Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Araş.Gör.Ayhan ÜNLÜ İstanbul Tıp Fak. Biyofizik A.B.D.
Homeogenler Araş.Gör.Ayhan ÜNLÜ İstanbul Tıp Fak. Biyofizik A.B.D.

2
GENETİK DÜZENLEMEDE – SOMATİK KALITIM EVRELERİ
1- Yumurta hücresi düzeyinde düzenleme 2. Zigot evresinde düzenleme 3. Gastrulasyon-Embriyogenez evresinde düzenleme 4. Fetus dönemi düzenleme

3
GENETİK DÜZENLEMEDE – SOMATİK KALITIM EVRELERİ
1- Yumurta hücresi düzeyinde düzenleme : Yumurta hücresinde bulunan anterioposterior gradiyent farkı fertilizisyon öncesi yumurta hücresinden meydana gelecek embriyonun anteriyor ve posteriyor kısmını verecek bölgeler öncelikle belirlenmektedir. Burada sadece yumurta hücresiyle sınırlı bazı regülatör-modülatör proteinler yumurta-polarity ve segmentasyondan sorumlu (25 adet tanımlanmıştır) genler görev almaktadırlar. 2. Zigot evresinde düzenleme : Bu evrede yine çoğu yumurtadan orijin alan ve döllenmeyi takiben aktive olan zigotik-effekt genler olarak bilinen; remodelling faktörler, transkripsiyonel faktörler ve kromatin bağlayıcı özgül proteinler gibi düzenleyici moleküllerin görev aldıkları saptanmıştır. Bu genlerin görevi yumurta ve sperm çekirdeklerinin kaynaşmasını sağlamak ve hücre bölünmesi öncesi görev yapan proteinlerin bazı regülasyonunda görev alırlar.
 genler görev almaktadırlar. 2. Zigot evresinde düzenleme : Bu evrede yine çoğu yumurtadan orijin alan ve döllenmeyi takiben aktive olan zigotik-effekt genler olarak bilinen; remodelling faktörler, transkripsiyonel faktörler ve kromatin bağlayıcı özgül proteinler gibi düzenleyici moleküllerin görev aldıkları saptanmıştır. Bu genlerin görevi yumurta ve sperm çekirdeklerinin kaynaşmasını sağlamak ve hücre bölünmesi öncesi görev yapan proteinlerin bazı regülasyonunda görev alırlar.")
4
3. Gastrulasyon-Embriyogenez evresinde düzenleme : Bu evrede görev alan en önemli gen grubunun yine yumurta hücresine ait 8 çift oldukalı saptanan pair-rule ve 10 adet oldukarı saptanan segment polarity genlerdir. Bu gen grubu her tür için farklı olmakla birlikte embriyogenezin 2, 8 ve 16 hücrelik bölünme evrelerinde inaktive edilirler. Örneğin farelerde zigot 2, koyun ve insanda 16 hücrelik embriyo olana kadar görev yapmaktadırlar. Bu sayı türe göre değişmektedir. Zigot hücresinin maksimum 16 hücreye kadar bölünmesinden sorumlu gen grubudur. Bu genler sadece totipotent hücrelerde görev alırlar.

5
4. Fetus dönemi düzenleme : Bu dönem, fetus hücrelerine ait genlerin ifade edilmesiyle başlar. Bu dönemden sonra yumurta regülasyonu yerini fetus gen regülasyonuna terk eder. Görev yapan genler homeotik ya da homeodometik (hox) gen ailesi olarak adlandırılır. Türlerarası somatik doku farklılaşmasından birinci dereceden bu gen grubu sorumludur. Bir dokunun normal ya da anormal bir şekilde farklılaşması bu gen grubunun normal ve zamanında fonksiyon yapmasına bağlıdır. İlk kez blastoderm evresinde aktive olurlar. Memelilerde 4 adet homolog homeotik kompleks genin varlığı saptanmıştır. Homeodometik seçici genleri meydana getiren homeodomain zincir genleri evrim süresince korunan ve en az varyasyon gösteren genlerdir. Herbiri 60 aa uzunluğunda protein sentezinden sorumlu bç uzunluğunda regülatör alt birimlerinden ibaret genlerdir. Genlerin regülatör alt birimlerini meydana getiren diziler, segmentasyonel ve yumurta -polarity gen ürünlerine özgül bağlantı bölgeler içerirler. Vücudun segmentasyonunda spesifik görev yapan bu gen grubudur. Moleküler mekanizmaları kesin olarak bilinmemekle birlikte regülatör alt birimleri aracılığıyla aktive ve inhibe edildikleri sanılmaktadır.

6
Erişkin bir çok hücreli canlı ne kadar kompleks vücut yapısına sahip olursa olsun, yapı ve işlev bakımından çok farklı olan kaç çeşit hücreden, kaç çeşit dokudan ve kaç organdan meydana gelmiş olursa olsun; başlangıç noktası tek bir hücredir.

7
“ Döllenmiş yumurta ve sonraki erken embryo hücreleri salt (=tam) potansiyelli (totipotent) hücrelerdir.
 potansiyelli (totipotent) hücrelerdir.")
8
Gelişme Regülasyonu ? Bu zigot daha sonra milyonlarca (veya milyarlarca) hücreden oluşacak organizmayı oluşturacaktır. Bu gelişime hangi mekanizmalar etkilidir? Zigot kompleks organizmanın oluşabilmesi için gereken tüm genetik bilgiye sahip olduğu gibi bu genlerden hangilerinin ne zaman, nerede ve ne kadar çalıştırılması gerektiği bilgisine, genetik bilgiyi en doğru şekilde kullanma yeteneğine de sahiptir. Bu yeteneğe, gen regülasyonu denir.
 hücreden oluşacak organizmayı oluşturacaktır. Bu gelişime hangi mekanizmalar etkilidir Zigot kompleks organizmanın oluşabilmesi için gereken tüm genetik bilgiye sahip olduğu gibi bu genlerden hangilerinin ne zaman, nerede ve ne kadar çalıştırılması gerektiği bilgisine, genetik bilgiyi en doğru şekilde kullanma yeteneğine de sahiptir. Bu yeteneğe, gen regülasyonu denir.")
9
Gelişim, Regülatör Proteinler tarafından kontrol edilir
Gelişim, Regülatör Proteinler tarafından kontrol edilir. Meyve sineğinin yaşam döngüsünde, yumurtadan ergine geçişte tam bir metamorfoz görülür.

10
Drosophila yumurtası, 15 hemşire hücre (nurse cell) ile birlikte bulunur ve bir folikül hücre tabakası ile çevrelenmiştir. Yumurta hücresi oluşunca (döllenmeden önce) hemşire ve folikül hücrelerde sentezlenen mRNA ve proteinler, hücre içinde birikir. Bu protein ve mRNA'ların gelişimde kritik rolleri vardır.
 hemşire ve folikül hücrelerde sentezlenen mRNA ve proteinler, hücre içinde birikir. Bu protein ve mRNA ların gelişimde kritik rolleri vardır..")
11
Döllenmiş bir yumurta yumurtlanınca, nükleus bölünür ve yavru nükleuslar senkronize olarak her 6-10 dakikada bir bölünmeye devam ederler. Etraflarında plazma membranı oluşmayan bu nükleuslar, yumurta sitoplazması içinde (veya sinsityumda) dağılırlar. Bölünmelerin devrelerinde nükleuslar, nükleusu çevreleyen zengin sitoplazmadan meydana gelen bir tek tabaka oluşturacak şekilde yumurtanın dış tabakasına doğru hareket ederler. Böylece sinsitiyal blastoderm oluşur. Bir kaç ek bölünmeden sonra membran, invaginasyon yaparak nükleusları birbirinden ayırarak hücresel blastodermi meydana getirecek hücreleri oluşturur. Bu basamakta çeşitli hücrelerdeki mitotik senkroni kaybolur.
 dağılırlar. Bölünmelerin devrelerinde nükleuslar, nükleusu çevreleyen zengin sitoplazmadan meydana gelen bir tek tabaka oluşturacak şekilde yumurtanın dış tabakasına doğru hareket ederler. Böylece sinsitiyal blastoderm oluşur. Bir kaç ek bölünmeden sonra membran, invaginasyon yaparak nükleusları birbirinden ayırarak hücresel blastodermi meydana getirecek hücreleri oluşturur. Bu basamakta çeşitli hücrelerdeki mitotik senkroni kaybolur..")
12
Hücrelerin gelişimsel kaderini, hemşire ve folikül hücreleri tarafından sentezlenip yumurtada depolanan mRNA ve proteinler belirler. lokus konsantrasyonları veya aktiviteleri, çevreleyici dokunun özel şekil ve yapıalmasını sağlayan proteinler, "morfogen" olarak adlandırılırlar. Bu proteinler, patern regüle edici genlerin ürünleridir. Patern regüle edici genlerin 3 ana sınıfı vardır. maternal genler segmentasyon genler homeotik genlerdir.

13
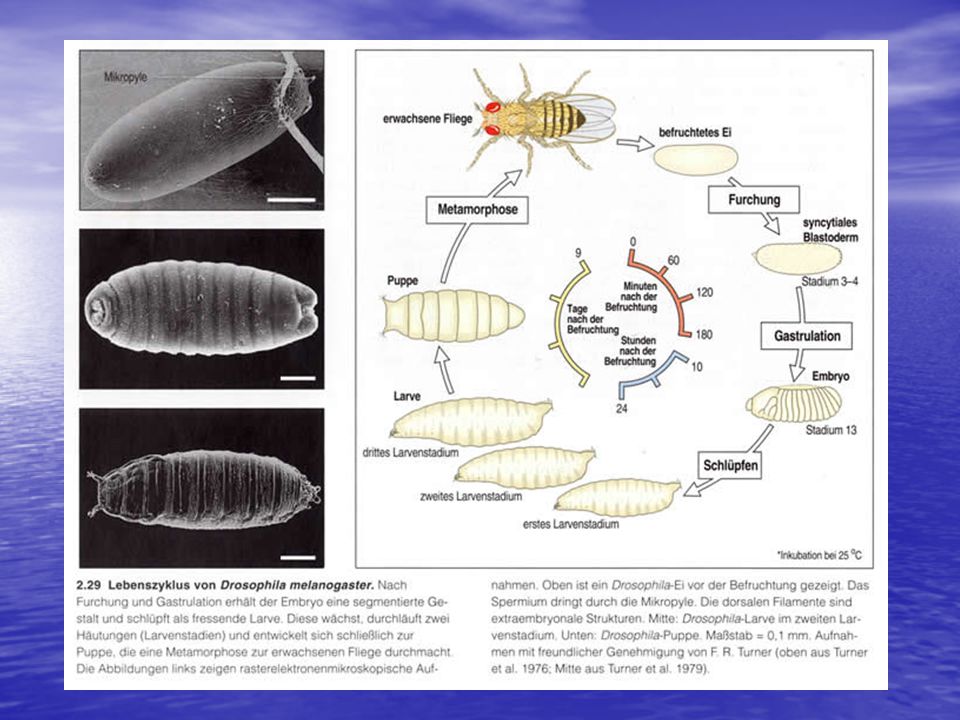
Drosophila ergin olmadan önce, döllenmiş yumurtadan başlayarak beş farklı evre geçirir: embriyo, üç larva aşaması ve pupa oluşum aşaması. Döllenmeden aşağı yukarı 10 gün sonra pupadan ergin sinek gelişir . Drosophila yumurtasında dışarıdan pekçok yapı görülür. Bu yapılar, anterior (ön), posterior (arka), dorsal (sırt) ve ventral (karın) bölgelerin şeklini verir. Yumurtanın anterior ucu mikropil denen ve spermin yumurtaya girmesi için özelleşmiş konik yapıyı içerir, posterior uç ise yuvarlaktır ve bir seri aeropillerle (gelişim sırasındaki gaz giriş çıkışını sağlayan açıklıklar) tanınır. Yumurtanın dorsal bölgesi düzdür ve koryonik çıkıntılar içerir, ventral bölge ise yuvarlaktır. Yumurtanın içerisinde, sitoplazma anneden gelen bir dizi moleküler gradiyent şeklinde düzenlenmiştir. Bu moleküler yapılar, gelişim sırasında çekirdeğin embriyonun özgül bölgelerine doğru gitmesinde anahtar rol oynar.
, posterior (arka), dorsal (sırt) ve ventral (karın) bölgelerin şeklini verir. Yumurtanın anterior ucu mikropil denen ve spermin yumurtaya girmesi için özelleşmiş konik yapıyı içerir, posterior uç ise yuvarlaktır ve bir seri aeropillerle (gelişim sırasındaki gaz giriş çıkışını sağlayan açıklıklar) tanınır. Yumurtanın dorsal bölgesi düzdür ve koryonik çıkıntılar içerir, ventral bölge ise yuvarlaktır. Yumurtanın içerisinde, sitoplazma anneden gelen bir dizi moleküler gradiyent şeklinde düzenlenmiştir. Bu moleküler yapılar, gelişim sırasında çekirdeğin embriyonun özgül bölgelerine doğru gitmesinde anahtar rol oynar.")
14
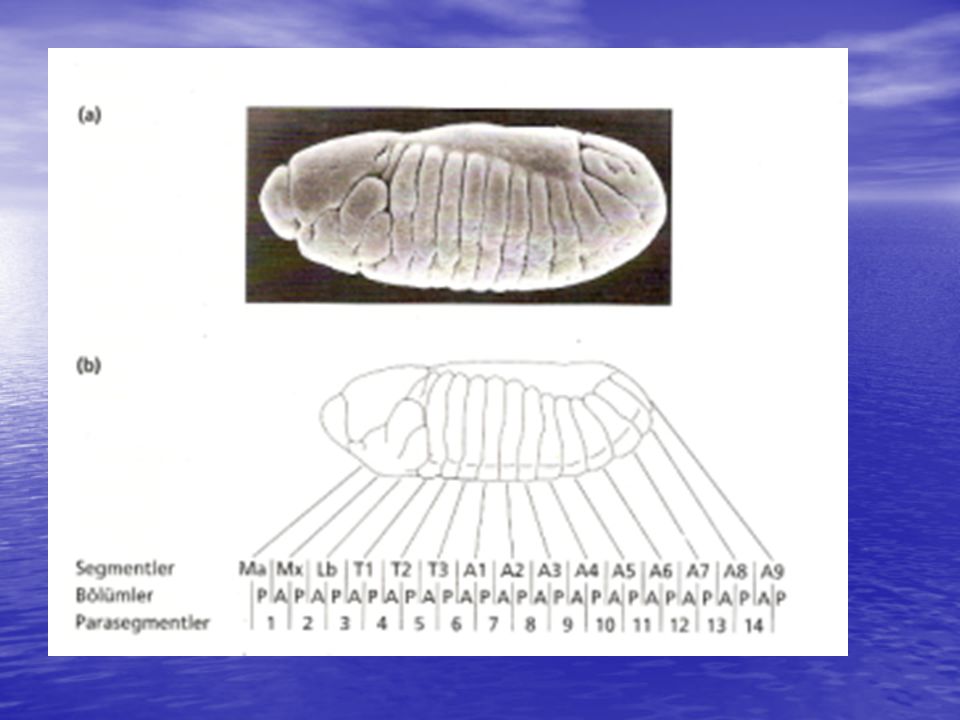
Embriyonun en önemli karakteristiklerinden biri, polaritesi; yani, hayvanın anteryör ve posteryör kısımlarının, dorsal ve ventral kısımları gibi birbirinden tamamen ayrılmış olması; bir diğeri de metamerizmidir (embriyo vücudunun, her birinin karakteristik özelliği olan seri olarak tekrarlayan segmentlerden oluşması). 5´ 3´ anterior posterior

15
Bithorax controls development of parasegments 5-14
Antennapedia controls anterior parasegments Bithorax is best understood (discussed first) 3 genes (regulated by gap & pair-rule) ultrabithorax (parasegment 5 onward) abdominal-A (parasegment 7 onward) abdominal-B (parasegment 10 onward) abdominal-B suppresses ultrabithorax (low in 14) 5´ 3´ anterior posterior
 3 genes (regulated by gap & pair-rule) ultrabithorax (parasegment 5 onward) abdominal-A (parasegment 7 onward) abdominal-B (parasegment 10 onward) abdominal-B suppresses ultrabithorax (low in 14) 5´ 3´ anterior. posterior.")
19
Gelişim sırasında bu segmentler, baş, toraks ve abdomeni oluşturacak şekilde organize olurlar. Erişkin toraksının her segmenti, farklı bir ilave setine (kanatlar ve bacaklar...) sahiptir. Bu kompleks paternin gelişimi, genetik kontrol altındadır. Vücut organizasyonunu kuvvetli bir şekilde etkileyen bir grup protein keşfedilmiştir.
 sahiptir. Bu kompleks paternin gelişimi, genetik kontrol altındadır. Vücut organizasyonunu kuvvetli bir şekilde etkileyen bir grup protein keşfedilmiştir..")
21
Drosopbila'da anatomik gelişimle ilgili olarak elde edilen bilgiler oldukça faydalı olmasına rağmen, genetik analizler embriyo oluşumu ile ilgili olaylar hakkında daha önemli bilgileri sağlar. Embriyo gelişimini kontrol eden genler ya maternal-etkili genler (anneden gelen etkin genler) ya da zigotik genlerdir. Maternal etkili genler, ürünleri (mRNA ve proteinler) oogenez sırasında gelişmekte olan yumurta içinde depolanan genlerdir. Bu ürünler yumurta sitoplazmasına düzgün bir şekilde dağılmış olmasına rağmen bazen katmanlar halinde ve bazen de yumurtanın belirli bir bölgesinde yoğunlaşırlar.
 ya da zigotik genlerdir. Maternal etkili genler, ürünleri (mRNA ve proteinler) oogenez sırasında gelişmekte olan yumurta içinde depolanan genlerdir. Bu ürünler yumurta sitoplazmasına düzgün bir şekilde dağılmış olmasına rağmen bazen katmanlar halinde ve bazen de yumurtanın belirli bir bölgesinde yoğunlaşırlar..")
22
Zigotik genler zigotun kendi gelişimini sağlayan genlerdir ve bu nedenle döllenmeden sonra transkripsiyona uğrarlar. Bu sınıftaki mutasyonlar embriyonun ölmesine neden olur; çekinik zigot mutasyonu için heterozigot olan iki sineğin çaprazlaması sonucu embriyoların dörtte biri (homozigotlar) gelişemez. Drosopbila'da zigotik genlerin pekçoğu, maternal etkili proteinlerin dağılımına bağlı olarak transkripsiyon profili gösterir.
 gelişemez. Drosopbila da zigotik genlerin pekçoğu, maternal etkili proteinlerin dağılımına bağlı olarak transkripsiyon profili gösterir..")
24
Maternal Genler Maternal genler, döllenmemiş embriyoda ekspresse edilirler ve oluşan mRNA'lar, döllenmeye kadar etkisiz kalır. Bunlar, hücresel blastoderm oluşumuna kadar olan çok erken gelişimde ihtiyaç duyulacak proteinleri sağlarlar. Maternal mRNA'ların kodladığı proteinlerden bazıları, gelişen embriyonun erken evrelerdeki bölümlenmiş organizasyonunu yönlendirir. Bunlar, polariteyi sağlayan yapılardır.

25
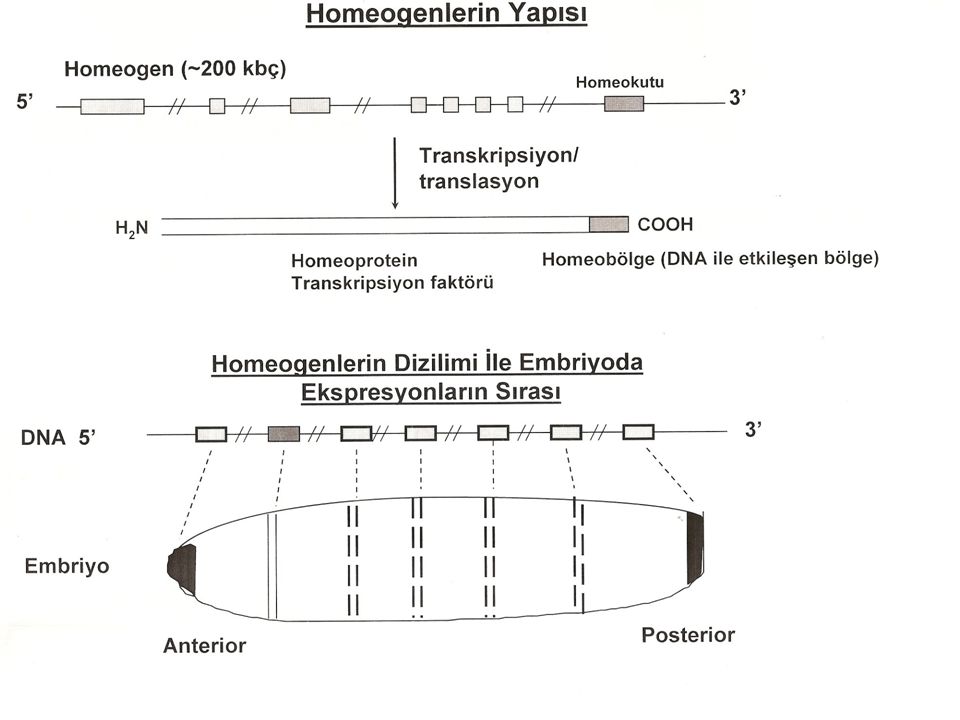
Homeotik Genler Homeotik genler ise, daha sonraki basamaklarda ekspresse edilirler. HOMEOTİK GENLER; her bir homeotik gen embriyogenezis sırasında farklı zamanda ve farklı dokularda ifade edilir. Homeotik genler,vücut kısımlarının nasıl şekilleneceğini ya da kalıba sokulacağını belirleyen genlerdir. Kodladığı proteinler diğer genlerin ekspresyonunu denetler.Hücre bölünme hızının ve hücrenin yerleşeceği yerin düzenlenmesinde, Hücre şeklinin ve büyüklüğünün belirlenmesinde. Hücre yüzey özelliklerinin (yapışkanlığının) belirlenmesinde.Son farklılaşma ürünlerinin düzenlenmesinde rolleri vardır. Tüm homeotik genler,dublikasyon yoluyla oluşmuşlardır.Bu genler HOMEOBOX (veya HOX) denilen 180 nükleotitlik (60 amino asitlik) ortak bir bölge içerir. Homeobox proteinleri,DNA promotör ve enhancer (kontrol) dizilerine bağlanan düzenleyici protein bölgelerine benzerlik gösterir.Bu dizilere bağlandığında bir seri gende koordine bir ekspresyon sağlarlar.Homeotik mutasyonlar,yetişkindeki bir segmentin diğerine kısmen ya da tümüyle dönüşmesine neden olabilir.
 belirlenmesinde.Son farklılaşma ürünlerinin düzenlenmesinde rolleri vardır. Tüm homeotik genler,dublikasyon yoluyla oluşmuşlardır.Bu genler HOMEOBOX (veya HOX) denilen 180 nükleotitlik (60 amino asitlik) ortak bir bölge içerir. Homeobox proteinleri,DNA promotör ve enhancer (kontrol) dizilerine bağlanan düzenleyici protein bölgelerine benzerlik gösterir.Bu dizilere bağlandığında bir seri gende koordine bir ekspresyon sağlarlar.Homeotik mutasyonlar,yetişkindeki bir segmentin diğerine kısmen ya da tümüyle dönüşmesine neden olabilir.")
26
Zigotik genler segmet oluşumunu tayin eder
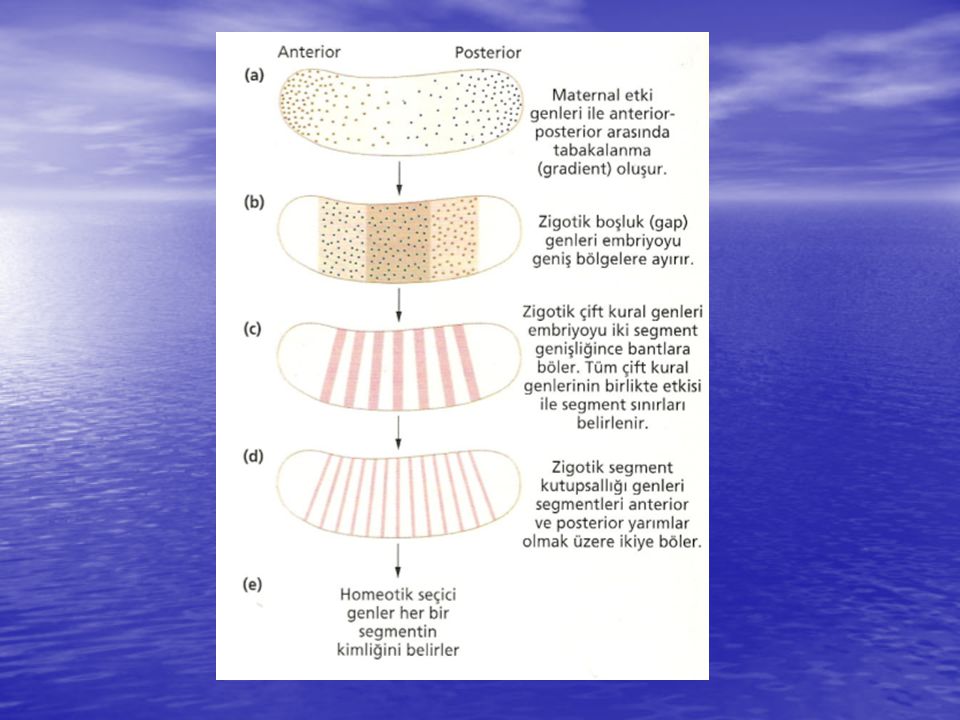
Zigotik genler maternal-etkili gen ürünleri tarafından tabakalanmadaki yerlerine göre aktive olur ya da baskılanır ve bu genlerin üç tanesi; boşluk: gap, çift: kural: pair-rule ve segment polarite: parça kutupsallığı (segment polarity) birlikte segmentasyon genleri olarak adlandırılırlar. Embriyoyu anterior-posterior ekseni boyunca bir seri segmentlere bölerler. Bu segmentasyon genleri gelişen embriyoda transkripsiyona uğrar ve bu genlerdeki mutasyonların etkileri embriyonik ölümcül fenotip olarak belirir.
 birlikte segmentasyon genleri olarak adlandırılırlar. Embriyoyu anterior-posterior ekseni boyunca bir seri segmentlere bölerler. Bu segmentasyon genleri gelişen embriyoda transkripsiyona uğrar ve bu genlerdeki mutasyonların etkileri embriyonik ölümcül fenotip olarak belirir.")
27
Boşluk (Gap) Genleri Gap genlerinin transkripsiyonu, maternal tabakalanma sistemdeki bicoid ve diğer genler dahil olmak üzere anterior-posterior ekseni boyunca daha önce ifade edilmiş gen ürünleri tarafından aktive olur veya baskılanır. Bu genler mutasyona uğrarsa embriyonun segmentasyon profilinde büyük boşluklar görülür. Gap genlerinin transkripsiyonu embriyoyu geniş bölgelere böler (kabaca, baş, göğüs ve karın) ve her bir bölgede farklı gen aktivitesi kombinasyonu, segmentin hem oluşturacağı tipi hem de larvanın, pupanın ve erginin vücut segmentlerinin uygun sırasını belirler. Bu güne kadar, klonlanan tüm gap genleri çinko-parmak (zinc-finger) DNA bağlanma motifindeki transkripsiyon faktörlerini şifreler. Gap genleri çift-kural genlerinin transkripsiyonunu kontrol eder.
 ve her bir bölgede farklı gen aktivitesi kombinasyonu, segmentin hem oluşturacağı tipi hem de larvanın, pupanın ve erginin vücut segmentlerinin uygun sırasını belirler. Bu güne kadar, klonlanan tüm gap genleri çinko-parmak (zinc-finger) DNA bağlanma motifindeki transkripsiyon faktörlerini şifreler. Gap genleri çift-kural genlerinin transkripsiyonunu kontrol eder.")
28
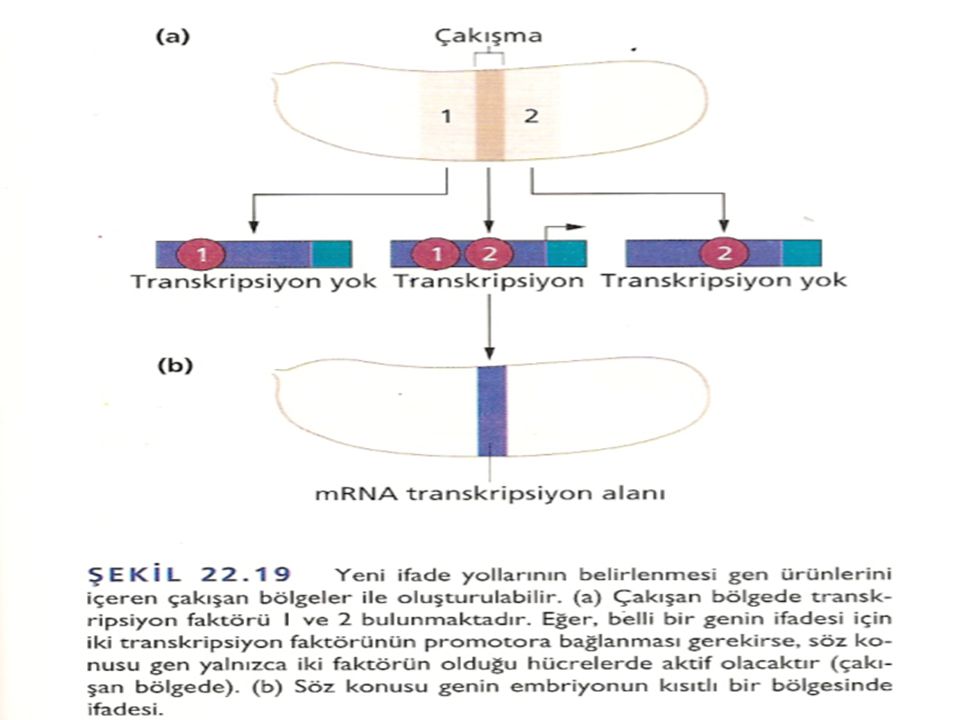
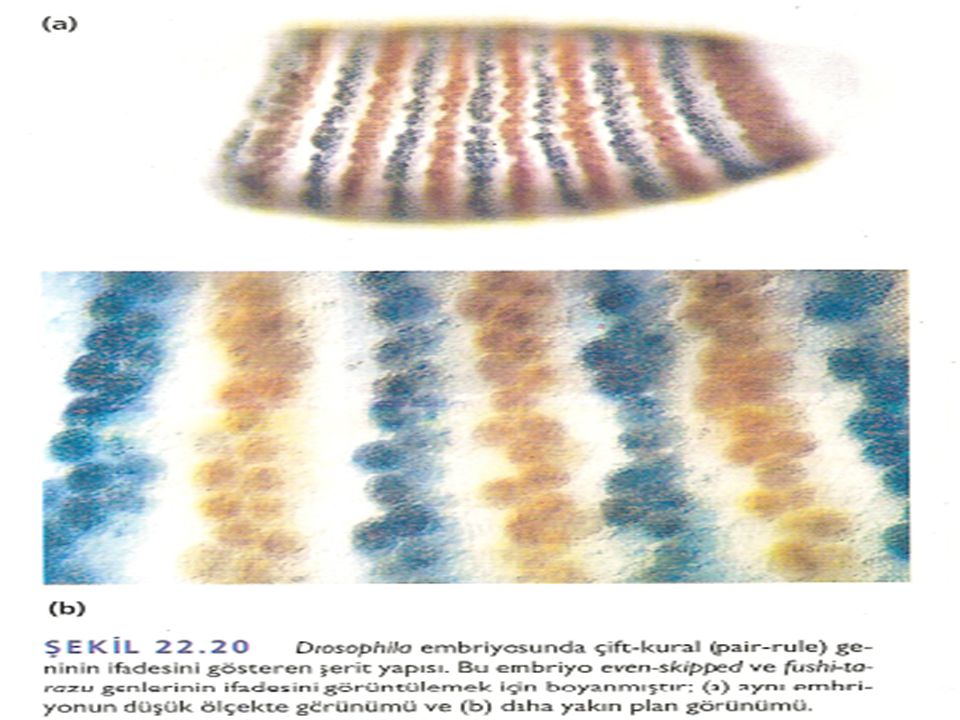
Çift-Kural Genleri Çift-kural (pair-rule) genleri, boşluk (gap) genlerinin oluşturduğu geniş bölgeleri yaklaşık bir segment büyüklüğünde parçalara böler. Çift-kural (pair-rule) genlerindeki mutasyonların, iki segmentten birinde, neredeyse segment boyutunda bölgelerin yapıdan çıkmasına neden olduğu görülmüştür. Çift-kural genleri, çekirdeğin embriyonun çevresine kadar uzanan dar bant veya şeritlerinde ifade olur. Bu gen takımının ifadesi önce segmentlerin sınırını belirler ve sonra segment polarite genlerini kontrol ederek, her bir segmentteki hücrelerin gelişim yolunu belirler. Embriyoyu bir seri şeritlere bölen en az sekiz tane çift-kural geni vardır. Bu şeritlerin sınırlarında çakışan kısımlarının olması, şeritlerdeki hücrelerde çift-kural genlerinin birbiri ile çakışacak şekilde farklı kombinasyonlarda ifade olduklarını gösterir .
 genleri, boşluk (gap) genlerinin oluşturduğu geniş bölgeleri yaklaşık bir segment büyüklüğünde parçalara böler. Çift-kural (pair-rule) genlerindeki mutasyonların, iki segmentten birinde, neredeyse segment boyutunda bölgelerin yapıdan çıkmasına neden olduğu görülmüştür. Çift-kural genleri, çekirdeğin embriyonun çevresine kadar uzanan dar bant veya şeritlerinde ifade olur. Bu gen takımının ifadesi önce segmentlerin sınırını belirler ve sonra segment polarite genlerini kontrol ederek, her bir segmentteki hücrelerin gelişim yolunu belirler. Embriyoyu bir seri şeritlere bölen en az sekiz tane çift-kural geni vardır. Bu şeritlerin sınırlarında çakışan kısımlarının olması, şeritlerdeki hücrelerde çift-kural genlerinin birbiri ile çakışacak şekilde farklı kombinasyonlarda ifade olduklarını gösterir .")
31
Segment Polarite Genleri
Segment polarite genlerinin ifadesi çift-kural (pair-rule) genlerinin sentezlediği transkripsiyon faktörleri tarafından kontrol edilir. Segment polarite genleri, embriyonun etrafını bir şerit gibi saran segment hücrelerinde aktif hale gelir. Bu ifadenin oluşturduğu bant profili embriyoyu 14 kısma ayırır ve gen ürünleri segment içindeki hücresel kimliği kontrol eder. Segment polarite genlerinin bazıları, engrailed geni dahil, transkripsiyon faktörleri sentezler. Engrailed proteini transkripsiyonu aktive etmekten ziyade diğer home-odomain proteinlerinin aktivitelerini onlarla rekabete girerek engeller ve sonuçta segment sınırını oluşturur.
 genlerinin sentezlediği transkripsiyon faktörleri tarafından kontrol edilir. Segment polarite genleri, embriyonun etrafını bir şerit gibi saran segment hücrelerinde aktif hale gelir. Bu ifadenin oluşturduğu bant profili embriyoyu 14 kısma ayırır ve gen ürünleri segment içindeki hücresel kimliği kontrol eder. Segment polarite genlerinin bazıları, engrailed geni dahil, transkripsiyon faktörleri sentezler. Engrailed proteini transkripsiyonu aktive etmekten ziyade diğer home-odomain proteinlerinin aktivitelerini onlarla rekabete girerek engeller ve sonuçta segment sınırını oluşturur.")
32
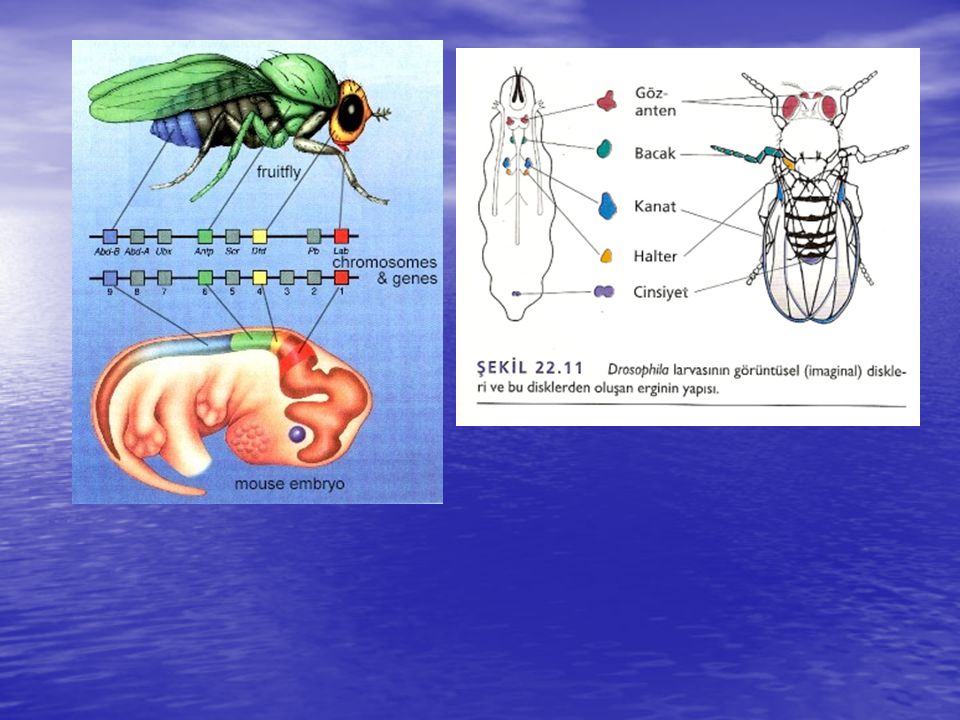
Seçici Genler Segmentlerin sınırları, segmentasyon genlerinin etkisiyle oluşturulurken seçici genler (selektör) aktive olur. Bu genlerin aktivitesi ile anten, ağız kısmı, bacaklar, kanatlar, göğüs ve karın gibi her bir vücut segmentinin oluşacağı yapı belirlenir. Bu genlerin mutandan homeotik mutantlar (Yunanca "aynı" kelimesinden gelmektedir) olarak bilinir, çünkü, segmentlerden birinin özelliği, komşusegmentin özelliği ile aynı olacak şekilde değişime uğrar. Örneğin Antennapedia (Antp) geninin yabanıl tip alleli ikinci göğüs (toraks) segmentindeki (bir bacak taşır) yapıların özelleşmesi için gereklidir. Baskın Antp mutasyonunun oluşması baş ve göğüsteki genin ifadesini engelleyerek anteni mutant olan sineklerin oluşumuna neden olur ve sonuçta anten oluşacak olan segmentte bacaklar meydana gelir .
 aktive olur. Bu genlerin aktivitesi ile anten, ağız kısmı, bacaklar, kanatlar, göğüs ve karın gibi her bir vücut segmentinin oluşacağı yapı belirlenir. Bu genlerin mutandan homeotik mutantlar (Yunanca aynı kelimesinden gelmektedir) olarak bilinir, çünkü, segmentlerden birinin özelliği, komşusegmentin özelliği ile aynı olacak şekilde değişime uğrar. Örneğin Antennapedia (Antp) geninin yabanıl tip alleli ikinci göğüs (toraks) segmentindeki (bir bacak taşır) yapıların özelleşmesi için gereklidir. Baskın Antp mutasyonunun oluşması baş ve göğüsteki genin ifadesini engelleyerek anteni mutant olan sineklerin oluşumuna neden olur ve sonuçta anten oluşacak olan segmentte bacaklar meydana gelir .")
33
Seçici genlerin aktivasyonu boşluk (gap) genleri ve çift-kural (pair-rule) genlerinin kontrolü altındadır. Seçici gen ifadesi embriyoda belirli bölgelerde görülmesine rağmen, ifadenin zamanlaması ve profili karmaşıktır ve segmentasyon ile seçici genler arasında olduğu gibi aynı zamanda diğer çeşidi seçici genler arasındaki etkileşimler ile ilişkilidir.

34
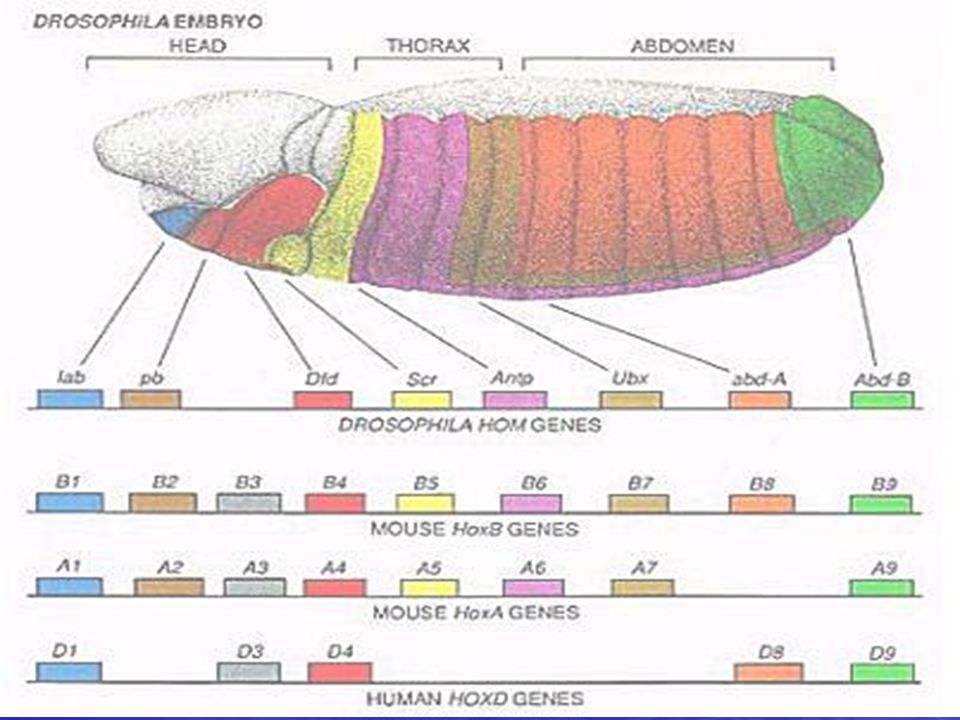
Segmentasyona katılan bitoraks kompleksindeki genlerin ve diğer genlerin, peş peşe duplikasyon sonucu, ortak bir atadan oluştuğunu ve birbirlerinden yapısal ve işlevsel olarak giderek uzaklaştığını ileri sürmüştür. Homeotik seçici genlerinin her biri transkripsiyon faktörlerini kodlar. Bu faktörler homeo kutusu (homeobox) olarak bilinen 180-bç'lik bir dizinin şifrelediği DNA-bağlanma bölgesi içerir. Homeobox, homeodomain olarak bilinen 60 amino asitlik bir bölgeyi kodlar. Benzer diziler, Kenopîts, tavuk, fare ve insanlar dahil çeşitli ökar-yotların genomlarında da bulunmuştur. Tüm memelilerde seçici gen yapıları Hox gen kümeleri olarak isimlendirilir. Bu güne kadar incelenen tüm organizmalara ait homeodomainlerin benzer amino asit dizileri vardır ve özgül gen takımlarının transkripsiyonal kontrolü ile ilgili proteinleri kodlarlar. Bu sonuç metamerik veya segmentli vücut yapısının sadece bir kez evrimleşnıiş olabileceği fikrini vermektedir.
 olarak bilinen 180-bç lik bir dizinin şifrelediği DNA-bağlanma bölgesi içerir. Homeobox, homeodomain olarak bilinen 60 amino asitlik bir bölgeyi kodlar. Benzer diziler, Kenopîts, tavuk, fare ve insanlar dahil çeşitli ökar-yotların genomlarında da bulunmuştur. Tüm memelilerde seçici gen yapıları Hox gen kümeleri olarak isimlendirilir. Bu güne kadar incelenen tüm organizmalara ait homeodomainlerin benzer amino asit dizileri vardır ve özgül gen takımlarının transkripsiyonal kontrolü ile ilgili proteinleri kodlarlar. Bu sonuç metamerik veya segmentli vücut yapısının sadece bir kez evrimleşnıiş olabileceği fikrini vermektedir..")
35
Bu üç sınıftaki pek çok regülatör gen, belirli sayıda segmentleri ve her segmentte doğru ilaveleri bulunan, bir başı, toraksı ve abdomeni olan bir meyve sineğinin oluşmasını yönetir. Embriyogenezisin tamamlanması yaklaşık bir gün alsa da, tüm bu genler ilk 4 saatte aktif olurlar. Bazı mRNA'lar ve proteinler bu periyod içinde yalnız bir kaç dakika ve özgül noktalarda bulunurlar. Genlerden bazıları, gelişim zincirindeki diğer bazı genlerin ekspresyonlarını etkileyen transkripsiyon faktötrlerini kodlar. Ayrıca, translasyon seviyesinde regülasyon da önemli yer tutar; çoğu regülatör gen, translasyonel repressörleri kodlar. Translasyonel regülatörlerin çoğu, mRNA'nın 3' UTR'sine bağlanır. Çoğu mRNA, translasyonunun gerektiği zamandan önce yumurta hücresinde depolandığı için, translasyonel represyon gelişimsel yollarda özellikle önemlidir.

36
What is a homeobox? "Since their discovery in 1983, homeobox genes, and the proteins they encode, the homeodomain proteins, have turned out to play important roles in the developmental processes of many multicellular organisms. While certainly not the only developmental control genes, they have been shown to play crucial roles from the earliest steps in embryogenesis - such as setting up an anterior-posterior gradient in the egg of the fruit fly Drosophila melanogaster - to the very latest steps in cell differentiation - such as the differentiation of neurons in the nematode Caenorhabditis elegans (C. elegans). They have a wide phylogenetic distribution, having been found in baker's yeast, plants, and all animal phyla that have been examined so far. Since their original discovery, hundreds of homeobox genes have been described" 1983 te keşiflerinden beri, homeobox genleri ve onların kodladığı proteinler yani homeodomain proteinleri, pek çok multicellular organismanın oluşum sürecinde önemli roller oynamaktadırlar. Aynı zamanda, sadece control genlerinin gelişiminde değil, embriyo genlerinin en erken safhalarından itibaren, örneğin; meyva sineğinin yumurtasında anterior-posterior oluşumunda… hücre farklılaşmasında en son safhalara kadar, örneğin; Caenorhabditis (iplikkurdu)nun sinir hücrelerinin farklılaşmasında… önemli rol oynamaktadırlar. Homeobox genler geniş bir fologenetic dağılıma sahipler; hamur mayasında, bitkilerde, bu güne kadar araştırılan hayvan organellerinde bulunmuşturlar. Orijinal keşiflerinden itibaren, yüzlerce homeobox geni tanımlanmıştır.
. They have a wide phylogenetic distribution, having been found in baker s yeast, plants, and all animal phyla that have been examined so far. Since their original discovery, hundreds of homeobox genes have been described 1983 te keşiflerinden beri, homeobox genleri ve onların kodladığı proteinler yani homeodomain proteinleri, pek çok multicellular organismanın oluşum sürecinde önemli roller oynamaktadırlar. Aynı zamanda, sadece control genlerinin gelişiminde değil, embriyo genlerinin en erken safhalarından itibaren, örneğin; meyva sineğinin yumurtasında anterior-posterior oluşumunda… hücre farklılaşmasında en son safhalara kadar, örneğin; Caenorhabditis (iplikkurdu)nun sinir hücrelerinin farklılaşmasında… önemli rol oynamaktadırlar. Homeobox genler geniş bir fologenetic dağılıma sahipler; hamur mayasında, bitkilerde, bu güne kadar araştırılan hayvan organellerinde bulunmuşturlar. Orijinal keşiflerinden itibaren, yüzlerce homeobox geni tanımlanmıştır.")
38
Homeobox Genes Transcription factors encoding the DNA binding homeodomain. Expressed in temporal and spatial specific patterns during development. Cell differentiation is disrupted when they are mutated. Transkripsiyon faktörler homeodomain DNA bağlantısını kodlar. Gelişim esnasında spesifik modelin geçici ve o anki durumunu ifade eder. Homeobox genleri mutasyona uğradığında, hücre farklılaşması oluşur. A transcription factor is a protein that binds to DNA and plays a role in the regulation of gene expression by promoting transcription. Homeobox: A gene containing a 180-base-pair segment (the “homeobox”) that encodes a protein domain involved in binding to (and thus regulating the expression of) DNA. This homeobox is very similar in many genes with different functions. Homeo box is highly conservative gene fragment coding peptide with high affinity to some DNA sequences. It play a key role in regulation of gene expression. The proteins encoded by such Hox genes are possible candidate as transcription factors believed to be involved in given positional cues to different cell types along different rudiments and body axes. First identified in a series of Drosophila genes that regulate development, the homeodomain is a conserved sequence of 60 amino acids. Homeodomain-containing proteins have now been found in all eukariotes examined, ranging from yeast to man. The tertiary structure of the homeo domain, as first determined by solution NMR studies of the Antennapedia homeodomain from Drosophila, consists of three alfa-helices (1, 2, 3) and a flexible N-terminal arm. Lbx is a specific homeobox gene having important functions. Homeobox genes induce expression patterns specific to time and also to space.
 that encodes a protein domain involved in binding to (and thus regulating the expression of) DNA. This homeobox is very similar in many genes with different functions. Homeo box is highly conservative gene fragment coding peptide with high affinity to some DNA sequences. It play a key role in regulation of gene expression. The proteins encoded by such Hox genes are possible candidate as transcription factors believed to be involved in given positional cues to different cell types along different rudiments and body axes. First identified in a series of Drosophila genes that regulate development, the homeodomain is a conserved sequence of 60 amino acids. Homeodomain-containing proteins have now been found in all eukariotes examined, ranging from yeast to man. The tertiary structure of the homeo domain, as first determined by solution NMR studies of the Antennapedia homeodomain from Drosophila, consists of three alfa-helices (1, 2, 3) and a flexible N-terminal arm. Lbx is a specific homeobox gene having important functions. Homeobox genes induce expression patterns specific to time and also to space.")
39
HOX Geni ? Bu genler, tüm hayvanlarda, yapı ve işlev bakımından çarpıcı şekilde benzerlik gösteriyor. Bu nedenle de, hayvanların vücutlarının belirli planlara göre oluşumunun mimarları olarak kabul ediliyorlar. Yaklaşık olarak 60 amino asitlik bir diziyi kodlayan Hox genleri, DNA üzerindeki belirli gen şifrelerinin okunuşunu etkiliyor. Yani, hayvan hücrelerinin gelişimi ve kaderleri (cell fate), Hox genleri tarafından kontrol ediliyor. Örneğin insandaki ve farelerdeki Hox genleri, sayı ve kromozom düzeni bakımından büyük benzerlik gösteriyor. Gelişmiş memelilerdeki vücut planlarının mimarisi, görünüşü ve gelişimi, yaklaşık 40 kadar Hox geniyle kontrol ediliyor. Belirli hayvan gruplarında ise, Hox genleri, birbirinden farklı ancak birbiriyle yakın kabul edilebilecek özelliklerin ortaya çıkışını kontrol ediyor. Örneğin omurgasızlarda karın kısmının gelişiminden sorumlu olan Hox genleri, omurgalıların sırt kısmının gelişiminden sorumlu. Kısacası bu genler, söz konusu vücut planlarının ortaya çıkmasında en büyük rolü oynayan moleküler mimarlar.
, Hox genleri tarafından kontrol ediliyor. Örneğin insandaki ve farelerdeki Hox genleri, sayı ve kromozom düzeni bakımından büyük benzerlik gösteriyor. Gelişmiş memelilerdeki vücut planlarının mimarisi, görünüşü ve gelişimi, yaklaşık 40 kadar Hox geniyle kontrol ediliyor. Belirli hayvan gruplarında ise, Hox genleri, birbirinden farklı ancak birbiriyle yakın kabul edilebilecek özelliklerin ortaya çıkışını kontrol ediyor. Örneğin omurgasızlarda karın kısmının gelişiminden sorumlu olan Hox genleri, omurgalıların sırt kısmının gelişiminden sorumlu. Kısacası bu genler, söz konusu vücut planlarının ortaya çıkmasında en büyük rolü oynayan moleküler mimarlar.")
40
Hox genes: Hox genes are a subgroup of homeobox genes
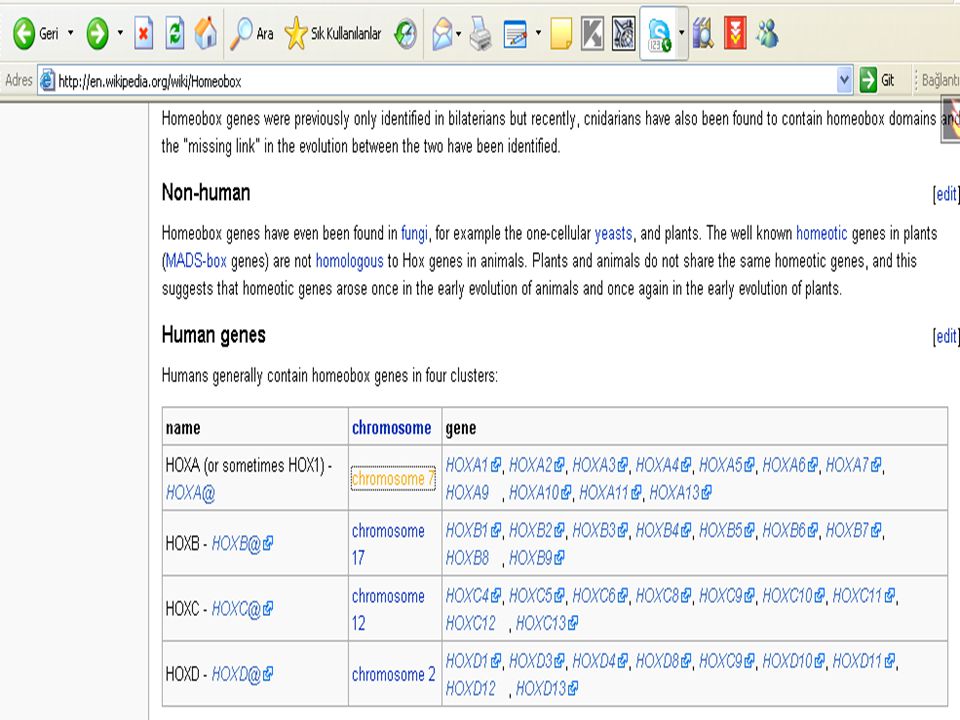
Hox genes: Hox genes are a subgroup of homeobox genes. In vertebrates these genes are found in gene clusters on the chromosomes. In mammals four such clusters exist, called Hox clusters. The gene name "Hox" has been restricted to name Hox cluster genes in vertebrates. Only genes in the HOX cluster should be named Hox genes. So note: homeobox genes are NOT Hox genes, Hox genes are a subset of homeobox genes. Hox genleri: Hox genleri, homeobox genlerin bir alt grubudur. Omurgalı hayvanlarda, bu genler kromozomlarda, gen kümelerinde bulunmaktadır. Memelilerde, Hox sınıfları olarak adlandırılan, bu şekilde dört küme vardır. Bu Hox geni, omugalılarda, Hox kümesi genlerine bağlı olarak isimlendirilmiştir. Sadece Hox kümesindeki genler, Hox geni olarak isimlendirilmelidir. Bundan dolayı, homeobox genleri Hox genleri değildir. Çünkü, Hox genleri homeobox genlerin bir alt kümesidir.

41
HOX cluster: The term Hox cluster refers to a group of clustered homeobox genes, named Hox genes in vertebrates, that play important roles in pattern formation along the anterior-posterior body axis. In fact, the first homeobox genes discovered where those of the Drosophila homeotic gene clusters, i.e. the "Antennapedia complex" and the "Bithorax complex", which summarily are referred to as HOM-C (homeotic complex). This HOM-C complex in Drosophila is the evolutionary homolog of the vertebrate Hox clusters and the evolutionarily related homeobox gene clusters in other animals (i.e. chordates, insects, nematodes, etc.) are now also called HOX clusters. Hox kümesi: Hox kümesi terimi, omurgalılarda, Hox genleri olarak adlandırılan, kümelenmiş homeobox genlerin bir grubunu belirtmektedir ve bu genler omuiliğin anterior-posterior oluşumu boyunca önemli rol oynar. İlk homeobox genleri, Drosophila (meyva sineği) homeotic gen kümelerinde keşfedildi; yani, Antennapedia kompleksi ve Bithorax kompleksi, özet olarak, HOM-C (homeotic kompleksi) olarak belirtilmektedir. Meyva sineğindeki bu HOM-C kompleksi, omurgalıların Hox kümelerinin evrimsel homoloğudur ve böcekler gibi diğer hayvanlarda, bugünlerde, Hox kümeleri olarak da adlandırılan, homeobox gen kümelerine ilişkin evrimselliğidir. homeodomain: a DNA-binding domain, usually about 60 amino acids in length, encoded by the homeobox. Homeodomain: Homeobox tarafından kodlanan, genellikle 60 amino asit uzunluğunda, bir DNA-bağlayıcısı alanı/bölgesi.
 homeotic gen kümelerinde keşfedildi; yani, Antennapedia kompleksi ve Bithorax kompleksi, özet olarak, HOM-C (homeotic kompleksi) olarak belirtilmektedir. Meyva sineğindeki bu HOM-C kompleksi, omurgalıların Hox kümelerinin evrimsel homoloğudur ve böcekler gibi diğer hayvanlarda, bugünlerde, Hox kümeleri olarak da adlandırılan, homeobox gen kümelerine ilişkin evrimselliğidir. homeodomain: a DNA-binding domain, usually about 60 amino acids in length, encoded by the homeobox. Homeodomain: Homeobox tarafından kodlanan, genellikle 60 amino asit uzunluğunda, bir DNA-bağlayıcısı alanı/bölgesi.")
43
Utah araştırmacılarının yeniden yapılandırdığı ‘Hox’ geni, embriyonun gelişiminde diğer genlerin hareketlerini düzenlemekle görevli. İlk hayvanlar, 500 milyon yıl önce 13 adet Hox geninden oluşuyordu, bunların her biri dörde bölününce ortaya 52 adet gen çıktı. Bölünmeyle ortaya çıkan genlerin bazıları zamanla çoğaldı, bazıları ise yokoldu. Bugün insanlarda ve memelilerde 39 adet Hox geni bulunuyor. Bugün insanlar ve memelilerde orijinal Hox geninin torunları sayılan Hoxa1 ve Hoxb1 genleri var.

44
HOXA1 ve HOXB1 HOXA1 VE HOXB1 GENLERİ Hoxa1, solumayla ilgili önemli bir gen. Fare embriyonunda Hoxa1 geni bozulduğunda fare doğumdan kısa bir süre sonra ölüyor. Hoxb1 ise, sinir hücrelerinin ve yüz ifade kaslarının formasyonundan sorumlu. Hoxb1 geni embriyo farelerde iptal edildiğinde, fare ileri derece yüz felci oluyor ve gözlerini kırpamaz, kaşlarını oynatamaz hale geliyor. Araştırmacılar Hoxa1 ve Hoxb1 genlerini temel alarak, orijinal Hox1 genini yeniden yarattılar. Orijinal gen olan Hox1, torunları Hoxa1 ve Hoxb1 genlerinin her ikisinin de işlevlerini yerine getirebiliyor. Hox1 geniyle doğan bir fare, hem solunum yapabiliyor hem de yüz kaslarını idare edebiliyor. Bu gen çıkarıldığında da bu iki işlev kayboluyor.

45
YENİ GEN ORİJİNALİNİN KOPYASI
Yeniden yaratılan gen, aslında orijinal genin torunlarının bir hibridi, ancak 530 milyon yıl önce yokolmuş genin aynısı değil. Ancak orijinal genin bir kopyası ve aynı işlevleri yerine getirebiliyor. Yeniden yapılandırılan Hox1 geninde, yine orijinal genin torunları olan Hoxc1 ve Hoxd1 genleri bulunmuyor, zira bu iki genin, büyük ihtimalle mükerrer oldukları için evrim sürecinde yok olduğu düşünülüyor. Araştırma, gen tedavilerinde bozulan bir geni, kendisine akraba bir başka genle birleştirerek onarmanın yolunu açacak.

Benzer bir sunumlar















>")




