve Van der Waals-London çekme kuvveti diye bilinen zayıf çekme kuvveti ile (CH3⋅⋅⋅⋅⋅CH3) oluşan bağlardır. Hidrofobik etkileşimler gerçek bağ değildirler; elektron paylaşımı yoktur. Hidrofobik etkileşimler, proteinlerin iç kısımlarının kararlı olarak devamlılığının sağlanmasında rol oynar.

10
Proteinlerin yapısında itici güçler de bulunmaktadır: 1) Aynı yükü taşıyan gruplar arasında, iyonik güçlerin tersi olan, elektrostatik itme olur. 2) Çok yakın duran atomlar arasında Van der Waals itici güçleri vardır. Önemli not: Burada anlatılan bağlar daha çok Üçüncül yapıda gözlenen Kimyasal bağlar ya da çekmelerdir.!!!

11
Protein moleküllerinin yapısı ve konformasyonu
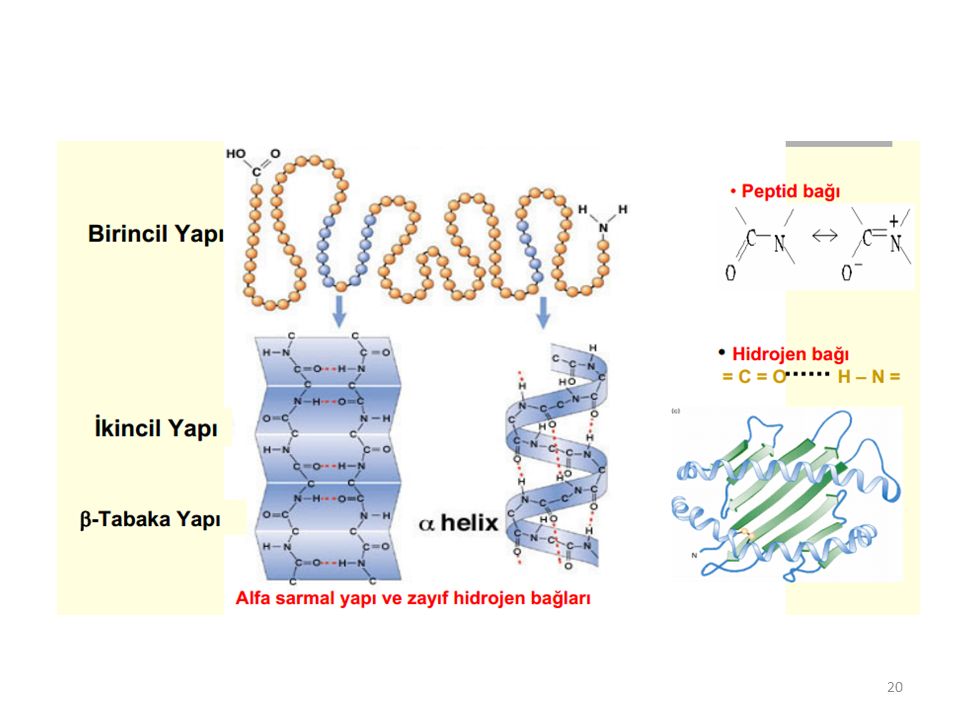
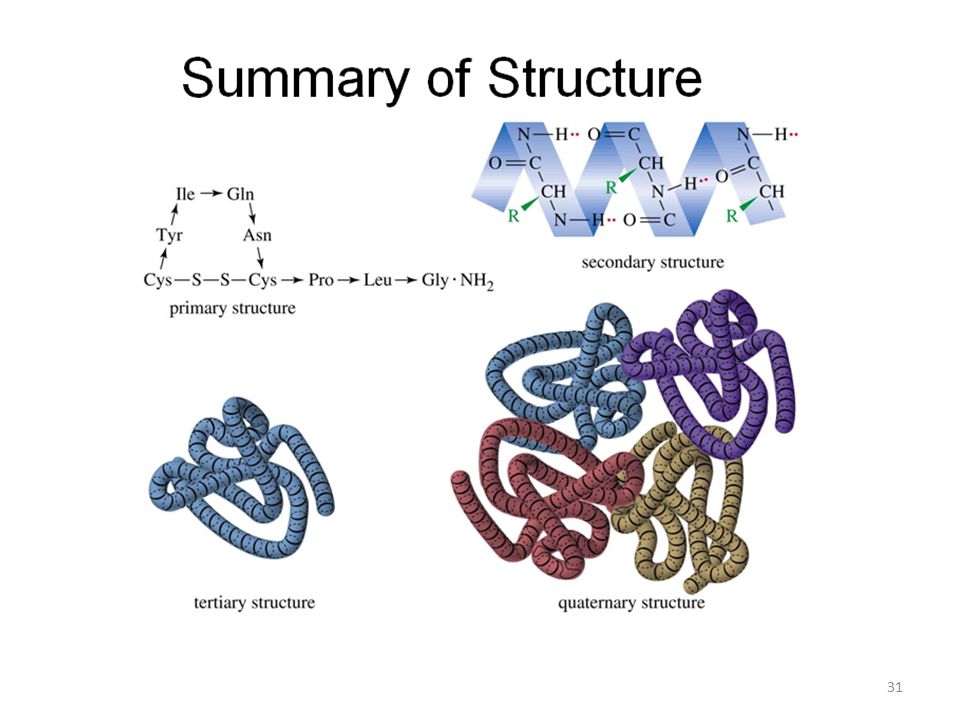
Proteinlerde birinci (primer), ikinci (sekonder), üçüncü (tersiyer) ve dördüncü (kuaterner) yapı diye dört yapı tanımlanır:
, ikinci (sekonder), üçüncü (tersiyer) ve dördüncü (kuaterner) yapı diye dört yapı tanımlanır:")
12
Bir proteinin primer (birinci) yapısı, bir protein için karakteristik ve genetik olarak tespit edilmiş olan amino asit dizilişidir; belirli türde, belirli sayıda, belirli diziliş sırasında amino asitlerin birbirlerine peptit bağlarıyla bağlanarak oluşturdukları bir polipeptit zinciri biçimindeki yapısıdır:
 yapısı, bir protein için karakteristik ve genetik olarak tespit edilmiş olan amino asit dizilişidir; belirli türde, belirli sayıda, belirli diziliş sırasında amino asitlerin birbirlerine peptit bağlarıyla bağlanarak oluşturdukları bir polipeptit zinciri biçimindeki yapısıdır:")
13
Bir proteinin sekonder (ikinci) yapısı
Bir proteinin sekonder (ikinci) yapısı, yarı sertleşmiş polipeptit zincirlerinin bükülmeler ve katlanmalarla oluşturdukları özgün kangallar biçimindeki yapısıdır. Bir proteinin sekonder yapısının oluşmasını ve sürdürülmesini sağlayan, primer yapı ile meydana gelen polipeptit omurgasının özelliği ve yan bağlardan özellikle hidrojen bağlarıdır. Ayrıca komşu R gruplarının sterik gerginliği ve aynı cins yük etkileşimini azaltmak hedefiyle yeterli uzaklığa yönlenme gereksinimidir. R- yan gruplarından uzanan özel kimyasal gruplar, özgün katlanmalar yönetirler.
 yapısı, yarı sertleşmiş polipeptit zincirlerinin bükülmeler ve katlanmalarla oluşturdukları özgün kangallar biçimindeki yapısıdır. Bir proteinin sekonder yapısının oluşmasını ve sürdürülmesini sağlayan, primer yapı ile meydana gelen polipeptit omurgasının özelliği ve yan bağlardan özellikle hidrojen bağlarıdır. Ayrıca komşu R gruplarının sterik gerginliği ve aynı cins yük etkileşimini azaltmak hedefiyle yeterli uzaklığa yönlenme gereksinimidir. R- yan gruplarından uzanan özel kimyasal gruplar, özgün katlanmalar yönetirler.")
14
Proteinler için: gelişigüzel kangallanım, α-heliks yapısı β-konformasyonu veya kırmalı tabaka yapısı olmak üzere üç değişik sekonder yapı tanımlanır. Proteinlerin gelişigüzel kangallanım tipi sekonder yapısında, polipeptit zincirin R- kalıntıları, α-karbonlar etrafında dönüşler yaparlar; fakat polipeptit zinciri boyunca tekrarlanmış bir örneğine rastlanmayacak biçimde davranışlar olur. Gelişigüzel kangallanımda hidrojen bağları ve diğer yan bağlar rol almaz; peptit bağları düzlemleri arasında kurucu bir ilişki yoktur.

15
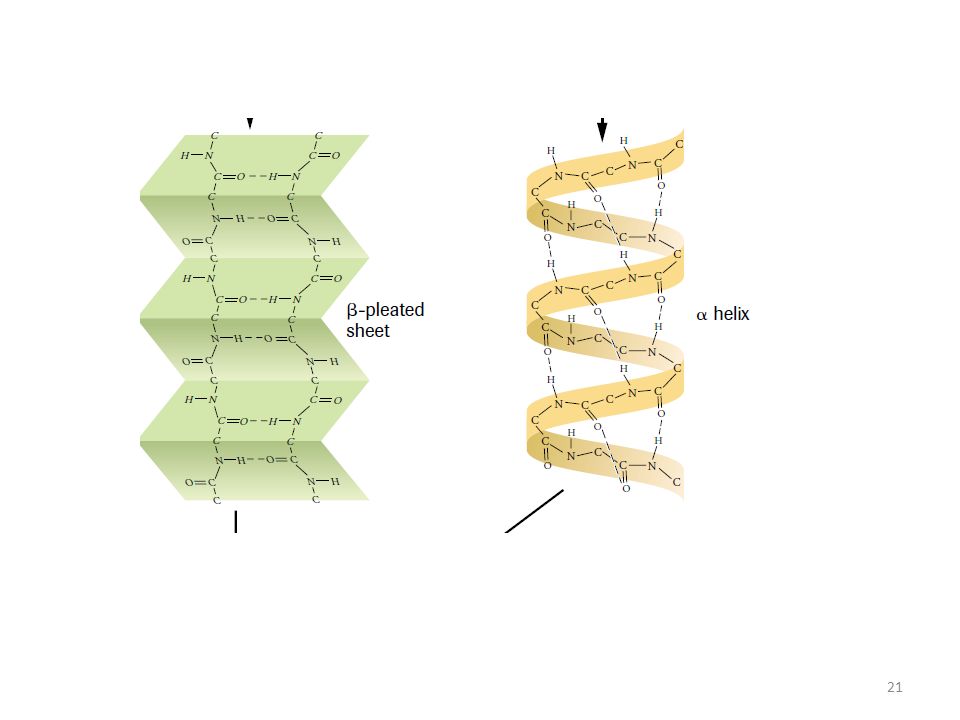
Proteinlerin α-heliks yapısı tipi sekonder yapısında, polipeptit omurgası, oluşması mümkün olan bütün hidrojen bağlarının oluşması için, kıvrımları sağa dönen bir heliks biçiminde bükülmüştür. Ve aminoasitlerin hepsi L-konfigürasyonunda olduğundan oluşan spiraller hep saat-ibresi yönündedir (right-handed) Proteinlerin α-heliks yapısı tipi sekonder yapısında, α-heliksin her kıvrımında 3,6 amino asit ünitesi bulunur ve bir kıvrımın yüksekliği 0,56 nm kadardır; (yani tekrarlanma mesafesi) polipeptit zincirdeki amino asit kalıntılarının R- grupları, heliks yüzeyinden dışarı sarkmışlardır.
 Proteinlerin α-heliks yapısı tipi sekonder yapısında, α-heliksin her kıvrımında 3,6 amino asit ünitesi bulunur ve bir kıvrımın yüksekliği 0,56 nm kadardır; (yani tekrarlanma mesafesi) polipeptit zincirdeki amino asit kalıntılarının R- grupları, heliks yüzeyinden dışarı sarkmışlardır.")
16
Deri, tırnak, boynuz, saç, tüy… yapılarında α-heliksler bulunmaktadır.
Yün ve kas proteinleri de α-heliks yapısındadır ve uzayabilirler. Yün esneyebilir ve yumuşaktır. Deterjan ve ısı ile muamele edilirse α-bağlantıları kopar ve yerine β-köprüleri oluşur. Böylelikle de kırlır ve sertleşir. Ancak ipek ağırlıklı β-pleated sheet-düzgün katmanlar yapısındadır ve bu nedenle gerilemez. Halkalı yapısı nedeniyle pirolin nadiren α- heliks yapısında görülür. Glisin ise genellikle rastgele sarmal oluşturma eğilimine sahip olduğundan sıklıkla α-heliks yapısına katılmaz.

17
Genel olarak bir alfa heliksin kararlılığını etkileyen faktörleri şu şekilde verebiliriz.
Yüklü R gruplarına sahip amino asit kalıntıları arasındaki elektrostatik itme (veya çekme); Birbirine yakın R gruplarının boyutu; 3 veya 4 amino asit uzaklıktaki R gruplarının etkileşimi; Pro veya Gly amino asitlerinin varlığı; Alfa heliks üzerinde oluşan elektrik dipollerin dağılımı
; Birbirine yakın R gruplarının boyutu; 3 veya 4 amino asit uzaklıktaki R gruplarının etkileşimi; Pro veya Gly amino asitlerinin varlığı; Alfa heliks üzerinde oluşan elektrik dipollerin dağılımı.")
18
Proteinlerin β-konformasyonu veya kırmalı tabaka yapısı tipi sekonder yapısında, molekülün şekli, kırmalı tabakalı görünümdedir. Proteinlerin kırmalı tabaka yapısı tipi sekonder yapısında, hidrojen bağları önemli rol almıştır. α-Yapısına göre daha gevşek bir yapıdır.

19
Proteinlerin kırmalı tabaka yapısı tipi sekonder yapısında, polipeptit zincirdeki amino asit ünitelerinin R- grupları, ya aynı yönde birbirlerine yaklaşabiliyorsa paralel dizilme (paralel β-pleated sheet) gösterirler, ya da zıt yönlerde sıralanma ile antiparalel dizilme (antiparalel β-pleated sheet) gösterirler. Yaklaşan R guplarının küçük olması gerekir.

22
Bir proteinde hem α- hem de β- yapı segmentleri bulunabilir.
Örnek: kollajen Kollajen hareket sisteminin yapı taşlarını, özellikle kemik, kıkırdak, lif ve eklemleri oluşturan protein. Bu protein birbiri üzerine sarılmış üç alfa zincirinden meydana gelir.Bu üç segment birbirine β-bağlantılarıyla bağlanmıştır.

23
Zincirler arası bağlantılar H-köprüleri yerine S-S disülfid köprüleri ile de bağlanabilir. Buna en güzel örnek kıvırcık ve düz saçlardır: Saçın yapısı ağırlıklı keratin den oluşur. Keratin içindeki ağırlıklı aminoasit de sisteindir. Ve bunlar disülfid köprüler,i ile sistini oluştururlar. Kıvırcık saçta köprüler zincir içerisinde, düz saçlarda zincirler arasındadır. düz saç kıvırcık saç

24
İndirgeme ile disülfit köprüleri kopar, ve sisteinlere dönüşür, değişik boyutlarda bigudilerle sarılır, sarilan saç oksitlenir ve bigudilerin yönlendirmesine uygun olarak yeni disülfid köprüleri oluşur.

25
Bir proteinin tersiyer (üçüncü) yapısı
Bir proteinin tersiyer (üçüncü) yapısı, polipeptit zincirinin, sekonder yapı oluşumundan sonra, daha önce açıklanan bağlayıcı güçlerin hepsinin toplamı ile uzayda daha ileri katlanmalar, kıvrılmalar yapar ve tersiyer yapı oluşur.
 yapısı, polipeptit zincirinin, sekonder yapı oluşumundan sonra, daha önce açıklanan bağlayıcı güçlerin hepsinin toplamı ile uzayda daha ileri katlanmalar, kıvrılmalar yapar ve tersiyer yapı oluşur.")
26
Hatırlatma 1 : küçük ve nötr R grupları zincirlerarası H köprülerine izin verir (Glisin, val, ala gibi)yani beta yapısı olasılığı doğar. Ancak R grupları hacimli ve H köprüsü oluşturamayacak gruplar da olabilir (Pirolin gibi) pirolin (peptid köprüsü yaptığında H üzerinde H kalmaz. Bu nedenle H köprüsü kuramaz.!! Bu nedenle içerisinde daha çok pirolin içeren bir peptid α-heliks yapısını tercih eder.

27
Hatırlatma 2: Yan zincirlerde yük olduğunda: Örnek lisin ve arginin
Yüklerden dolayı itme çekmeler proteinlerin tersiyer yapılarını etkiler. Bu iki AA fizyolojik pH da + yüklü yan zincirler içerirler. Dizilişte birbirlerine yakın iseler, bu durumda birbirlerini itecek ve α-heliks yapısına engel olacaklardır. Glutamik ve aspartik asit de fizyolojik pH da - yüklü yan zincirler içerirler. Dizilişte birbirlerine yakın iseler, bu durumda birbirlerini itecek ve α-heliks yapısına engel olacaklardır. Fakat zıt yüklüler yakın iseler (lisin ve aspartik asit gibi) birbirlerini çekecekler ve H köprülerinden daha güçlü etkileşimler oluşturacaklardır. Böylelikle tersiyer yapının şekli ortaya çıkacaktır.
 birbirlerini çekecekler ve H köprülerinden daha güçlü etkileşimler oluşturacaklardır. Böylelikle tersiyer yapının şekli ortaya çıkacaktır.")
28
Küresel bir proteinde tüm bu etkiler sonucunda hidrofobik gruplar içe yönelirler ve VanDer Waals etkileşimleriyle kararlılık kazanırlar. Polar /yüklü gruplar ise küresel proteinin dışında (yüzeyinde) kalıp bir sıvı sistem olarak kabul gören canlı sisteminde faaliyet gösterirler. Örnek: hemoglobin: küresel bir konjuge proteindir. İçi hidrofobdur. Dışı ise hidrofil olduğundan sıvı kan sisteminde rahatlıkla hareket eder. Bir protein ömrünü ancak sınırlı bir pH aralığında ve belli bir çözücüde koruyabilir. Örneğin böbrek yetmezliğinde kanda üre miktarı artar. Yani ortamın bazikliği artar. Böylelikle kandaki pH değişir ve kan dolaşımı nedeniyle tüm vücut hücrelerinin pH ı değişir. Ve de sonuç proteinlerin yıkımı yani ölüm olur.

29
Bir proteinin kuaterner (dördüncül) yapısı
Bazı proteinler iki yada daha fazla alt üniteye – protein kökenli olmayan “prostetik gruplar”a bağlıdırlar. Prostetik gruplar içeren bu tür proteinlere KONJUGE PROTEİNLER adı verilir. Bu alt ünitelerin tersiyer yapılarının birbirleri ile etkileşmesi ile kuaterner (dördüncül) yapılar oluşur. Bazı önemli konjuge proteinler: HEMOGLOBİN = Hem (prostetik grup) + Globülin (Protein) ENZİMLER = Vitamin ( Koenzim prostetik grubu) protein NÜKLEOPROTEİNLER =Nükleik asit (Polimerik yapıda prostetik grup) + Protein Nükleoproteinler genetik özelliklerin korunması ve sonraki nesillere aktarılmasını sağlarlar) GLUKOPROTEİNLER = Karbonhidrat (prostetik grup) Protein Glukoproteinler kan gruplarının tayininde kullanılırlar:
 yapılar oluşur. Bazı önemli konjuge proteinler: HEMOGLOBİN = Hem (prostetik grup) + Globülin (Protein) ENZİMLER = Vitamin ( Koenzim prostetik grubu) + protein. NÜKLEOPROTEİNLER =Nükleik asit (Polimerik yapıda prostetik grup) + Protein. Nükleoproteinler genetik özelliklerin korunması ve sonraki nesillere aktarılmasını sağlarlar) GLUKOPROTEİNLER = Karbonhidrat (prostetik grup) + Protein. Glukoproteinler kan gruplarının tayininde kullanılırlar:")
32
HEMOGLOBİN : Hemoglobin, kanda solunum organından dokulara oksijen, dokulardan solunum organına ise karbondioksit ve proton taşıyan protein (reversible işlem yapar) . Ve de kana rengini veren pigmenttir. Eritrositlerin (alyuvar-kırmızı kan hücresi) içerisinde bulunur. Akciğerden aldığı Oksijeni +2 değerlikli demir içeren HEM molekülleri ile bağlar ve hücrelere taşır. Reversible oksijen alışverişi demir-2 ve demir-3 oksidasyon-reduksiyon reaksiyonunun sürekliliği ile sağlanır. Bu sürekliliği +2 demir içeren HEM molekülü sağlar. Hemoglobinin 3-boyutlu yapısı. Dört altbirim kırmızı, sarı olarak ve hem grubu yeşil olarak gosterilmiştir.

33
Başlıca sentez yeri eritrosit üretimi sırasında kemik iliğidir.
Hemoglobinde her birinin özel tersiyer yapısı olan dört adet polipeptid zinciri vardır. İkisi alfa, ikisi de beta- sekonder yapılarında olan bu 4 adet peptid zinciri hem birbirlerine bağlanmakta, hem de 4 adet HEM molekülüne bağlanmaktadırlar. HEM ler porpirin türevi olup elipsoid yapılardadırlar. Porpirin halkası tamamen konjugedir ve planar yapıdadır. HEM grubu

34
Hemoglobindeki Fe2+'in koordinasyon sayısı 6 olup bu koordinasyon yerlerinden dördüne pirol halkasının azotu, beşincisine globin molekülünün histidininin imidazol grubunun azotundan bağlanır. Ve de boş olan altıncı koordinasyon yeri oksijenin tutunup bırakılmasında rol oynar. Ve de reversible reaksiyonda CO tutar. HATIRLATMA: sigara tiryakilerinde tütün dumanındaki CO akciğerlerde temizlenmek üzere HEM lere bağlanır. HEM lerin yeterli oksijen tutmasını engeller böylelikle. Tiryakilerin Hemoglobinlerinin yaklaşık %20 si körlenmiş durumdadır bu nedenle. Bu durum da kansızlığa ve onun neden olduğu diğer hasarlara yol açar.

35
Küresel yapıdaki bir adet hemoglobinde 4 adet HEM protein zincirlerinin oluşturduğu yuvalara girmişlerdir. HEM lerin hidrofobik ( Vinil ve metil grupları) grupları etraflarındaki polipeptidlerin hidrofobik yan gruplarıyla etkileşir. HEM lerin hidrofilik propanoik asit yan zincirleri ise etraflarındaki polipeptidlerin zıt yüklü yan grupları ile ( lisin ve arginin gibi fizyolojik pH da + yüklü gruplar içerenlerle) etkileşirler. Bu etkileşimler sonucunda hemoglabindeki HEM lerin etrafında hidrofobik bir ortam oluşur. Yani hemogloinin içi HİDROFOB, dışı ise kan sisteminde olduğundan zaten HİDROFİL dir. Hemoglobinin içindeki hidrofob ortam iki yönlü demir-2 ve demir-3 oksidasyon-reduksiyon reaksiyonuna olanak sağlar. Çünkü ortam polar olsa idi sadece Fe(2) den Fe(3)e dönüşüm olurdu ve oksijenin sürekli alınıp verilmesi sağlanamazdı.
 den Fe(3)e dönüşüm olurdu ve oksijenin sürekli alınıp verilmesi sağlanamazdı.")
36
ENZİMLER: Yaşam için gerekli olan kimyasal işlemleri katalizlerler
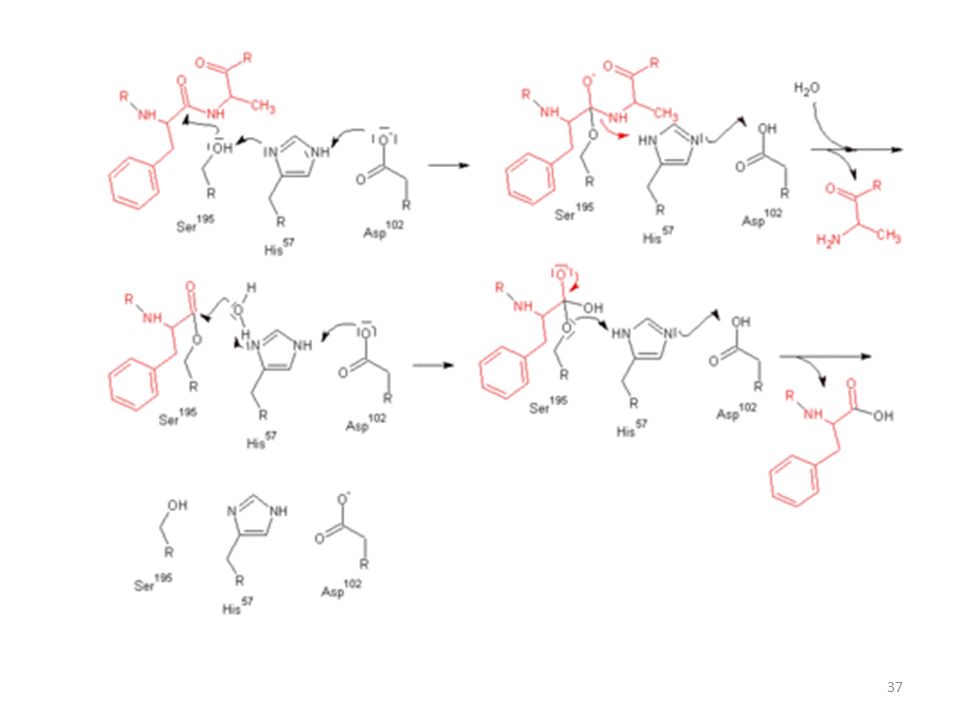
ENZİMLER: Yaşam için gerekli olan kimyasal işlemleri katalizlerler. Vücutta miktarları az, ancak işlevleri çok önemlidir. Binlerce enzim vardır, ancak yapısı tamamen bilinen enzim sayısı çok azdır. Yapısı bilinenlerden örnek olarak pankreas tarafından salgılanan ve gıdalardan alınan proteinleri küçük peptidlere bölen bir sindirim sistemi enzimi CHYMOTRYPSIN verilebilir. CHYMOTRYPSIN Disülfid köprüleri ile birbirine bağlanan üç adet polipeptid zincirinden oluşmuştur. 57 numaralı AA Histidin, 102 numaralı AA Aspartik asit ve 195 numaralı AA SERİN dir. Bunlar yapının aktif ucunu oluştururlar. Ve bu aktif uç proteini belli yerlerinden hidroliz ile parçalar ve böylelikle sindirim gerçekleşir. En sonunda protein bağırsaktan emilebilecek küçüklükteki parçalara ayrılır. Bu reaksiyonlar canlıda pH 6-7 lerde saniyelerden daha az zamanda olmaktadır.

38
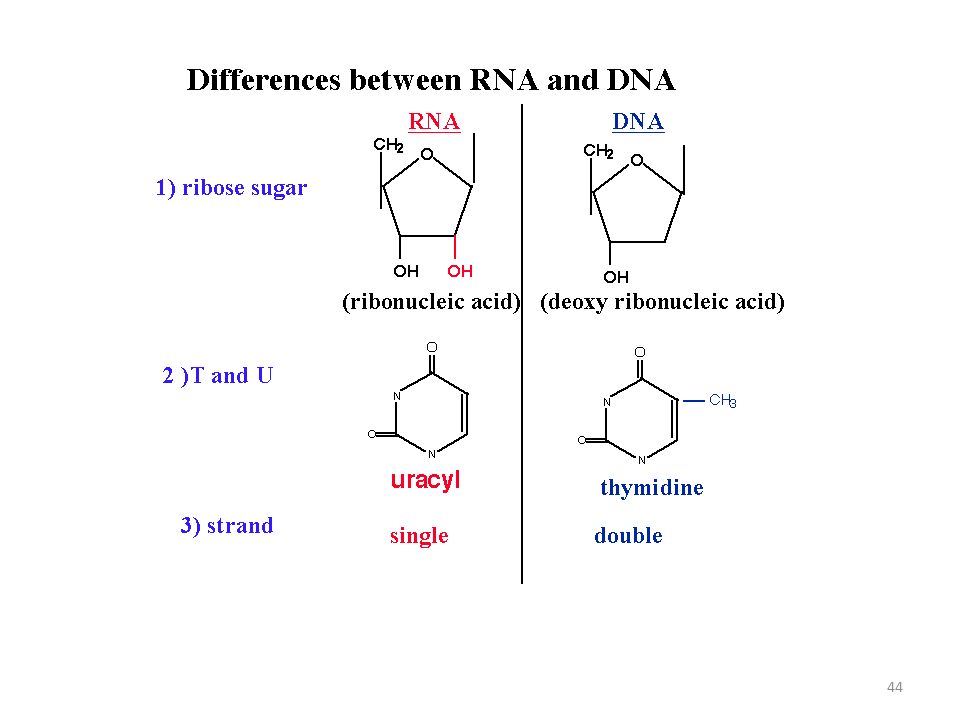
NÜKLEOPROTEİNLER: Her canlı hücrede proteinlerin nükleik asit denilen ve biyolojik polimerlerle birleşmesinden oluşan Nükleoproteinler bulunur. Bunlar prostetik grubu nükleik asit olan konjuge proteinlerdir. Nükleoproteinler genetik özelliklerin korunması ve sonraki nesillere aktarılmasını sağlarlar. Nükleik asit polimerinin monomerine NÜKLEOTİD denir. (monomer nükleotidlerin polimerleşmesi sonucunda nükleik asit oluşur) Bu polimerleri oluşturan nükleotid birimlerin her biri üç bölümden oluşur: 1) Azotlu heterosiklik bir baz (PÜRİNLER (Çift halkalı) : Adenin (A) , Guanin (G) – DNA ve RNA’da bulunur; PRİMİDİNLER (Tek halkalı ) : Sitozin (S) – DNA ve RNA’da bulunur; Timin (T) Yalnız DNA’da ve Urasil (U) yalnız RNA’da bulunur) 2) beş karbonlu (pentoz) bir şeker (monosakkarit olan “riboz”) 3) bir fosfat grubu.
 Bu polimerleri oluşturan nükleotid birimlerin her biri üç bölümden oluşur: 1) Azotlu heterosiklik bir baz (PÜRİNLER (Çift halkalı) : Adenin (A) , Guanin (G) – DNA ve RNA’da bulunur; PRİMİDİNLER (Tek halkalı ) : Sitozin (S) – DNA ve RNA’da bulunur; Timin (T) Yalnız DNA’da ve Urasil (U) yalnız RNA’da bulunur) 2) beş karbonlu (pentoz) bir şeker (monosakkarit olan riboz ) 3) bir fosfat grubu.")
39
RNA'da bulunan şeker “riboz” , DNA'da bulunan şeker ise “deoksiribozdur”. (RNA’da pentoz halkasının 2′ konumunda bir hidroksil grubu yoktur, RNA’da ise pentoz halkasının iki hidroksil grubu vardır. RNA’da fazladan bulunan hidroksil grupları, hidroliz nedeniyle onun DNA’dan daha az dayanıklı olmasına neden olur)
.")
40
DNA ve RNA içerdikleri azotlu bazlarda da farklılık gösterirler: adenin, guanin ve sitozin her ikisinde, timin yalnızca DNA'da, urasil ise yalnızca RNA'da bulunur.

41
RNA çok kıvrımlı ancak tek zincirli yapıya sahiptir
RNA çok kıvrımlı ancak tek zincirli yapıya sahiptir. DNA birbirini sarmalamış iki heliks şeklindedir.

43
RNA'yı oluşturan kimyasal gruplar
RNA'yı oluşturan kimyasal gruplar. P, fosfat; Z, riboz şeker; A, C, G, U, sırasıyla adenin, sitozin, guanin ve urasil. Zincirin doğrultusu şekerlerin 5' ve 3' karbonlarının sırası tarafından belirlenir. İçerideki hidrofobik bazlardan adenin-timin ve guanin-sitozin ikilileri arasında H köprüleri oluşur. Bu durum DNAya şeklini verir. Dış etkenler (radyasyon, kimyasallar..) bu dizilimi bozarak genetik hastalıklara yol açar.
 bu dizilimi bozarak genetik hastalıklara yol açar.")
Benzer bir sunumlar












>")
 Elementler :>")


>")




