Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Kalıtımın Moleküler Temeli-Genden Proteine..
FFMBG101- Genel Biyoloji Kalıtımın Moleküler Temeli-Genden Proteine.. 2-6 Aralık 2013

2
Yaşamı Yönetim ve işletme kılavuzu 1953, James Watson ve Francis Crick
Deoksiribonükleotit yapısına ait İkili sarmal modeli Şekil 1

3
DNA, kalıtım materyali Kalıtım bilgisi DNA programı
DNA’nın kimyasal dilinde kodlanır ve tüm hücrelerde üretilir DNA programı Pekçok farklı tipte özelliklerin gelişimini yönetir

4
Genetik Materyal Araştırması: Bilimsel Araştırma
Kalıtımda DNA’nın rolü İlk çalışmalar bakteriler ve onları enfekte eden virüsler üzerinde yapıldı DNA’nın Bakterileri transforme edilebileceğinin (dönüştürebileceğinin) kanıtı Frederick Griffith_Streptococcus pneumoniae Memelilerde zatürreye neden olan bakteri Bakterinin iki suşu ile çalıştı Patojenik (virülent) suş ve patojenik olmayan (avirülent) suş
 kanıtı. Frederick Griffith_Streptococcus pneumoniae. Memelilerde zatürreye neden olan bakteri. Bakterinin iki suşu ile çalıştı. Patojenik (virülent) suş ve patojenik olmayan (avirülent) suş.")
5
Griffith Patojenik olmayan suşun canlı hücreleri; transformasyonla bunlardan bazıları patojenik özellik kazandı CanlıS hücreleri (kontrol) virülent Canlı R hücreleri (kontrol) avirülent Isı ile öldürülmüş S hücreleri Isı ile öldürülmüşS hücreleri ve canlı R hücrelerii karışımı Fare ölür Kan örneklerinde canlı S hücreleri bulundu Fare sağlıklı Mouse ölür SONUÇLAR Griffith bu olayı transformasyon olarak adlandırdı Yabancı DNA’nın bir hücre tarafından asimilasyonundan dolayı genotipte ve fenotipte meydana gelen bir değişme Şekil 2
 virülent. Canlı R hücreleri. (kontrol) avirülent. Isı ile öldürülmüş. S hücreleri. Isı ile öldürülmüşS hücreleri. ve canlı R hücrelerii karışımı. Fare ölür. Kan örneklerinde. canlı S hücreleri. bulundu. Fare sağlıklı. Mouse ölür. SONUÇLAR. Griffith bu olayı transformasyon. olarak adlandırdı. Yabancı DNA’nın bir hücre tarafından. asimilasyonundan dolayı genotipte. ve fenotipte meydana gelen bir. değişme. Şekil 2.")
6
Viral DNA’nın hücreleri programlayabileceğine dair kanıt
DNA’nın Genetik materyal olduğuna dair diğer kanıt Bakterileri enfekte eden bir virüsle çalışılarak elde edildi Bakterileri enfekte eden virüsler, bakteriyofajlar Moleküler genetik çalışmalarında araştırıcıların yaygın olarak kullandığı bir materyal Şekil 3 Faj başı Kuyruk Kuyruk fiberi DNA Bakteri hücresi 100 nm

7
The Hershey ve Chase deneyi, 1952
Bakteri hücrelerini enfekte eden T2 fajının DNA ve proteinini işaretlemek için radyoaktif fosfor ve sülfürü kullandılar Faj proteinleri enfeksiyon sırasında bakteri hücresinin dışında kalırken faj DNA’sı hücre içerisine girer. Kültürü yapıldığında radyoaktif faj DNA sı bulunan bakteri hücreleri radyoaktif fosfor içeren bazı yeni fajları dışarı salar. Sonuç: Çıkarım: T2 fajının genetik materyali protein değil DNA’dır Radyoaktif işaretli fajları bakterilerle karıtşırdı. Fajlar bakteri hücrelerini enfekte etti Bakterinin dışındaki fajları ayırmak için belnderla karıştırıldı Pelet ve sıvıda radyoaktivite ölçüldü Kültür1: Fajlar faj proteinine bağlanan radyoaktif sülfür (pembe renkli) lü ortamda üretildi Santrifüj edildi, bakteriler Tüpün dibinde pellet oluşturdular Kültür 2: Fajlar faj DNA sına entegre olan radyoaktif fosfor lu (mavi) ortamda üretildi Şekil 4
 lü ortamda üretildi. Santrifüj edildi, bakteriler. Tüpün dibinde pellet. oluşturdular. Kültür 2: Fajlar faj DNA sına entegre. olan radyoaktif fosfor lu (mavi) ortamda üretildi. Şekil 4.")
8
DNA’nın genetik materyal olduğunun diğer kanıtı
1950 lerin öncesinde, DNA zaten bilinmekteydi DNA nükleotit polimeridir ve her bir nükleotit üç birimden meydana gelir: bir nitrojen baz, bir şeker ve bir fosfat grubu Şeker-fosfat iskeleti Nitrojen bazı 5 end O– O P CH2 5 4 H 3 1 CH3 N Timin (T) Adenin (A) Sitozin (C) OH 2 Şeker (deoksiriboz) 3 ucu Fosfat Guanin (G) DNA nükleotiti Şekil 5
 Adenin (A) Sitozin (C) OH. 2 Şeker (deoksiriboz) 3 ucu. Fosfat. Guanin (G) DNA nükleotiti. Şekil 5.")
9
Erwin Chargaff farklı organizmalardan DNA nın baz kompozisyonunu analiz etti
1947, Chargaff kuralları DNA kompozisyonu bir türden diğerine değişir A=T, G=C Bu türler arasındaki moleküler çeşitliliğin kanıtıdır Bu bulgu genetik materyal olarak DNA yı daha güvenilir bir aday yaptı

10
DNA’nın yapısal modelinin oluşturulması:Bilimsel araştırma
DNA’nın genetik materyal olduğuna dair biyologlar ikna oldu. Ancak; DNA’nın kalıtımdaki rolü üzerinde yapısal özellikleri nasıl bir etki gösterir?

11
Maurice Wilkins ve Rosalind Franklin
DNA nın moleküler yapısını çalışmak için X ışının kristalografisi adlı tekniği kullandılar Rosalind Franklin Bu tekniği kullanarak DNA molekül yapısınının resmini ortaya çıkardı (a) Rosalind Franklin Franklin’s X-ray diffraction Photograph of DNA (b) Şekil 5 a, b İkili sarmal yapı 2 nm lik genişlik Bazlar arasında 0.34 nm lik mesafe
 Rosalind Franklin. Franklin’s X-ray diffraction. Photograph of DNA. (b) Şekil 5 a, b. İkili sarmal yapı. 2 nm lik genişlik. Bazlar arasında 0.34 nm lik mesafe.")
12
(a) DNA yapısının anahtar özellikleri (c) Uzay doldurma modeli
Watson ve Crick; DNA’nın ikili sarmal yapıda olduğu sonucuna vardı X-ışını kristalografik görüntülerinden elde edilen veriler ışığında Şekil 6 a, c C T A G 0.34 nm 3.4 nm (a) DNA yapısının anahtar özellikleri 1 nm (c) Uzay doldurma modeli
 DNA yapısının anahtar özellikleri. 1 nm. (c) Uzay doldurma modeli.")
13
Franklin DNA’nın Nitrojen bazlar
İki antiparalel şeker-fosfat iskeletinden meydana geldiğini ve nitrojen bazlarının molekülün iç kısımda yer aldığını belirtti Nitrojen bazlar Spesifik kombinasyonlarda eşleşmektedirler: adenin ile timin, ve sitozin ile guanin

14
(b) Kısmi kimyasal yapı
O –O OH H2C T A C G CH2 O– 5 ucu Hidrojen bağı 3 ucu P (b) Kısmi kimyasal yapı Şekil 7b
 Kısmi kimyasal yapı. Şekil 7b.")
15
Herbir baz çifti farklı sayıda hidrojen bağı oluşturur
Watson ve Crick eşlemede ek bir özgünlük olması gerektiğini ortaya koydular Baz yapısı ile alakalı Herbir baz çifti farklı sayıda hidrojen bağı oluşturur Adenin ve timin iki H bağı; sitozin ve guanin 3 H bağı oluşturur Bazlar iç kısımda, fosfat grupları dış tarafta Yarı korunumlu (semi konservatif) DNA replikasyon modeli
 DNA. replikasyon modeli.")
16
N H O CH3 şeker Şeker Adenin (A) Timin (T) Guanin (G) Sitozin (C) Şekil 8
 Timin (T) Guanin (G) Sitozin (C) Şekil 8")
17
Temel Prensip: Kalıp zincire baz eşleşmesi
DNA nın iki zinciri komplementer (birbirinin tamamlayıcısı) olduğundan Replikasyonda her bir zincir yeni zinciri oluşturmak üzere kalıp olarak görev yapar
 olduğundan. Replikasyonda her bir zincir yeni zinciri oluşturmak üzere kalıp olarak görev yapar.")
18
faj (a) ve ondan izole edilen DNA (b)
Virüs ve Bakteri kromozomları nispeten basit DNA molekülleridir faj (a) ve ondan izole edilen DNA (b)
 ve ondan izole edilen DNA (b)")
19
Table 12-1 The Genetic Material of Representative Viruses and Bacteria

20
Supercoiling virüs vebakteri Kromozomlarının sıkışmasını kolaylaştırır
T2 faj DNA sı Supercoiling virüs vebakteri Kromozomlarının sıkışmasını kolaylaştırır Figure 12-2 Electron micrograph of bacteriophage T2, which has had its DNA released by osmotic shock. The chromosome is 52m long. Figure 12.2

21
Figure 12-4 Depictions of the transformations leading to the supercoiling of circular DNA. L equals the linking number. Figure 12.4

22
DNA Fun Fact İnsan hücresindeki DNA nın uzunluğu ~ 2 metres İnsan vücudundaki hücre sayısı ~1014 İnsan vücudundaki DNA’nıntoplam uzunluğu ~ 2 x 1014 metre = 2 x 1011 km Dünyanın Güneşe uzaklığı ~ 1.5 x 108 km Dünya ve güneş arasında 1000 kez gidip gelecek uzunlukta DNA ya sahipsiniz!

23
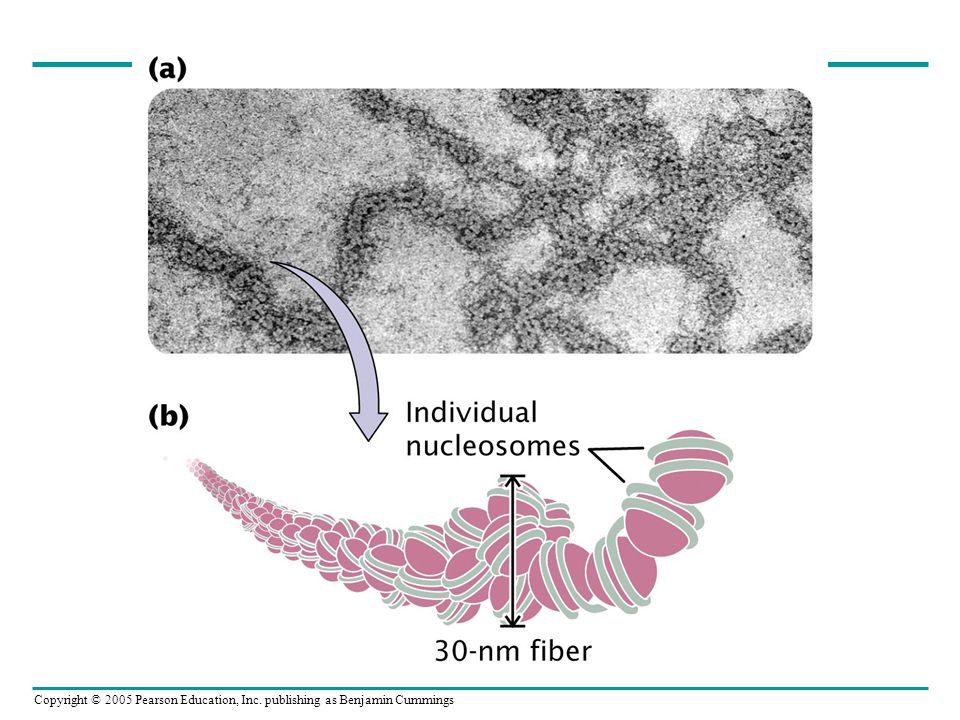
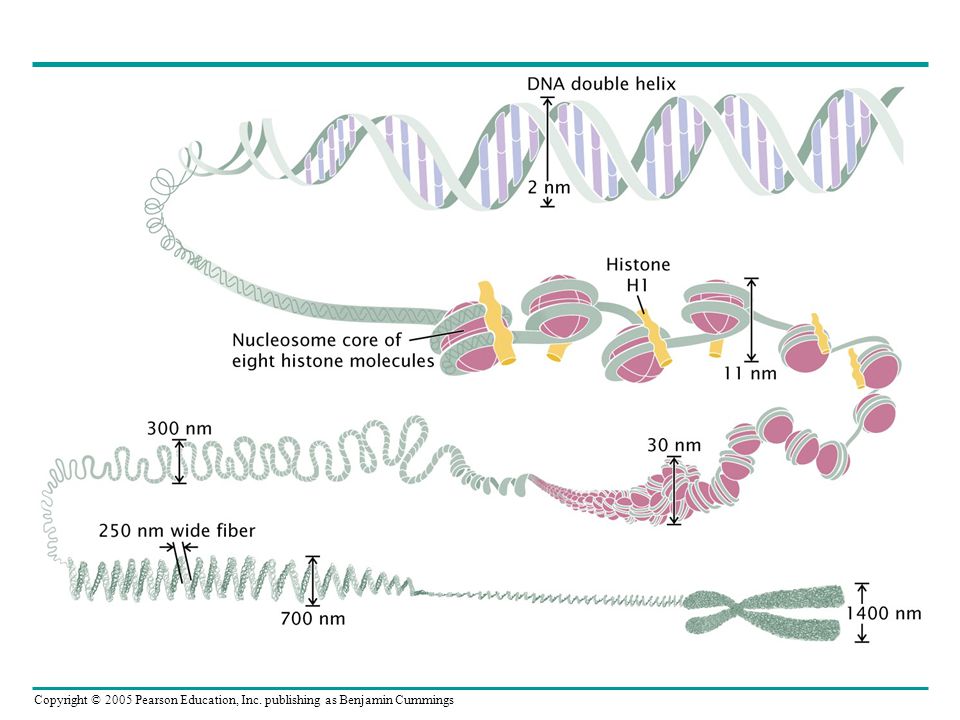
Ökaryotlarda DNA kromatinler içerisinde organize olurlar
Kromatin yapısı ve Nükleozomlar Table Categories and Properties of Histone Proteins Table 12.2

24
Figure 12-8 (a) Dark-field electron micrograph of nucleosomes present in chromatin derived from a chicken erythrocyte nucleus. (b) Dark-field electron micrograph of nucleosomes produced by micrococcal nuclease digestion. Figure 12.8

25
~200 bp digest linker DNA 147 bp Nükleozomların core histonları
Nükleaz ~200 bp Nükleaz 147 bp digest linker DNA

28
Heterochromatin-Kromozom bantları mitotik kromozom boyunca bölgeleri farklılaştırır
İnsan mitotik kromozomu G-banded karyotype of a normal human male. Chromosomes were derived from cells in metaphase.

29
Sentromerik DNA dizileri
Ökaryot kromozomu Tekrarlı DNA larla karakterize edilen kompleks sekans organizasyonu gösterirler Satellit DNA Sentromerik DNA dizileri

30
Figure Separation of main-band (MB) and satellite (S) DNA from the mouse by using ultracentrifugation in a CsCl gradient. Figure 12.15

31
DNA REPLİKASYONU DNA replikasyonunda
Atasal molekül ayrılır, baz eşleşme kurallarına göre yeni zincirler oluşturulur Ana DNA molekülü iki komplementer zincirden oluşmaktadır. Her baz özgün partneri ile H bağı yaparak Eşleşir: A ve T; G ve C Replikasyondaki ilk basamak İki DNA zincirinin ayrılması Her bir parental zincir yeni komplementer zinciri boyunca Nükleotiterin sırasını belirlemek Üzere kalıp olarak kullanılır (d) Yeni zincirin Şeker-fosfat iskeletini Oluşturmak için bazlar Birbirlerine bağlanırlar. Her yeni oluşan DNA Bir parental zincir ve bir yeni zincirden meydana gelmektedir A C T G Şekil 9 a–d
 Yeni zincirin. Şeker-fosfat iskeletini. Oluşturmak için bazlar. Birbirlerine bağlanırlar. Her yeni oluşan DNA. Bir parental zincir ve bir yeni. zincirden meydana gelmektedir. A. C. T. G. Şekil 9 a–d.")
32
DNA replikasyonu semikonservatiftir
Yeni oluşan iki molekülden biri eski zinciri ve yeni sentezlenen zinciri içermektedir Şekil 10 Konservatif Model. İki parental zincir yeni oluşacak zincirler için kalıp olarak görev görür ve parental ikili sarmalı muhafaza eder Semikonservatif Model. Parental molekülün zincirleri ayrılır, Ve her biri yeni komplementer zincirin sentezi için Kalıp olarak görev yapar. Dağıtılmış Model. Her iki yeni zincir eski ve yeni sentezlenmiş DNA karışımından meydana gelir Parental hücre İlk replikasyon İkinci replikasyon (a) (b) (c)
 (b) (c)")
33
Meselson ve Stahl deneyleri
Yarıkorunumlu DNA replikasyon modelini destekledi Şekil 11 Her iki araştırıcı E. coli bakterisini 15N, ağır nitrojen izotopu işaretli nükleotid öncüllerini içeren ortamda uzun süre ürettiler. Dolayısıyla üreyen bakterinin DNA sı bu işaretli nükleotitleri yapısına kattı. Bilim insanları bu kez bakteriyi sadece 14N içeren (daha hafif ve yaygın N izotopu) besiyerine ekip orada ürettiler. Yeni oluşan DNA ‘ların yoğunşuklarını ölçerek DNA replikasyon modelini aydınlattılar. DENEY Bu iki santrifüj tüplerindeki bantlar biri 20 diğeri ise 40 dakia sonra aynı erlenden alınan DNA örnekler in santrifüjü sonrası sonuçları göstermektedir SONUÇLAR Bakteri 15N içeren besi- yerinde üretildi Bakteriler 14N içeren besiyerine aktarıldı 2 1 DNA örenekleri 20 dak. Sonra santrifüj edildi (ilk replikasyon sonrası) 3 DNA örnekleri 40 dk. sonra santrifüj edildi (2. replikasyon Sonrası) 4 Az yoğun Daha yoğun
 besiyerine ekip orada ürettiler. Yeni oluşan DNA ‘ların yoğunşuklarını ölçerek DNA replikasyon modelini aydınlattılar. DENEY. Bu iki santrifüj tüplerindeki bantlar biri 20 diğeri ise 40 dakia sonra aynı erlenden alınan DNA örnekler in. santrifüjü sonrası sonuçları göstermektedir. SONUÇLAR. Bakteri 15N. içeren besi- yerinde üretildi. Bakteriler. 14N içeren besiyerine. aktarıldı DNA örenekleri. 20 dak. Sonra santrifüj edildi. (ilk replikasyon sonrası) 3. DNA örnekleri. 40 dk. sonra. santrifüj edildi. (2. replikasyon. Sonrası) 4. Az. yoğun. Daha yoğun.")
34
ÇIKARIM Meselson ve Stahl’ın bulguları: yarıkorunumlu DNA replikasyon modeli. 14N besiyerindeki ilk replikasyonda hibrit bir DNA bandı oluştu (15N–14N). Bu sonuç konservatif modelini çürüttü. İkinci replikasyonda hem hafif hem de hibrit DNA üretildi, dispersif modeli çürüttü, semikonservtif modeli destekledi İlk replikasyon İkinci replikasyon Konsevatif model Yarıkorunumlu Dispersif model
. Bu sonuç konservatif modelini çürüttü. İkinci replikasyonda hem hafif hem de hibrit DNA üretildi, dispersif modeli çürüttü, semikonservtif modeli destekledi. İlk replikasyon. İkinci replikasyon. Konsevatif. model. Yarıkorunumlu. Dispersif model.")
35
DNA’nın kendini eşlemesi
DNA Replikasyonu DNA’nın kendini eşlemesi Hızı ve doğruluğu bakımından dikkate çekicidir 10 dan fazla enzim ve diğer prOteinler DNA replikasyonuna katılırlar

36
Başlangıç: Replikasyon orijini-başlangıç noktası
Bir DNA molekülünün replikasyonu Her iki zincirin ayrıldığı replikasyon orjini adlı özel bölgelerde başlar

37
Bir ökaryotik kromozomu
Yüzlerce ve hatta binlerce replikasyon orijini içerebilir Replikasyon iki parental zincirin ayrıldığı ve raplikasyon balonlarının oluşturdukları spesifik bölgelerde başlar Balonlar lateral olarak genişlerler DNA replikasyonu her iki yönde ilerler Sonunda, replikasyon balonları birleşir, ve yeni zincirlerin sentezi tamamlanır 1 2 3 Replikasyon orjini Balon Parental (kalıp) zincir Daughter (yeni) zincir Replikasyon çatalı İki yeni DNA molekülü Ökaryotlarda, her kromozomun kocaman DNA molekülü boyunca birçok yerde DNA replikasyou başlar Bu mikrografta, Chinese hamster cell kültürüne ait DNA boyunca üç replikasyon balonu Görülebilir. (b) (a) 0.25 µm Şekil 12 a, b
 zincir. Daughter (yeni) zincir. Replikasyon çatalı. İki yeni DNA molekülü. Ökaryotlarda, her kromozomun kocaman DNA molekülü boyunca birçok yerde. DNA replikasyou başlar. Bu mikrografta, Chinese hamster cell. kültürüne ait DNA boyunca üç. replikasyon balonu. Görülebilir. (b) (a) 0.25 µm. Şekil 12 a, b.")
38
Yeni DNA zincirinin uzaması
Replikasyon çatalında yeni bir DNA’nın uzaması Yeni oluşan zincirin 3 ucuna nükleotitleri ekleyen DNA polimerazlar adı verilen enzimler tarafından katalizlenir Şekil 13 Yeni zincir Kalıp zincir 5 uç 3 uç Şeker A T Baz C G P OH 5 ucu 3 ucu Pirofosfat 2 P Fosfat Nükleozit trifosfat

39
Çift sarmalın antiparalel yapısı replikasyonu nasıl etkiler?
Antiparalel Uzama Çift sarmalın antiparalel yapısı replikasyonu nasıl etkiler? DNA polimeraz nükleotitleri ekler Sadece yeni oluşan zincirin serbest 3ucuna Kesintisiz zincir boyunca-DNA’nın tek bir kalıp zinciri boyunca DNA polimeraz III tamamlayıcı-kompementer zinciri devamlı olarak sentezleyebilir ve böylelikle replikasyon çatalı ilerler

40
Kesintili zincir’in- DNA’nı n diğer yeni zincirinin uzaması için
DNA polimeraz III replikasyon çatalından uzak yönde çalışmalıdır Kesintili zincir Okazaki parçacıkları adı verilen bir dizi segment olarak sentezlenir ve daha sonra bu parçacıklar DNA ligaz tarafından birleştirilir

41
Replikasyonun nihai yönü
leading ve lagging strandlerin DNA replikasyonu süresince sentezleri Atasal DNA DNA pol Ill sadece 5 yönünde DNA zincirlerini uzatır 1 Okazaki fragmentleri DNA pol III Kalıp zincir Lagging strand 3 2 DNA ligaz Replikasyonun nihai yönü Bir yeni zincir-leading strand, replikasyon çatalı ilerledikçe Devamlı suretle 5 yönünde uzar Diğer yeni zincir-lagging strand, 5 yöünde büyüyen ve Okazaki parçacıkları adlı Kısa segmentlerin 3 yönünde eklenmesiyle oluşur DNA ligaz Okazaki fragmentlerini serbest Uçları arasında bir bağ oluşturarak birleştirir ve Bu devalı bir zincir meydana getirir.uous strand. 4 Şekil 14 3 5 Leading strand

42
DNA polimerazlar bir polinükleotidin sentezini başlatamazlar
Primer RNA DNA polimerazlar bir polinükleotidin sentezini başlatamazlar Çünkü onlar sadece nükleotitleri 3 ucuna eklerler. Başlangıç nükleotit zinciri Bir RNA ya da DNA primeri dir. Sadece bir primer leading strand in sentezi için gereklidir. Ancak kesintili zincir (lagging strand) in sentezi için her bir Okakazi parçacığı ayrı ayrı başlatılmalıdır.
 in sentezi için her bir Okakazi parçacığı ayrı ayrı başlatılmalıdır.")
43
Kalıp DNA RNA Yeni DNA RNA primerine eklenir primerinin başlangıcı
Figure The initiation of DNA synthesis. A complementary RNA primer is first synthesized, to which DNA is added. All synthesis is in the 5’ to 3’ direction. Eventually, the RNA primer is replaced with DNA under the direction of DNA polymerase I. Şekil 15 Figure 11.10

44
Kesikli sentez Sürekli sentez Başlangıç RNA primeri DNA sentezi
Figure Opposite polarity of DNA synthesis along the two strands, necessary because the two strands of DNA run antiparallel to one another and DNA polymerase III synthesizes only in one direction ( to ). On the lagging strand, synthesis must be discontinuous, resulting in the production of Okazaki fragments. On the leading strand, synthesis is continuous. RNA primers are used to initiate synthesis on both strands. Sürekli sentez Başlangıç RNA primeri DNA sentezi Şekil 16
. On the lagging strand, synthesis must be discontinuous, resulting in the production of Okazaki fragments. On the leading strand, synthesis is continuous. RNA primers are used to initiate synthesis on both strands. Sürekli sentez. Başlangıç. RNA primeri. DNA sentezi. Şekil 16.")
45
Şekil 17 1 2 Kalıp zincir Parçacık 1 in RNA primeri 3 4 5 6 7
Primaz RNA nükleotitlerini bir primer şeklinde birleştirir. 1 Replikasyonun tüm yönü 3 5 1 2 Kalıp zincir Parçacık 1 in RNA primeri Okazaki fragment Şekil 17 DNA pol III DNA nükleotitlerini primere ekler ve 1. Okazaki parçacığı oluşur. 2 Birsonraki RNA primeri sağ tarafa ulaştığında DNA pol III ayrılır. 3 İkinci parçacığa primer bağlanır. DNA pol III DNA nucleotitlerini ekler ve o parçacık 1.primere ulaşınca ayrılır 4 DNA pol 1 2. parçacığın 3’ ucuna ekleyerek RNA yı DNA ile yer değiştirir 5 DNA ligaz en yeni DNA ile Parçacık 1 in DNA sı arasında Bir bağ oluşturur. 6 Bu bölgedeki kesintili zincir tamamlanmış olur 7

46
DNA replikasyonuna yardım eden diğer proteinler
Helikaz, topoizomeraz, tek-zincir bağlama proteini Table 16.1

47
DNA replikasyonunun özeti
Şekil 18 Replikasyonun esas yönü Leading strand Lagging Genel bakış Replikasyon çatalı DNA pol III Primaz Primer DNA pol I Parental DNA 5 3 4 3 2 Origin of replication DNA ligaz 1 Helikaz parental ikili sarmalı açar Tek-zincir bağlama proteini açılan kalıp zincirlere bağlanarak sabitler leading zincir DNA pol III tarafından 5 3 yönünde kesintisiz olarak sentezlenir Primaz 5. Okazaki fragmenti için RNA primerinin sentezini başlatır DNA pol III 4. fragmentin sentezini Bitiriyor, 3. fragmetteki RNA primerine Ulaştığında ayrılacak, replikasyon çatalına yönelecek, ve 5. fragment primerinin 3 Ucuna DNA nükleotitlerini ekleyecek 5 DNA pol I ikinci fragmentin 5 ucundan primeri Ayırır, ve üçüncü fragmentin 3 ucuna tek tek DNA nükleotitlerini yerine yerleştirir. Son RNA Nükleotidinin DNA ile yer değiştirmesi şeker-fosfat İskeletinde serbest 3 ucu bırakır 6 DNA ligaz ikinci fragmentin 3 ucunu ilk fragmentin 5 ucuna bağlar 7

48
Proofreading ve DNA onarımı
DNA polimerazlar yeni yapılmış DNA’yı kontrol ederler-proofread Yanlış nükleotitleri yer değiştirirler DNA’nın mismatch onarımın da Onarım enzimleri baz eşleşmesindeki hataları düzeltirler

49
Nükleotit eksizyon onarımında
Enzimler zarar görmüş DNA parçalarını kesip değiştirirler T dimeri DNA ya zarar verir 1 2 Bir nükleaz enzimi hasar görmüş DNA zincirini iki noktadan keser Ve zarar görmüş parça çıkarılır Nükleaz DNA polimeraz 3 Bir DNA polimeraz tarafından onarım sentezi kayıp Nükleotit boşluklarını doldurur DNA ligase 4 DNA ligaz yeni DNA’nın serbest ucunu eski DNA ya yapıştırır ve böylece zincir tamamlanır Şekil 19

50
DNA molküllerinin Uçlarının replikasyonu
Ökaryotik kromozomal DNA’nın sonları Her bir replikasyon evresinde gitgide kısalır Şekil 20 Parental zincir uçları Leading strand Lagging strand Son fragment Önceki fragment RNA primer Kesintili zincir Primerlerin çıkarılması ve DNA ile yer değiştirmesi 3’ ucu varsa mümkündür Primer uzaklaştırılır ancak DNA ile yer değiştirilemez, çünkü DNA polimeraz için gerekli replaced 3ucu yoktur İkinci tur replikasyon Yeni kesintisiz zincir Yeni kesintili zincir 5 Daha sonraki replikasyon turları kısa ve Daha kısa genç (kardeş) DNA molekülleri 5 3
 DNA molekülleri. 5 3")
51
Ökaryotik kromozomal DNA molekülleri
Telomerler: Uçlardaki nükleotit sekansları, DNA moleküllerinin uçlarına yakın genlerin erozyonunu erteler Şekil 21 1 µm

52
Eğer her hücre döngüsünde germ hücrelerinin kromozomları kısalırlarsa
Temel genler üretildikleri gametlerden kaybolacaktır Telomeraz Germ hücrelerinde telomerlerin uzunluğun kontrol eder

53
Genel Bakış: Genetik Bilgi akışı
Gen ifadesi Transkripsiyon ve translasyon: Genler bu iki proses ile proteinleri meydana getirirler Beadle ve Tatum developed: “bir gen bir enzim hipotezi” Bir genin fonksiyonu spesifik bir enzimin üretimini dikte eder Bu hipotezde ufak değişiklikler yapıldı: Genler polipeptit zincirlerini ya da RNA moleküllerini kodlarlar

54
Transkripsiyon ve Translasyonun temel prensipleri
DNA’nın yönetiminde RNA sentezi Mesajcı RNA (mRNA) oluşur Translasyon mRNA’nın yönetiminde meydana gelen, bir polipeptidin gerçek (actual) sentezi Ribozomlarda gerçekleşir
 oluşur. Translasyon. mRNA’nın yönetiminde meydana gelen, bir polipeptidin gerçek (actual) sentezi. Ribozomlarda gerçekleşir.")
55
Prokaryotlarda; Transkripsiyon ve translasyon birlikte gerçekleşir
Prokaryotik hücre. I Bir nükleusu olmayan bir hücrede, transkripsiyonla üretilen mRNA anında ek bir işlem olmaksızın translasyon a uğrar. (a) TRANSLASYON TRANSKRİPSİYON DNA mRNA Ribozom Polipeptit Şekil 22
 TRANSLASYON. TRANSKRİPSİYON. DNA. mRNA. Ribozom. Polipeptit. Şekil 22.")
56
Ökaryotlarda; RNA transkriptleri gerçek mRNA meydana gelmeden önce değişikliğe uğrar Şekil 23 Ökaryotik hücre. Nukleus transkripsiyon için ayrı bir kompartıman sağlar. Orjinal RNA transkripti, öncül mRNA, nukleusu mRNA olarak terk etmeden önce çeşitli yollarla işlemlerden geçer (b) TRANSKRİPSİYON RNA PROSESİ TRANSLASYON mRNA DNA öncül-mRNA Polipeptit Ribozom Hücre zarfı
 TRANSKRİPSİYON. RNA PROSESİ. TRANSLASYON. mRNA. DNA. öncül-mRNA. Polipeptit. Ribozom. Hücre zarfı.")
57
Hücreler hücresel bir komuta zinciri ile yönetilirler
DNA RNA protein Genetik Kod Kaç baz bir aminoaside karşılık gelir? Kodonlar: Üçlü Bazlar Genetik bilgi Çakışmayan üçlü baz serileri, ya da kodonlar olarak kodlanırlar

58
Transkripsiyon sırasında (süresince)
Geni bir mRNA molekülünün uzunluğu boyunca baz sıraları belirler Şekil 24 DNA molekülü Gen 1 Gen 2 Gen 3 DNA zinciri (kalıp) TRANSKRİPSİYON mRNA Protein TRANSLASYON Amino asit A C G T U Trp Phe Gly Ser Kodon 3 5
 TRANSKRİPSİYON. mRNA. Protein. TRANSLASYON. Amino asit. A. C. G. T. U. Trp. Phe. Gly. Ser. Kodon. 3 5")
59
Cracking the Code mRNA’daki bir kodon;
Ya bir amino aside transle olur ya da translasyonel stop sinyali olarak görev yapar Şekil 25 İkinci mRNA bazı U C A G UUU UUC UUA UUG CUU CUC CUA CUG AUU AUC AUA AUG GUU GUC GUA GUG Met or start Phe Leu lle Val UCU UCC UCA UCG CCU CCC CCA CCG ACU ACC ACA ACG GCU GCC GCA GCG Ser Pro Thr Ala UAU UAC UGU UGC Tyr Cys CAU CAC CAA CAG CGU CGC CGA CGG AAU AAC AAA AAG AGU AGC AGA AGG GAU GAC GAA GAG GGU GGC GGA GGG UGG UAA UAG Stop UGA Trp His Gln Asn Lys Asp Arg Gly İlk mRNA bazı (5 uç) Üçüncü mRNA bazı (3 uç) Glu
 Üçüncü mRNA bazı (3 uç) Glu.")
60
Genetik kod büyük ölçüde (neredeyse) evrenseldir
Kodonlar belli bir polipeptidin üretilmesi için doğru reading frame i okumalıdır Genetik Kodun evrimi Genetik kod büyük ölçüde (neredeyse) evrenseldir En basit bakteriden en karmaşık hayvanlara organizmalar tarafından paylaşılır
 evrenseldir. En basit bakteriden en karmaşık hayvanlara organizmalar tarafından paylaşılır.")
61
Transkripsiyon: DNA direktifli RNA sentezi
RNA polimeraz tarafından katalize edilir, enzim DNA zincirlerini ayırır ve RNA nükleotitlerini biraraya getirir RNA da T yerine U bazı A ile eşleşir

62
Bir RNA Transkriptinin sentezi
Transkripsiyon basamakları Başlama Uzama Sonlanma Şekil 26 Promotor Transkripsiyon birimi RNA polimeraz Başlangıç noktası 5 3 Geri birleşen RNA transkripti tamamlanmş RNA transkripti Ayrılmış DNA transkriptit Kalıp DNA zinciri 1 Initiation. RNA polimeraz promotora bağlandıktan sonrai, DNA zincirleri ayrılır, ve polimeraz kalı zincir üzerindeki başlangıç noktasında RNA sentezini başlatır. 2 Elongation. Polimeraz DNA’yı açarak aşağıya doğru ilerler, ve RNA transkirptini 5 3 ynünde uzatır. Transkripsiyon yolunda DNA zincirleri tekrar eski ikili sarmal yapısına döner. 3 Termination. Sonunda, RNA transkripti salınır ve polimeraz DNA’dan ayrılır

63
Elongation- Uzama RNA polimeraz Kalıp olmayan DNA zinciri RNA nükleotitleri 3 ucu C A E G U T 3 5 Yeni sentezlenen Transkripsiyon yönü (“aşağıya doğru”) Kalıp DNA zinciri
 Kalıp DNA zinciri.")
64
RNA Polimeraz Bağlanması ve Transkripsiyonun başlaması
Promotorlar RNA sentezinin başlaması için işaret verirler Transkripsiyon faktörleri Ökaryotik RNA polimerazın promotoz sekanslarını tanımalarına yardımcı olurlar Şekil 27 TRANSCRIPTION RNA PROCESSING TRANSLATION DNA Pre-mRNA mRNA Ribosome Polypeptide T A TATA box Başlangıç noktası Kalıp DNA zinciri 5 3 Transkripsiyon faktörleri Promotor RNA polimeraz II RNA transkripti Transkripsiyon başlangıç kompleksi Ökaryotik promotorlar 1 Çeşitli transkripsiyon faktörleri 2 İlave transkripsiyon faktörleri 3

65
RNA zincirinin Uzaması
RNA polimeraz DNA boyunca hareket ettiğinden RNA nükleotitleri ile eşleştirme için bir defada 10 ila 20 DNA bazınının açığa çıktığı bir sürede ikili sarmalı açmaya devam eder Transkripsiyonun sonlanması Terminasyon mekanizması Prokaryot ve ökaryotlarda farklıdır Ökaryotik hücreler transkripsiyon sonrasında RNA yı değiştirirler Ökaryot nükleusundaki enzimler Genetik mesajlar sitoplazmaya gönderilmeden önce belirli şekillerde pre-mRNA modifiye edilir.

66
mRNA uçlarının değişmesi
Pre-mRNA’nın her ucu belirli şekillerde modifiye edilirler 5 ucu modifiye bir nükleotit cap alır 3 ucu bir poly-A ucu alır Modifiye bir guanin nükleotidi 5 ucuna eklenir 50 ila 250 adenin nükleotidi 3 ucuna eklenir Protein-kodlayan segment segment Poliadenilasyon sinyali Poly-A kuyruğu 3 UTR Stop kodon Start kodon 5 Cap 5 UTR AAUAAA AAA…AAA TRANSCRIPTION RNA PROCESSING DNA Pre-mRNA mRNA TRANSLATION Ribosome Polypeptide G P 5 3 Şekil 28

67
Bölünmüş genler ve RNA ekleme (Split Genes and RNA Splicing)
İntronları çıkarıp ekzonları birleştirir TRANSCRIPTION RNA PROCESSING DNA Pre-mRNA mRNA TRANSLATION Ribosome Polypeptide 5 Cap Exon Intron 1 5 30 31 104 105 146 3 Poly-A kuyruğu Intronlar kesilir ve ekzonlar biraraya getirirli Kodlayan segment 3 UTR Şekil 29

68
RNA transkripti (pre-mRNA)
RNA splicing Bazı durumlarda spliceosom lar tarafından gerçekleştirirlir RNA transkripti (pre-mRNA) Exon 1 Intron Exon 2 Diğer proteinler Protein snRNA snRNPs Spliceosome bileşenleri Kesilen intron mRNA 5 1 2 3 Şekil 30
 Exon 1. Intron. Exon 2. Diğer proteinler. Protein. snRNA. snRNPs. Spliceosome. bileşenleri. Kesilen intron. mRNA. 5 Şekil 30.")
69
İntronların varlığı alternatif RNA splicing e izin verir
Ribozimler Ribozimler Enzim olarak görev yapan ve RNA ekleyebilen (splice) katalitik RNA molekülleridir İntronların işlevsel ve evrimsel önemi İntronların varlığı alternatif RNA splicing e izin verir
 katalitik RNA molekülleridir. İntronların işlevsel ve evrimsel önemi. İntronların varlığı alternatif RNA splicing e izin verir.")
70
Proteinler çoğunlukla modüler mimariye sahiptirler
Domain denilen ayrı yapısal ve işlevsel bölgelerden oluşurlar Çoğu durumda Farklı ekzonlar bir proteinde farklı domainleri kodlarlar Gene DNA Exon 1 Intron Exon 2 Exon 3 Transcription RNA processing Translation Domain 3 Domain 1 Domain 2 Polipeptit Şekil 31

71
Translasyon-RNA direktifli polipeptit sentezi
Translasyonun moleküler bileşenleri Bir hücre mRNA mesajını proteine transle eder ve bunun transfer RNA (tRNA) yardımıyla yapar
 yardımıyla yapar.")
72
Translasyon: temel konsept
TRANSCRIPTION TRANSLATION DNA mRNA Ribosome Polypeptide Polipeptit Amino asitler Amino asit bağlı tRNA Ribozom tRNA Antikodon Trp Phe Gly A G C U Kodonlar 5 3 Şekil 32

73
tRNA’nın yapısal ve fonksiyonel özellikleri
Bir tRNA molekülü Sadece 80 nükleotit uzunluğunca tek zincirli bir RNA da oluşur Kabaca L-şeklindedir A C C Şekil 33 İki boyutlu yapı. 3’ ucundaki amino asit bağlanma yerinin baz dizisi olarak dört baz çiftli bölgeler ve üç loop tüm tRNA lar için karakteristiktir. Antikodon üçlüsü her tRNA tipi için bözgüldür (Yıldızlar tRNA karakteristiği olarak kimyasal olarak modifiye olmuş bazları işaret eder) (a) 3 C A G U * 5 Amino asit bağlanma yeri Hidrojen bağları Antikodon tRNA molekülleri özdeş değildir Her biri bir ucunda belirli amino asitleri taşırlar Her biri diğer ucunda antikodon a sahiptir
 (a) 3 C. A. G. U. * 5 Amino asit. bağlanma yeri. Hidrojen bağları. Antikodon. tRNA molekülleri özdeş değildir. Her biri bir ucunda belirli amino asitleri taşırlar. Her biri diğer ucunda antikodon a sahiptir.")
74
5 3 A G Şekil 34 Amino asit bağlanma yeri Hydrogen bonds Antikodon
(b) Üç boyutlu yapı Sembolik şekli Amino asit bağlanma yeri Hydrogen bonds Antikodon A G 5 3 (c) Şekil 34
 Üç boyutlu yapı. Sembolik şekli. Amino asit bağlanma yeri. Hydrogen. bonds. Antikodon. A. G. 5 3 (c) Şekil 34.")
75
Ribozomlar Protein sentezi boyunca tRNA antikodonlarının mRNA kodonları ile spesifik olarak eşleşmesini kolaylaştırır Ribozom altbirimleri Ribozomal RNA (rRNA) denilen protein ve RNA dan oluşturulur TRANSCRIPTION TRANSLATION DNA mRNA Ribosome Polypeptide Çıkış tüneli Büyüyen polipeptit tRNA molekülleri E P A Büyük altbirim Küçük altbirim Fonksiyonel ribozomun bilgisayar modeli. Bir bakteri ribozom yapısı (ökaryot ribozom da kabaca benzerdir) (a) 5 3 Şekil 36
 denilen protein ve RNA dan oluşturulur. TRANSCRIPTION. TRANSLATION. DNA. mRNA. Ribosome. Polypeptide. Çıkış tüneli. Büyüyen polipeptit. tRNA. molekülleri. E. P. A. Büyük altbirim. Küçük altbirim. Fonksiyonel ribozomun bilgisayar modeli. Bir bakteri ribozom yapısı (ökaryot ribozom da kabaca benzerdir) (a) 5 3 Şekil 36.")
76
Ribozom tRNA için üç bağlanma bölgesine sahiptir
P bölgesi A bölgesi E bölgesi P bölgesi (Peptidil-tRNA Bağlanma bölgesi) E bölgesi (Çıkış bölgesi) mRNA Bağlanma bölgesi A bölgesie (Aminoaçil- tRNA bağlanma bölgesi) Büyük alt birim Küçük altbirim Bağlanma bölgelerini gösteren şematik model.. (b) E P A Şekil 36
 E bölgesi. (Çıkış bölgesi) mRNA. Bağlanma bölgesi. A bölgesie (Aminoaçil- tRNA bağlanma bölgesi) Büyük alt birim. Küçük altbirim. Bağlanma bölgelerini gösteren şematik model.. (b) E. P. A. Şekil 36.")
77
Amino ucu Büyüyen polipeptit Polipeptit zincirine eklenecek bir sonraki amino asit tRNA mRNA Kodonlar 3 5 mRNA ve tRNA lı şematik bir model tRNA antikodonunun bir mRNA kodonu ile eşleştiği bir bağlanma bölgesine uyum gösterir. P bölgesi büyüyen polipeptide bağlı tRNA yı tutar. A bölgesi polipeptit zincirine yeni eklenecek amino asiti taşıyan tRNAyı tutar. Deşarj olan tRNA E bölgesinden ayrılır. (c) Şekil 36
 Şekil 36.")
78
Bir polipeptidin inşaası
Translasyon basamakları Initiation- başlama Elongation-uzama Termination-sonlanma

79
Ribozom etkileşimi ve translasyonun başlaması
Translasyonun başlangıç basamağı mRNA’yı polipeptidin ilk amino asidini taşıyan tRNA’yı, ve ribozomun iki altbirimini biraraya getirir Büyük ribozom altbirimi Ribozomal altbirimiin gelişi başlangıç kompleksini tamalmlar Başlama faktörleri adlı proteinler tüm translasyon bileşenlerini biraraya getirmek için gereklidir. GTPbiraraya gelmek için gerekli Enerjiyi sağlarBaşlatıcı tRNA P bölgesindedir; A bölgesibir sonraki Aminoasidi taşıyan tRNA için açıktır 2 Başlatıcı tRNA mRNA mRNA bağlanma bölgesi Küçük ribozom altbirimi Translasyon başlangıç kompleksi P bölgesi GDP GTP Start codon Küçük ribozomal altbirim mRNA molekülüne bağlanır. Prokaryotik hücrede, bu bölgedeki mRNA bbağlanma bölgesi başlangıç kodonunun hemen üstündeki bir spesifik nükleotit sekansınaı tanır. Antikodon UAC li başlatıcı tRNA, başlangıç kodonu AUG ile eşleşir. Bu tRNA metiyonin aminoasidini taşır. 1 Met U A C G E 3 5 Şekil 38

80
Polipeptit zincirinin uzaması
Translasyonun uzama basamağı Amino asitler bir önceki aminoaside sırayla eklenirler Kodon tanıma. Gelen aminoaçil tRNA’nın antikodonu A bölgesindeki komplementer mRNA ile eşleşir. GTP nin hidrolizi bu basamağın doğruluğu ve efficiency si ni artırır 1 Şekil 39 Polipeptidin amino ucu mRNA Bir sonraki aminoaçil tRNA İçin hazır ribozom E P A GDP GTP 2 site 5 3 TRANSCRIPTION TRANSLATION DNA Ribosome Polypeptide Translokasyon. Ribozom A bölgesindeki tRNA yı P bölgesine geçirir. P bölgesindeki boş tRNA E bölgesine hareket ettirilir ve buradan salınır. mRNA ona bağlanan tRNA ile birlikte hareket eder, böylece A bölgesinde translate edilecek bir sonraki kodonu getirir. 3 Peptit bağı oluşumu Büyük altbirimin rRNA molekülü A bölgesindeki yeni aminoasit ile P bölgesindeki büyüyen aminoasidin karboksil ucu arasında peptit bağı oluşumunu katalizler. Bu basamak A bölgesindeki tRNA yı polipeptide bağlar 2

81
Translasyonun sonlanması
Translasyonun son basamağı terminasyondur Ribozom mRNA daki stop kodonuna ulaşır Şekil 40 Salınma faktörü Serbest polipeptit Stop kodonu (UAG, UAA, veya UGA) 5 3 Bir ribozom mRNA daki stop kodonuna ulaştığında, ribozomun A bölgesi tRNA yerine release faktör denilen proteini kabul eder. 1 Release faktörü P bölgesindeki tRNA ile polipeptit zincirinin son aminoasidi arasındaki bağı hidroliz eder. Polipeptit ribozomdan ayrılır 2 3 İki ribozom altbirimi ve diğer komponentler kompleksten ayrılır
 5 3 Bir ribozom mRNA daki stop. kodonuna ulaştığında, ribozomun. A bölgesi tRNA yerine release. faktör denilen proteini kabul eder. 1. Release faktörü P bölgesindeki. tRNA ile polipeptit zincirinin. son aminoasidi arasındaki. bağı hidroliz eder. Polipeptit ribozomdan ayrılır İki ribozom altbirimi ve diğer. komponentler kompleksten ayrılır.")
82
Poliribozomlar Tek bir mRNA molekülünü aynı anda çevirebilen çok sayıdaki ribozomlardır Poliribozom Şekil 42 Büyüyen polipeptitler Tamamlanmış polipeptit Gelen ribozom altbirimleri mRNA başlangıcı (5 uç) sonu (3 ucu) Poliribozom mRNA molekülü poliribozom adlı ribozom jümeleri tarafından aynı anda çevrilirler (a) Ribosomes Prokaryotik hücrede büyük bir poliribozom (TEM). 0.1 µm (b)
 sonu. (3 ucu) Poliribozom. mRNA molekülü poliribozom adlı ribozom jümeleri tarafından aynı anda çevrilirler. (a) Ribosomes. Prokaryotik hücrede büyük bir poliribozom (TEM). 0.1 µm. (b)")
83
Fonksiyonel proteinin tamamlanması ve hedeflenmesi
Polipeptit zincirleri Translasyon işleminden sonra modifikasyona uğrar Protein Katlanması ve Post-Translasyonel Modifikasyonlar Translasyon sonrası Proteinler üç boyutlu yapılarının etkileyen bir biçimde modifiye olabilirler

84
Belirli lokasyonlara polipeptitlerin hedeflenmesi
Hücrelerde iki tip ribozom populasyonu yer alır: Serbest ve bağlı Serbest ribozomlar sitozolde yer alırlar Tüm proteinlerin sentezini başlatırlar

85
Endomembran sistemine yada sekresyona hedeflenen proteinler
ER ye taşınmalıdırlar Bunu sahip olduğu sinyal tanıma parçasını (a signal-recognition particle (SRP)) bağlayan sinyal peptitler sayesinde gerçekleştirir
) bağlayan sinyal peptitler sayesinde gerçekleştirir.")
86
Proteinleri ER ye taşıyan sinyal mekanizması
Polypeptide synthesis begins on a free ribosome in the cytosol. 1 An SRP binds to the signal peptide, halting synthesis momentarily. 2 The SRP binds to a receptor protein in the ER membrane. This receptor is part of a protein complex (a translocation complex) that has a membrane pore and a signal-cleaving enzyme. 3 The SRP leaves, and the polypeptide resumes growing, meanwhile translocating across the membrane. (The signal peptide stays attached to the membrane.) 4 The signal- cleaving enzyme cuts off the signal peptide. 5 The rest of the completed polypeptide leaves the ribosome and folds into its final conformation. 6 Şekil 43 Ribosome mRNA Signal peptide Signal- recognition particle (SRP) SRP receptor protein Translocation complex CYTOSOL removed ER membrane Protein ERLUMEN
 that has a membrane pore. and a signal-cleaving enzyme. 3. The SRP leaves, and. the polypeptide resumes. growing, meanwhile. translocating across the. membrane. (The signal. peptide stays attached. to the membrane.) 4. The signal- cleaving. enzyme. cuts off the. signal peptide. 5. The rest of. the completed. polypeptide leaves. the ribosome and. folds into its final. conformation. 6. Şekil 43. Ribosome. mRNA. Signal. peptide. Signal- recognition. particle. (SRP) SRP. receptor. protein. Translocation. complex. CYTOSOL. removed. ER. membrane. Protein. ERLUMEN.")
Benzer bir sunumlar


>")



 ZM106 Biyokimya 11. Hafta.>")





>")



 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")
>")

