Sunuyu indir
30
E N Z İ M L E R

31
Enzimler, metabolizma reaksiyonlarını hızlandıran,
I.TANIM Enzimler, metabolizma reaksiyonlarını hızlandıran, biyolojik katalizörlerdir. Protein yapısında maddelerdir. Hücrenin gereksinimine uygun kataliz yaparlar. Hücre içindeki yerleşimleri metabolik olayların özelliğine göre düzenlenmiştir.

32
III.ENZİMLERİN İSİMLENDİRİLMESİ
Etkiledikleri maddelerin (substratlarının) sonuna -az- takısı eklenerek isimlendirilir. Madde Hidroliz Enzimi Nişasta(Amilon) Amilaz Yağ (Lipos) Lipaz Protein Proteaz Üre Üreaz Bu kurala uymayanlar: Pepsin, tripsin, pityalin Katalizledikleri reaksiyon tipine göre: Oksidaz, dekarboksilaz
 sonuna -az- takısı eklenerek. isimlendirilir. Madde Hidroliz Enzimi. Nişasta(Amilon) Amilaz. Yağ (Lipos) Lipaz. Protein Proteaz. Üre Üreaz. Bu kurala uymayanlar: Pepsin, tripsin, pityalin. Katalizledikleri reaksiyon tipine göre: Oksidaz, dekarboksilaz.")
33
Günümüzde bu tür karışıklıkları önlemek amacıyla Uluslararası Biyokimya
Birliğinin (IUB) Enzim Komisyonu tarafından Sistematik isimlendirme önerilmiştir. Bu sistemde her enzim, katalizlediği reaksiyon tipine ve mekanizmasına göre isimlendirilmektedir. Günlük kullanımda önerilen kısa isme ilaveten daha detaylı sistematik ismin kullanılması öngörülmüştür. SİSTEMATİK İSİMLENDİRMENİN TEMEL ÖZELLİKLERİ 1. Reaksiyonlar ve bu reaksiyonları katalizleyen enzimler, reaksiyon mekanizmalarına göre 6 sınıfa bölünürler. Bu sınıflarında alt sınıfları vardır. 2. Her enzimin bir kod numarası vardır (EC). Bu kod, dörtlü sayı grubu ile gösterilir.
 Enzim Komisyonu tarafından Sistematik isimlendirme. önerilmiştir. Bu sistemde her enzim, katalizlediği reaksiyon tipine ve. mekanizmasına göre isimlendirilmektedir. Günlük kullanımda önerilen kısa isme ilaveten daha detaylı sistematik. ismin kullanılması öngörülmüştür. SİSTEMATİK İSİMLENDİRMENİN TEMEL ÖZELLİKLERİ. 1. Reaksiyonlar ve bu reaksiyonları katalizleyen enzimler, reaksiyon. mekanizmalarına göre 6 sınıfa bölünürler. Bu sınıflarında alt sınıfları. vardır. 2. Her enzimin bir kod numarası vardır (EC). Bu kod, dörtlü sayı grubu. ile gösterilir.")
36
Sistematik İsimlendirme: ATP + D-Glukoz ADP + D-Glukoz-6-Fosfat
Önerilen kısa isim: Hekzokinaz Enzim kodu : EC ( ) Sistematik isim : ATP: glukoz fosfotransferaz EC ( ): İlk sayı: Reaksiyon tipini açıklar (Major sınıf) Transferaz sınıfı İkinci sayı: Alt sınıf Fosfotransferaz Üçüncü sayı:Alt alt sınıf Hidroksil grubunun alıcı olduğu fosfotransferaz Dördüncü sayı:Enzim için spesifiktir. Fosfat grubunun D-glukoza aktarıldığını açıklar. Enzimin listeye girdiği seri numarasıdır. Hekzokinaz
 Sistematik isim : ATP: glukoz fosfotransferaz. EC ( ): İlk sayı: Reaksiyon tipini açıklar (Major sınıf) Transferaz sınıfı. İkinci sayı: Alt sınıf. Fosfotransferaz. Üçüncü sayı:Alt alt sınıf. Hidroksil grubunun alıcı olduğu fosfotransferaz. Dördüncü sayı:Enzim için spesifiktir. Fosfat grubunun D-glukoza. aktarıldığını açıklar. Enzimin listeye girdiği seri numarasıdır. Hekzokinaz.")
37
IV.ENZİMLERİN ÖZELLİKLERİ
1. Enzimler protein yapısında maddelerdir: Protein yapısına istisna olarak bazı RNA tipleri gösterilebilir. Bunlar, fosfodiester bağlarının yıkımı ve sentezi esnasında enzim gibi davranabilirler. Katalitik etkiye sahip RNA’ya Ribozim denir. 2.Enzimler özgül moleküllerdir. Enzimler yalnız belirli reaksiyonları katalizledikleri ve ortamdaki moleküllerden sadece substratları ile etkileştiklerinden dolayı spesifik (özgül) moleküllerdir. 3.Enzimler katalitik etkinliğe sahiptir. Enzimle katalizlenen reaksiyonların çoğu katalizlenmeyen reaksiyonlara göre 103 –108 kere daha hızlı olarak gerçekleşmektedir. Bir enzim molekülü saniyede ortalama substrat molekülünün ürüne dönüşümünü sağlamaktadır. 4. Enzimler görev yerlerine göre kompartmanlanmıştır. Her enzim farklı orgenelde görev alabilir.
 moleküllerdir. 3.Enzimler katalitik etkinliğe sahiptir. Enzimle katalizlenen reaksiyonların çoğu katalizlenmeyen. reaksiyonlara göre 103 –108 kere daha hızlı olarak gerçekleşmektedir. Bir enzim molekülü saniyede ortalama substrat. molekülünün ürüne dönüşümünü sağlamaktadır. 4. Enzimler görev yerlerine göre kompartmanlanmıştır. Her enzim farklı orgenelde görev alabilir.")
38
4. Substrat-ürün dönüşümleri çift yönlü olabilmektedir.
Enzimin dönüşüm sayısı (Turnover sayısı): Enzim molekülü tarafından bir saniyede ürüne çevrilen substrat molekülü sayısıdır. CO2 + H2O H2CO3 Karbonik anhidraz, 1 saniyede 105 molekül CO2’e, H2O’yu bağlayarak karbonik asid oluşturur. 4. Substrat-ürün dönüşümleri çift yönlü olabilmektedir. C Karbonik anhidraz H O Trioz fosfat izomeraz CH2OH O H C C OH CH2OPO3-2 CH2OPO3-2 Dihidroksi aseton fosfat (DHAP) Gliseraldehid-3-fosfat (G3P)
: Enzim molekülü tarafından bir saniyede ürüne çevrilen substrat. molekülü sayısıdır. CO2 + H2O H2CO3. Karbonik anhidraz, 1 saniyede 105 molekül CO2’e, H2O’yu bağlayarak. karbonik asid oluşturur. 4. Substrat-ürün dönüşümleri çift yönlü olabilmektedir. C. Karbonik anhidraz. H. O. Trioz fosfat izomeraz. CH2OH. O. H. C. C. OH. CH2OPO3-2. CH2OPO3-2. Dihidroksi aseton fosfat (DHAP) Gliseraldehid-3-fosfat (G3P)")
39
Bu iki madde arasındaki izomerizasyon glikoliz yolunda rastlanır.
Enzim iki yöne doğru reaksiyon hızını arttırmaktadır. 5.Enzim moleküllerinde aktif bölge ismi verilen özel bir boşluk ya da cep kısmı bulunur. Aktif bölgedeki aminoasidlerin yan zincirleri, substratın yapısına uyumlu, üç boyutlu bir yapı oluşturmaktadır. Enzim-Substrat kompleksi Substrat Enzim Aktif bölge

40
Aktif bölgenin substratı bağlamasıyla oluşan enzim-substrat
kompleksi (ES), önce enzim-ürün kompleksine, daha sonra ise serbest enzim ve ürüne dönüşmektedir. E + S ES EÜ E + Ü Enzim ile substrat biribirlerine hidrojen, elektrostatik ve Van der Waals bağları gibi non kovalent (zayıf) bağlarla bağlanır. Zayıf bağlar ve bazı kuvvetli bağlar, aktif bölgelerin aminoasidlerini biribirlerine yanaştırmada önemli rol oynarlar.
, önce enzim-ürün kompleksine, daha sonra ise. serbest enzim ve ürüne dönüşmektedir. E + S ES EÜ E + Ü. Enzim ile substrat biribirlerine hidrojen, elektrostatik ve Van der. Waals bağları gibi non kovalent (zayıf) bağlarla bağlanır. Zayıf bağlar ve bazı kuvvetli bağlar, aktif bölgelerin aminoasidlerini. biribirlerine yanaştırmada önemli rol oynarlar.")
41
Birçok enzimin katalitik bölgesinde aşağıdaki aminoasidler yer alır:
Serin, sistein, histidin, tirozin ve lizin S Aktif Bölge S Bağlanma bölgesi Katalitik bölge S S 119 His Katalitik bölge S His 12 S S S Bağlanma bölgesi RİBONÜKLEAZ

42
Enzimler kolaylıkla denatürasyona uğrarlar:
Denatürasyon, proteinlerin doğal yapılarının bozulması sonucunda aktivitelerinin kaybolmasıdır. Enzim denatüre olduğunda aktif bölgesi de denatürasyona uğrayarak substratını bağlayamaz, bundan dolayı da etkili olamaz. Başlıca denatürasyona yol açan faktörler: Isı Işınlar (X ışınları, UV ışınları, vs) Çalkalama Dondurup eritme Derişik asid ve baz Alkol, eter, benzen, vs gibi organik çözücüler Üre, guanidin çözeltileri
 Çalkalama. Dondurup eritme. Derişik asid ve baz. Alkol, eter, benzen, vs gibi organik çözücüler. Üre, guanidin çözeltileri.")
43
RİBONÜKLEAZ: Bu enzim RNA molekülündeki nukleotidleri hidroliz yapar. Yapısı 4 disülfür bağı ile sağlamlaşmıştır. Katalitik bölgede 2 histidin kalıntısı yer alır. Histidin 12 ve Histidin 119 His 12, ribozun hidroksil grubu üzerine etki eder. His 119, ise fosforil kısmına etkilidir. Böylece molekülün 2 kısmından kırılma gerçekleşir. Molekülün taranmış kısmı ise 5 aminoasidin yer aldığı bazik bir bölgedir. Bu bölge RNA‘yı bağlar.

44
Enzimlerin substrat bağlama yeri olan aktif bölgedeki aminoasidler,
substratın ürüne dönüşmesini sağlayan pek çok kimyasal mekanizmayı kullanır. Bu aminoasidlerden bazıları substratın aktif merkeze bağlanmasını, bazıları ise kataliz olayını sağlamaktadır. Aktif merkezde yer alan iki bölgeden birincisi bağlanma bölgesi, diğeri ise katalitik bölgeyi oluşturur.

45
Enzim ile substrat bağlanmasında iki model ileri sürülmektedir.
1.Model: Anahtar-kilit modeli 2.Model: Katalitik bölgenin “uyum oluşturma modeli”

46
Kilit anahtara olan benzerliğe dayanılmıştır.
1.MODEL: Kilit anahtara olan benzerliğe dayanılmıştır. Bu modelde enzimin aktif merkezindeki bir bölge ile substrat yapılarının biribirini tamamlayıcı olmaları gerekmektedir. a c b Substrat + a b c ES kompleksi c a b Enzim Kilit-anahtar modeli

47
+ 2. MODEL: Katalitik bölgenin “Uyum-oluşturma” modelidir.
Başlangıçta enzim ve substrat biribirlerine uygun değildirler. Ancak substrat enzimin aktif bölgesine yaklaştıkça, enzim buna uymaktadır. Substrat, enzimde biçimsel değişiklik meydana getirir. Substrat a c b + a c b a c b ES kompleksi Enzim

48
6.Bazı enzimler, enzimatik reaksiyon için gerekli olan bir non-protein
kofaktör ile birleşirler. Sıklıkla karşılaşılan kofaktörler arasında metal iyonları (Zn2+,Fe2+,Cu2+, Mn+2 …vs) ve koenzim olarak adlandırılan bir organik molekül, genellikle vitamin türevleri (NAD+, FAD,CoA..gibi) yer alır. Koenzimlerin pek çoğu genellikle B grubu vitaminlerden türevlenmektedir. Kofaktörle birleşik durumda olan ve katalitik aktivite gösteren enzim holoenzim olarak bilinmektedir. Holoenzimin protein kısmına apoenzim adı verilir. Apoenzim kofaktörün yokluğunda biyolojik aktivite gösteremez. Protein kısmına sıkıca bağlı olan koenzime prostetik grup denir.
 ve koenzim olarak adlandırılan bir organik molekül, genellikle. vitamin türevleri (NAD+, FAD,CoA..gibi) yer alır. Koenzimlerin pek çoğu genellikle B grubu vitaminlerden. türevlenmektedir. Kofaktörle birleşik durumda olan ve katalitik aktivite gösteren. enzim holoenzim olarak bilinmektedir. Holoenzimin protein kısmına apoenzim adı verilir. Apoenzim kofaktörün yokluğunda biyolojik aktivite gösteremez. Protein kısmına sıkıca bağlı olan koenzime prostetik grup denir.")
49
Kofaktörü metal iyonu olan bazı metalloenzimler:
Kofaktör Enzim Fe Katalaz, peroksidaz Cu Sitokrom oksidaz, tirozinaz Mg Fosfohidrolaz, fosfotransferaz Mn Arginaz Zn Alkol dehidrogenaz Mo Ksantin oksidaz

50
Metal iyonları substrat bağlanmasını ve katalizi kolaylaştırır.
Aşağıdaki 4 form da, metal iyonları ile katalizlenen enzimatik reaksiyonlar için geçerlidir. Enz S M M Enz S Enz M S Enz Substrat-köprü kompleksi Enzim-köprü kompleksi M S Metal-köprü kompleksi Siklik metal-köprü kompleksi

51
Enzim Vitamin Koenzim Katalizlenen reaksiyon
KOENZİMİ VİTAMİN OLAN BAZI ENZİMLER: Enzim Vitamin Koenzim Katalizlenen reaksiyon Dekarboksilaz B1 vitamini TPP (Tiamin pirofosfat) ………………...R-CO-COOH (Tiamin) RCHO + CO2 (Dekarboksilasyon) Dehidrogenaz B2 vitamini FMN(Flavin mononukleotid ve…………..Hidrojen (Riboflavin) FAD Flavin adenin dinükleotid) transferi Transaminaz B6 vitamini Pridoksal fosfat ……………….Amino asidlerden aldığı (Pridoksal) NH2 grubunu α-keto asidlere transfer eder. Karboksilaz Biotin Biotin …………………………α-keto asidlere CO2 ‘i bağlar(Karboksilasyon) Transformilaz Folik asid THF…………………………… -CHO, -CH2 OH, -CH3 (Tetrahidrofolat) gruplarının transferi Transmetilaz B12 vitamini Kobamid…………………………-CH3 grubu transferi İzomeraz koenzim
 ………………...R-CO-COOH. (Tiamin) RCHO + CO2. (Dekarboksilasyon) Dehidrogenaz B2 vitamini FMN(Flavin mononukleotid ve…………..Hidrojen. (Riboflavin) FAD Flavin adenin dinükleotid) transferi. Transaminaz B6 vitamini Pridoksal fosfat ……………….Amino asidlerden aldığı. (Pridoksal) -NH2 grubunu α-keto. asidlere transfer eder. Karboksilaz Biotin Biotin …………………………α-keto asidlere CO2 ‘i. bağlar(Karboksilasyon) Transformilaz Folik asid THF…………………………… -CHO, -CH2 OH, -CH3. (Tetrahidrofolat) gruplarının transferi. Transmetilaz B12 vitamini Kobamid…………………………-CH3 grubu transferi. İzomeraz koenzim.")
52
7. Enzimler hücrenin metabolik gereksinimlerine uygun şekilde aktive
veya inhibe edilerek ürün oluşum hızı kontrol edilebilir. Bu olaya enzim aktivitesinin düzenlenmesi denir. 8. Enzimler enerji türlerini biribirine dönüştürürler. Mitokondrideki küçük moleküller içinde bulunan serbest enerji, ATP enerjisi şekline dönüşür. Kasta ise ATP enerjisi, kasılma esnasında mekanik enerjiye dönüşür. 9. Enzimler tranzisyon (geçiş) durumunu stabilize ederek reaksiyonları hızlandırırlar. Moleküllerin reaksiyona girebilmeleri için enerji tüketilmektedir. Substrat ürüne dönüşürken tranzisyon (geçiş) durumundan geçer. S T* Ü
 durumunu stabilize ederek reaksiyonları. hızlandırırlar. Moleküllerin reaksiyona girebilmeleri için enerji tüketilmektedir. Substrat ürüne dönüşürken tranzisyon (geçiş) durumundan geçer. S T* Ü.")
53
Tranzisyon durumu, S ve Ü’nün serbest enerjisinden çok daha yüksek
enerjiye sahiptir. Reaksiyon tranzisyon durumundan itibaren başlar. Enerji düzeyinin tepe noktasında bulunan yüksek enerjili geçiş ürünleri daha sonra son ürüne dönüşmektedir. Enzimlerin yokluğunda, tranzisyon durumuna ulaşabilen çok az sayıda molekül, ürüne dönüşebilmektedir. Reaksiyon hızı ise, bu yüksek enerjiye sahip moleküllerin sayısı tarafından belirlenmektedir.

54
Ortama salınan serbest enerji
Ea = Aktivasyon enerjisi Ea1 = Enzimle katalizlenmemiş reaksiyonun aktivasyon enerjisi Ea2 = Enzimle katalizlenmiş reaksiyonun aktivasyon enerjisi T* Ea1 Ea Ea2 S Serbest enerji Başlangıç durumu ∆G Ortama salınan serbest enerji Ü Bitiş durumu (ürünler) Reaksiyon akış yönü
 Reaksiyon akış yönü.")
56
Bir kimyasal reaksiyon ortama enerji salıyor ise, tranzisyon
durumuna geçebilmesi için, önce aktivasyon enerjisinden enerji borç alır, sonra bu enerjiyi sarfeder. Sarfedilen enerjiden geri kalanı ise ortama serbest enerji şeklinde salınır. Ea= Tranzisyon durumu serbest enerjisi – substrat serbest enerjisi Aktivasyon enerjisi düşük olan reaksiyonların hızı yüksek olmaktadır.

58
“ Enzimle katalizlenen reaksiyonlarda, enzimler aktivasyon enerjisini
azaltarak reaksiyonları hızlandırırlar. Böylece enzim ve substratı, tranzisyon enerjisi daha düşük olan yeni bir reaksiyon yolu oluşturur.” Spontan reaksiyon Kimyasal katalizör varlığındaki reaksiyon Spesifik enzim Tranzisyon durumu Ea Başlangıç durumu ∆G kalori Bitiş durumu

59
V.ENZİMLERİN KATALİZ HIZINA ETKİ EDEN
FAKTÖRLER Kataliz Hızı, birim zamanda oluşan ürün ya da kaybolan substrat miktarıdır. Enzimle katalizlenen reaksiyonların hızını etkileyen faktörler: 1.Enzim Konsantrasyonu 2.Substrat Konsantrasyonu 3.Sıcaklık 4.pH

60
1.ENZİM KONSANTRASYONU Enzimle katalizlenen reaksiyonlarda substrat konsantrasyonu yüksek miktarlarda ise, reaksiyonun başlangıç hızı (Vİ),enzim konsantrasyonu ile doğru orantılı olarak artmaktadır. Reaksiyon hızı Vİ Enzim konsantrasyonu
,enzim konsantrasyonu ile. doğru orantılı olarak artmaktadır. Reaksiyon hızı. Vİ. Enzim konsantrasyonu.")
61
2.SUBSTRAT KONSANTRASYONU
Enzimle katalizlenen bir reaksiyonun hızı (V), ortamda enzim konsantrasyonunun sabit olması koşuluyla, substrat konsantrasyonu ile [S] birlikte hızla artar ve maksimal hız (Vmax) değerine varıncaya kadar artış devam eder. Ancak Vmax’ta substrat konsantrasyonu ne kadar artarsa artsın, kataliz hızı artmaz. Yüksek substrat konsantrasyonlarında reaksiyon hızının yavaşlaması, enzim üzerindeki substrat bağlama bölgelerinin doygunluğa ulaşmasını göstermektedir. VMAX V [S] VO
, ortamda enzim. konsantrasyonunun sabit olması koşuluyla, substrat konsantrasyonu. ile [S] birlikte hızla artar ve maksimal hız (Vmax) değerine varıncaya. kadar artış devam eder. Ancak Vmax’ta substrat konsantrasyonu ne kadar artarsa artsın, kataliz hızı. artmaz. Yüksek substrat konsantrasyonlarında reaksiyon hızının yavaşlaması, enzim üzerindeki substrat bağlama bölgelerinin doygunluğa ulaşmasını göstermektedir. VMAX. V. [S] VO.")
62
Enzimlerin çoğu Michaelis-Menten Kinetiği gösterirler.
Belli sıcaklıkta ve sabit enzim konsantrasyonunda, bu kinetiğe uyan enzimler, değişen substrat konsantrasyonu ile başlangıç hızı (VO) arasında hiperbolik bir eğri çizerler (A). Buna karşılık allosterik enzimlerde, bu eğri sigmoidal özellik taşımaktadır (B). VMAX A B VO [S]
 arasında hiperbolik bir eğri çizerler (A). Buna karşılık allosterik enzimlerde, bu eğri sigmoidal özellik. taşımaktadır (B). VMAX. A. B. VO. [S]")
63
3.SICAKLIK Kimyasal reaksiyonlarda ısının artması moleküllerin hareketini arttırarak reaksiyon hızının da artmasına yol açar. Reaksiyon hızının sıcaklık ile artışı, belli bir enerji düzeyini aşabilecek molekül sayısı ile ilgilidir. Bu durum, enzimlerle katalizlenen reaksiyonlar için de geçerlidir. “Enzimle katalizlenen bir reaksiyonda sıcaklığın yükselmesi reaksiyon hızını arttırmaktadır.” Ancak enzimler protein yapısında maddeler olduklarından, belirli bir ısı derecesinden itibaren (genelde 45ºC) enzimin denatürasyonu söz konusu olacağından, reaksiyon hızında da bir azalma meydana gelecektir.
 enzimin denatürasyonu söz. konusu olacağından, reaksiyon hızında da bir azalma meydana. gelecektir.")
64
Optimum Temperatür: Enzimin en iyi etkilediği ısı derecesidir. Bu ısı derecesi, reaksiyon hızını maksimal arttırırken, daha ilerisinde enzimin denatürasyonuna yol açar. “Enzim etkinliği düşük ısıda az, tepe noktasında en fazla, sıcaklık arttığında ise hızla düşer.” Genellikle, başlangıçta sıcaklığın her 10 ºC artışı, reaksiyon hızının iki katına çıkmasını sağlamaktadır.

65
100 reaksiyon hızı (v) denatüre olmamış protein yüzdesi 50 O O
20 30 40 50 60 sıcaklık (ºC)
")
66
Enzim aktivitesinde,optimum temperatür 2 faktörle belirlenmektedir (Kırmızı eğri).
Reaksiyon hızı, gerek spontan gerekse katalizör varlığında, sürekli olarak artar (mavi eğri). Belirli bir ısı derecesinden sonra, enzim proteini denatüre olur (siyah eğri).
. Belirli bir ısı derecesinden sonra, enzim proteini denatüre olur. (siyah eğri).")
67
Termik İnaktivasyon: Enzim çözeltisi hazırlandıktan sonra enzim aktivitesi ölçülür (Eo). Daha sonra çözelti termostata konarak her 2-3 dakikada bir ısı derecesi arttırılır ve çözeltiden bir kısım alınarak aktivite ölçülür (E). Isıtma süresi arttıkça aktivitede giderek azalma gözlenir. İnaktivasyon, zamanın logaritmik fonksiyonudur. E Eo 30ºC Log 45ºC 55ºC 60ºC zaman(dak)
. Isıtma süresi arttıkça. aktivitede giderek azalma gözlenir. İnaktivasyon, zamanın logaritmik. fonksiyonudur. E. Eo. 30ºC. Log. 45ºC. 55ºC. 60ºC. zaman(dak)")
68
4.pH Enzimatik reaksiyonlar ortamın H+ iyonu konsantrasyonundan
kolaylıkla etkilenirler. Enzim etkinliğinin en yüksek olduğu pH , optimum pH’dır. Her enzimin etki ettiği pH farklıdır. b a enzimatik aktivite pH 4 5 6 7 8 9 10

69
Alkali ortamda en iyi etkiyi gösterir.
Enzimler pH değişikliklerine bağlı olarak başlıca 2 tip aktivite eğrisi gösterirler: a) Dar sınırlar içinde etkiyi gösteren çan eğri b) Geniş sınırlar içinde etkiyi gösteren plato eğri Çoğunlukla enzimlerin optimum pH’ı 5-8 arasında değişmektedir. Maksimum aktiviteyi pH= 2’de gösterir. Nötral pH’da çalışan enzimler asidik ortamda denatüre olurlar. Optimum etki Optimum etki Enzimatik aktivite pH 2 pH 4 6 8 10 12 3 4 5 6 7 TRİPSİN Alkali ortamda en iyi etkiyi gösterir. PEPSİN
 Dar sınırlar içinde etkiyi gösteren çan eğri. b) Geniş sınırlar içinde etkiyi gösteren plato eğri. Çoğunlukla enzimlerin optimum pH’ı 5-8 arasında değişmektedir. Maksimum aktiviteyi. pH= 2’de gösterir. Nötral. pH’da çalışan enzimler. asidik ortamda denatüre. olurlar. Optimum etki. Optimum. etki. Enzimatik aktivite. pH. 2. pH TRİPSİN. Alkali ortamda en iyi etkiyi gösterir. PEPSİN.")
70
pH değişikliklerinin enzimatik aktivite üzerindeki etkileri aşağıdaki
faktörler tarafından belirlenmektedir. 1. Ortamın çok yüksek ve çok düşük pH düzeyleri, enzimin denatürasyonuna yol açarak, yapısında geri dönüşümsüz değişiklikler yapar. 2. Apoenzim ile koenzim arasındaki bağlar etkilenir. 3. Bir protein olan enzimin yapısındaki amino ve karboksil gruplarının iyonizasyon durumu, ortamın pH değerine bağlı olarak değişir. Aktif bölgedeki iyonizasyon değişikleri, enzim-substrat reaksiyonunu ve buna bağlı olarak katalizi bozar.

71
Örnek: Katalitik aktivite için ortamdan H+ iyonu kazanılması gerekiyor ise, moleküldeki amino gruplarının protonlaşması gibi: - NH NH3+ Alkali pH ‘da proton kaybı olacağından, verilen örnekte enzimatik aktivite hızı düşer. 4. Büyük çoğunlukla pH değişikliklerinden substratın da iyonizasyon durumu etkilenir. H+

72
VI.ENZİM KİNETİĞİ Reaksiyon Hızı (v):
Enzim etkisiyle birim zamanda kaybolan substrat miktarı veya oluşan ürün miktarı ile ölçülür. Enzim Moleküler Aktivitesi: Optimum reaksiyon şartlarında, 1 molekül enzim tarafından 1 dakikada ürüne dönüşen substrat miktarıdır. Spesifik Enzim Aktivitesi: mg protein başına düşen enzim ünite sayısıdır. Enzim Ünitesi: Optimal şartlarda, 1 dakikada 1 mikromol (μmol) substratı ürüne dönüştüren enzim miktarıdır.
 substratı ürüne. dönüştüren enzim miktarıdır.")
73
Enzimlerin katalizledikleri reaksiyonlarda genel kimyasal reaksiyon
kinetikleri geçerlidir. Michaelis ve Menten isimli araştırmacılar, enzimlerle gerçekleşen reaksiyonlar için basit bir tanımlama yapmışlardır. Enzim kinetiklerinin kantitatif analizleri için geliştirilen bu model, tek substratlı reaksiyonlar için geçerlidir. E + S ES E + Ü k1, k2 ve k3 : reaksiyonların hız sabitleri k1 k3 k2

74
Tersinir olarak sabit bir hızla (k1) substratla [S] birleşen enzim [E],
önce enzim-substrat [ES] kompleksini oluşturur. [ES] kompleksi daha sonra başlıca 2 akıbete uğrayabilir: 1. Sabit bir hızla (k2) yeniden E ve S’a dönüşür. 2. Yahut k3 sabit hızıyla ürün [Ü] oluşurken enzim de serbestleşerek ilk yapısını kazanır. ES oluşum hızı = k1[E] [S] ES yıkılım hızı = (k2 +k3) [ES]
![Tersinir olarak sabit bir hızla (k1) substratla [S] birleşen enzim [E],](http://slideplayer.biz.tr/slide/9229731/27/images/74/Tersinir+olarak+sabit+bir+h%C4%B1zla+%28k1%29+substratla+%5BS%5D+birle%C5%9Fen+enzim+%5BE%5D%2C.jpg "önce enzim-substrat [ES] kompleksini oluşturur. [ES] kompleksi daha sonra başlıca 2 akıbete uğrayabilir: 1. Sabit bir hızla (k2) yeniden E ve S’a dönüşür. 2. Yahut k3 sabit hızıyla ürün [Ü] oluşurken enzim de serbestleşerek ilk. yapısını kazanır. ES oluşum hızı = k1[E] [S] ES yıkılım hızı = (k2 +k3) [ES]")
75
Reaksiyon hızı ile substrat konsantrasyonu arasındaki ilişkiyi tanımlayan
Michaelis-Menten denklemi kurulurken aşağıdaki varsayımlar gözönüne alınmıştır: 1. Substrat konsantrasyonu [S], enzim konsantrasyonun [E]’dan çok daha fazladır. Böylece belirli bir zamanda enzime bağlı olan substrat miktarı ihmal edilebilir. 2. Reaksiyonun denge durumunda ES kompleksinin oluşum ve yıkılım hızları biribirine eşittir.

76
Reaksiyonun denge durumunda : k1 [E] [S] = [k2 + k3] [ES] (1)
Km k1
![Reaksiyonun denge durumunda : k1 [E] [S] = [k2 + k3] [ES] (1)](http://slideplayer.biz.tr/slide/9229731/27/images/76/Reaksiyonun+denge+durumunda+%3A+k1+%5BE%5D+%5BS%5D+%3D+%5Bk2+%2B+k3%5D+%5BES%5D+%281%29.jpg "Km. k1.")
77
Michaelis-Menten denklemi hiperbolik bir eğrinin denklemidir.
V VMAX Reaksiyon hızı VMAX 2 [S] (mol/L) Km
 Km.")
78
VMAX : Katalizin ulaşabileceği en yüksek hız değeridir.
Enzim bölgeleri substrat ile tam doygunluğa geçince VMAX’a ulaşılır.

79
Km : (Michaelis-Menten Sabiti)
En yüksek hız (VMAX) değerinin yarısına ulaşmak için gerekli substrat miktarıdır. Ortamda bulunan tüm enzim moleküllerinin aktif bölgelerinin yarısını dolduran substrat miktarıdır. Km değerini tam olarak bulabilmek için farklı konsantrasyonlarda substrat kullanılmalıdır.
 değerinin yarısına ulaşmak için gerekli substrat. miktarıdır. Ortamda bulunan tüm enzim moleküllerinin aktif bölgelerinin yarısını. dolduran substrat miktarıdır. Km değerini tam olarak bulabilmek için farklı konsantrasyonlarda. substrat kullanılmalıdır.")
80
Km , bir enzime ve substratına özgüldür.
Enzimin substratına ilgisini (affinitesi) ni yansıtır. Km , enzim-substrat ilişkisinde bir ölçüdür. Km’i düşük olan bir enzim, substratına yüksek ilgi (affinite) gösterir. Enzim, aşağı substrat konsantrasyonunda doyar. Tersine büyük KM , enzimin substratına düşük ilgisini tanımlamaktadır. Km = mol/L olarak ifade edilir. Bir çok enzim için bu değer 10-3 – mol/L arasında değişir. Km = k2 + k3 k1 (mol/L)
 ni yansıtır. Km , enzim-substrat ilişkisinde bir ölçüdür. Km’i düşük olan bir enzim, substratına yüksek ilgi (affinite) gösterir. Enzim, aşağı substrat konsantrasyonunda doyar. Tersine büyük KM , enzimin substratına düşük ilgisini tanımlamaktadır. Km = mol/L olarak ifade edilir. Bir çok enzim için bu değer 10-3 – 10-5 mol/L arasında değişir. Km = k2 + k3. k1. (mol/L)")
81
Enzim a’nın küçük Km ‘i, enzimin substrata ilgisinin yüksek olduğunu yansıtır.
Enzim b’nin büyük Km’i, enzimin substrata olan ilgisinin az olduğunu yansıtır. V a VMAX b Reaksiyon hızı VMAX 2 [S] Kma Kmb

82
[S]= Km : Substrat Kmdeğerine eşitse, başlangıç hızı,
Yüksek substrat konsantrasyonlarında [S] > Km reaksiyon hızı sıfırıncı basamaktır-yani, substrat konsantrasyonundan bağımsız ve sabittir. vmax Vmax 2 Düşük substrat konsantrasyonunda [S] < Km, , reaksiyon hızı birinci basmaktır-yani, substrat konsantrasyonuyla orantılıdır. o [S] Km [S]= Km : Substrat Kmdeğerine eşitse, başlangıç hızı, maksimal hızın yarısına eşittir. Vmax 2 Vi =
![[S]= Km : Substrat Kmdeğerine eşitse, başlangıç hızı,](http://slideplayer.biz.tr/slide/9229731/27/images/82/%5BS%5D%3D+Km+%3A+Substrat+Kmde%C4%9Ferine+e%C5%9Fitse%2C+ba%C5%9Flang%C4%B1%C3%A7+h%C4%B1z%C4%B1%2C.jpg "Yüksek substrat konsantrasyonlarında. [S] > Km reaksiyon hızı sıfırıncı basamaktır-yani, substrat konsantrasyonundan bağımsız ve sabittir. vmax. Vmax. 2. Düşük substrat konsantrasyonunda [S] < Km, , reaksiyon hızı birinci basmaktır-yani, substrat konsantrasyonuyla. orantılıdır. o. [S] Km. [S]= Km : Substrat Kmdeğerine eşitse, başlangıç hızı, maksimal hızın yarısına eşittir. Vmax. 2. Vi =")
83
Belirli bir andaki kataliz hızı: Vo = Michaelis-Menten Eşitliği
Michaelis-Menten denklemi hiperbolik bir eğrinin denklemidir. Bu denklem tersine çevrildiğinde düz eğri elde edilir. Düz eğrinin (doğru) çizilmesi ile Km ve VMAX değerlerinin duyarlı bir biçimde belirlenmesi kolaylaşmaktadır. Bu doğru ayrıca enzim inhibitörlerinin etki mekanizmalarının saptanmasında da kullanılır. VMAX [S] [S] + Km
 çizilmesi ile Km ve VMAX değerlerinin duyarlı. bir biçimde belirlenmesi kolaylaşmaktadır. Bu doğru ayrıca enzim inhibitörlerinin etki mekanizmalarının saptanmasında da kullanılır. VMAX [S] [S] + Km.")
84
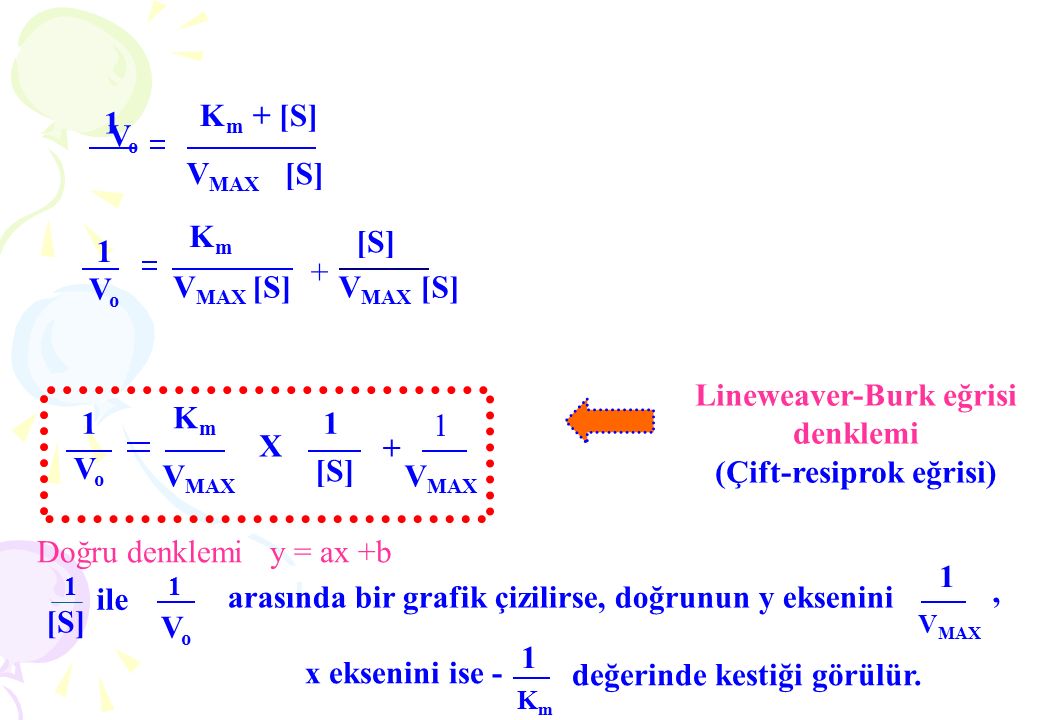
Lineweaver-Burk eğrisi (Çift-resiprok eğrisi)
1 Km + [S] 1 Vo VMAX [S] Km [S] 1 + Vo VMAX [S] VMAX [S] Lineweaver-Burk eğrisi denklemi (Çift-resiprok eğrisi) 1 Km 1 1 X + Vo VMAX [S] VMAX Doğru denklemi y = ax +b 1 1 arasında bir grafik çizilirse, doğrunun y eksenini , ile [S] Vo VMAX 1 x eksenini ise - değerinde kestiği görülür. Km
 1. Km X. + Vo. VMAX. [S] VMAX. Doğru denklemi y = ax +b arasında bir grafik çizilirse, doğrunun y eksenini. , ile. [S] Vo. VMAX. 1. x eksenini ise - değerinde kestiği görülür. Km.")
85
. . Lineweaver-Burk çift-resiprok Eğrisi 1 V0 Km Eğim VMAX 1 VMAX 1 1

86
Km Değerinin Bilinmesinin Önemi
Enzimlerin Saflaştırılması Dokularda enzim aktivitesinin saptanması İlaç imalatında Enzim inhibitörlerinin belirlenmesi

87
VII.ENZİM AKTİVİTESİNİN İNHİBİSYONU
Enzimle katalizlenen bir reaksiyonun hızını azaltan ya da engelleyen maddeye inhibitör adı verilir. İnhibitör, bir molekül yada iyon olabilir. Enzim katalizinin engellenmesi olayına ise inhibisyon denir. Enzim Katalizinin İnhibisyonu (engellenmesi) 1. Enzimlerin yapısal özelliklerinin ve katalitik aktivitelerinin incelenmesinde önemli rol oynar. 2. Hücre içindeki metabolik yolların belirlenmesinde yol gösterir. 3. Biyolojik sistemlerin temel kontrol mekanizmasını oluşturur. 4. Pek çok ilaç ve zehirli bileşik tarafından ortaya çıkabilir.
 1. Enzimlerin yapısal özelliklerinin ve katalitik aktivitelerinin. incelenmesinde önemli rol oynar. 2. Hücre içindeki metabolik yolların belirlenmesinde yol gösterir. 3. Biyolojik sistemlerin temel kontrol mekanizmasını oluşturur. 4. Pek çok ilaç ve zehirli bileşik tarafından ortaya çıkabilir.")
88
Enzimlerin inhibisyonu;
reversibl (tersinir, geriye dönüşümlü) veya irreversibl (tersinmez, geriye dönüşümsüz) olarak iki grupta incelenir. 1-Geriye dönüşümlü (reversibl) inhibisyonlar: Bu tip inhibisyonlarda substrat konsantrasyonunun ya da inhibitöre oranla enzim konsantrasyonunun arttırılması ile enzim inhibitör ilişkisi tersine çevrilebilmektedir. a)Yarışmalı (kompetitif) inhibisyon: b)Yarışmasız (nonkompetitif) İnhibisyon: c)Yarışmasız (unkompetitif) inhibisyon:
 veya. irreversibl (tersinmez, geriye dönüşümsüz) olarak iki grupta incelenir. 1-Geriye dönüşümlü (reversibl) inhibisyonlar: Bu tip inhibisyonlarda substrat konsantrasyonunun ya da inhibitöre oranla enzim konsantrasyonunun arttırılması ile enzim inhibitör ilişkisi tersine çevrilebilmektedir. a)Yarışmalı (kompetitif) inhibisyon: b)Yarışmasız (nonkompetitif) İnhibisyon: c)Yarışmasız (unkompetitif) inhibisyon:")
89
a)Yarışmalı (kompetitif) inhibisyon:
İnhibitör (İ) enzimin aktif bölgesine substratla (S) yarışarak bağlanmaktadır. İnhibitör molekülünün yapısı substratınkine benzediğinden dolayı enzim ile inhibitör kolaylıkla bağlanırlar, ancak ürün oluşamamaktadır. Bu tip inhibitörler, enzime asıl substratın bağlanmasını engelleyen maddelerdir. Bu tür inhibisyonda Vmax’a ulaşmak için daha fazla substrat gereklidir. Ortamda substrat moleküllerinin arttırılmasıyla enzimin inhibitöre olan ilgisi azalmakta ve inhibisyon ortadan kalkmaktadır.
 enzimin aktif bölgesine substratla (S) yarışarak. bağlanmaktadır. İnhibitör molekülünün yapısı substratınkine benzediğinden dolayı enzim ile inhibitör kolaylıkla bağlanırlar, ancak ürün oluşamamaktadır. Bu tip inhibitörler, enzime asıl substratın bağlanmasını engelleyen maddelerdir. Bu tür inhibisyonda Vmax’a ulaşmak için daha fazla substrat gereklidir. Ortamda substrat moleküllerinin arttırılmasıyla enzimin inhibitöre olan ilgisi azalmakta ve inhibisyon ortadan kalkmaktadır.")
90
Kompetitif İnhibisyon (Vmax değişmez, Km artar).
.")
91
VMAX VMAX 2 [S] Km Km 1 Vo 1 VMAX 1 Km 1 Km 1 [S] İnhibitörsüz
Yarışmalı İnhibitör ile VMAX 2 Reaksiyon hızı (Vo) [S] Km Km 1 Vo Yarışmalı inhibitör ile 1 VMAX 1 Km inhibitörsüz 1 Km 1 [S]
![VMAX VMAX 2 [S] Km Km 1 Vo 1 VMAX 1 Km 1 Km 1 [S] İnhibitörsüz](http://slideplayer.biz.tr/slide/9229731/27/images/91/VMAX+VMAX+2+%5BS%5D+Km+Km+1+Vo+1+VMAX+1+Km+1+Km+1+%5BS%5D+%C4%B0nhibit%C3%B6rs%C3%BCz.jpg "Yarışmalı. İnhibitör ile. VMAX. 2. Reaksiyon. hızı (Vo) [S] Km. Km. 1. Vo. Yarışmalı. inhibitör ile. 1. VMAX. 1. Km. inhibitörsüz. 1. Km. 1. [S]")
92
İnhibitör varlığında VMAX değişmez, buna karşılık Km
değişir. İnhibitör varlığında Km değeri büyür, enzimin substrata olan ilgisi azalmıştır. Km büyümüştür. Çünkü küçülmüştür ve Km (eğrinin VMAX eğimi) büyümüştür. Km
 büyümüştür. Km.")
93
Bir molekülün, belirli bir enzimin inhibitörü olup olmadığı,yahut
inhibisyonun tipi, kinetik analizlerle ortaya konulur! Metanol zehirlenmesinin tedavisinde yarışmalı inhibisyondan faydalanılır. Metanol, alkol dehidrogenaz etkisiyle FORMALDEHİD’e dönüşür. Formaldehid körlüğe ve müdahale edilmez ise ölüme yol açar. Metanol zehirlenmesinde damar-içi etanol uygulanır. Etanol, alkol dehidrogenaza bağlanmada metanol ile yarışır.Enzim Etanolü daha çok toksik olmayan asetaldehide dönüştürür. Böylece toksik formaldehid oluşumu yarışmalı olarak engellenir.

94
E +S ES E +Ü VMAX Km DEĞİŞMEZ Eİ + S EİS Ü
b)Yarışmasız (nonkompetitif) İnhibisyon: Substratla yapısal benzerliği olmayan inhibitör, substratla aynı bölgeye bağlanmaz. Yarışmasız inhibitör, ya serbest enzime ya da ES kompleksine bağlanarak reaksiyonu yavaşlatır. Substrat konsantrasyonu arttırılarak inhibisyon ortadan kaldırılamaz. E +S ES E +Ü VMAX Km DEĞİŞMEZ Eİ + S EİS Ü +İ +İ
Yarışmasız (nonkompetitif) İnhibisyon: Substratla yapısal benzerliği olmayan inhibitör, substratla aynı bölgeye bağlanmaz. Yarışmasız inhibitör, ya serbest enzime ya da ES kompleksine bağlanarak reaksiyonu yavaşlatır. Substrat konsantrasyonu arttırılarak inhibisyon ortadan kaldırılamaz. E +S ES E +Ü VMAX. Km DEĞİŞMEZ. Eİ + S EİS Ü. +İ. +İ.")
95
Non kompetitif İnhibisyon (Vmax azalır,Km değişmez)
")
96
1 VMAX VMAX VMAX 2 VMAX 2 [S] Km 1 Vo 1 VMAX 1 1 Km [S] İnhibitörsüz
Yarışmasız İnhibitör VMAX 2 [S] Km 1 Vo Yarışmasız inhibitör 1 VMAX 1 VMAX inhibitörsüz 1 Km 1 [S]
![1 VMAX VMAX VMAX 2 VMAX 2 [S] Km 1 Vo 1 VMAX 1 1 Km [S] İnhibitörsüz](http://slideplayer.biz.tr/slide/9229731/27/images/96/1+VMAX+VMAX+VMAX+2+VMAX+2+%5BS%5D+Km+1+Vo+1+VMAX+1+1+Km+%5BS%5D+%C4%B0nhibit%C3%B6rs%C3%BCz.jpg "Yarışmasız. İnhibitör. VMAX. 2. [S] Km. 1. Vo. Yarışmasız. inhibitör. 1. VMAX. 1. VMAX. inhibitörsüz. 1. Km. 1. [S]")
97
c)Yarışmasız (unkompetitif) inhibisyon:
İnhibitör, ES kompleksine bağlanır. İnhibitör bağlandığında ürün oluşumu sonlanır. E +S ES E + Ü + İ EİS Ü VMAX Km

98
Unkompetitif İnhibisyon (Vmax ve Km Azalır)
")
99
İ VMAX VMAX 2 [S] Km Km 1 Vo 1 VMAX 1 1 VMAX Km 1 1 [S] Km
Unkompetitif inhibitör VMAX 2 VMAX [S] Km Km 1 Vo İ 1 VMAX 1 VMAX 1 Km 1 Km 1 [S]
![İ VMAX VMAX 2 [S] Km Km 1 Vo 1 VMAX 1 1 VMAX Km 1 1 [S] Km](http://slideplayer.biz.tr/slide/9229731/27/images/99/%C4%B0+VMAX+VMAX+2+%5BS%5D+Km+Km+1+Vo+1+VMAX+1+1+VMAX+Km+1+1+%5BS%5D+Km.jpg "Unkompetitif. inhibitör. VMAX. 2. VMAX. [S] Km. Km. 1. Vo. İ. 1. VMAX. 1. VMAX. 1. Km. 1. Km. 1. [S]")
100
2- Geriye dönüşümsüz (tersinmez, irreversibl inhibisyon):
Bu tip inhibisyonlarda aktif bölgeye kovalent olarak bağlanan inhibitör, enzim yapısını bozduğu için geriye dönüş olmamaktadır. Sinir gazlarından DIPF (diizopropil fosfofluoridat) asetilkolin isimli nörotransmittörü parçalayan asetilkolin esteraz’ ın tersinmez inhibitörüdür. Asetilkolin bir sinir hücresindeki uyarıyı diğer bir hücreye aktaran maddedir. Bu etkisi sona erdikten sonra, asetilkolin esteraz tarafından hidrolize uğrar. Asetilkolin esteraz’ın aktif bölgesinin DIPF tarafından tersinmez inhibe edilmesi hidroliz olayını engeller ve bunun sonucunda felç meydana gelir.
 asetilkolin isimli. nörotransmittörü parçalayan asetilkolin esteraz’ ın tersinmez. inhibitörüdür. Asetilkolin bir sinir hücresindeki uyarıyı diğer bir hücreye aktaran maddedir. Bu etkisi sona erdikten sonra, asetilkolin esteraz tarafından hidrolize uğrar. Asetilkolin esteraz’ın aktif bölgesinin DIPF tarafından tersinmez inhibe. edilmesi hidroliz olayını engeller ve bunun sonucunda felç meydana gelir.")
101
Tarımda kullanılan insektisidler (böcek öldürücüler) de asetilkolin
esterazın tersinmez engelleyicisidir. Birçok insektisidin bileşiminde bulunan fosforlu organik yapı aktif bölgesinde serin amino asidini içeren enzimlerle geriye dönüşümsüz inhibisyon yapar. Kurşun zehirlenmesi Kurşun, proteinlerin yapısında yer alan sistein isimli aminoasidin sülfhidril grubuyla (-SH) kovalent bağlar oluşurur. Hemoglobin sentezinde, protoporfirin isimli maddeye Fe+2 girişini katalizleyen ferrokelataz ve yine bu sentezde yer alan δ-aminolevülinat dehidraz isimli enzimler kurşunun geri dönüşümsüz inhibisyonuna duyarlı enzimlerdir.
 kovalent bağlar oluşurur. Hemoglobin sentezinde, protoporfirin isimli maddeye Fe+2 girişini. katalizleyen ferrokelataz ve yine bu sentezde yer alan δ-aminolevülinat. dehidraz isimli enzimler kurşunun geri dönüşümsüz inhibisyonuna duyarlı enzimlerdir.")
102
3-Bazı ilaçlar da enzim inhibitörü olarak etki ederler.
Penisilin ve amoxicillin gibi yaygın olarak kullanılan antibiotikler,bakteri duvarı sentezine ait enzimleri inhibe ederek etki gösterirler. ACE (angiotensin-konverting enzim) inhibitörleri (captopril, enapril gibi) kan basıncını düşüren ilaçlardır. Bu etkilerini, angiotensin I’i vazokonstriktör (damar büzücü) etkili angiotensin II’ye çeviren ACE’yi bloke ederek gösterirler. Angiotensin I Angiotensin II (inhibitör etki gösteren ilaçlar) captopril, enapril ACE (Damar büzücü etkili)
 inhibitörleri (captopril, enapril gibi) kan basıncını düşüren ilaçlardır. Bu etkilerini, angiotensin I’i vazokonstriktör (damar büzücü) etkili. angiotensin II’ye çeviren ACE’yi bloke ederek gösterirler. Angiotensin I Angiotensin II. (inhibitör etki. gösteren ilaçlar) captopril, enapril. ACE. (Damar büzücü etkili)")
103
VIII. ENZİM AKTİVİTESİNİ DÜZENLENMESİ
Bir metabolik yolun belli bir hızda ve yönde ilerleyebilmesi için kontrol altında tutulması gerekir, bu da enzimlerin sayesinde olur. Enzim aktivitesi 2 şekilde kontrol edilebilir: 1-Enzimin mutlak (absolü) miktarının değişimi 2-Enzimin katalitik etkinliğinin değişimi 1-ENZİMİN MUTLAK MİKTARININ DEĞİŞİMİ Organizma varolan enzim miktarını, enzim sentez hızlarını değiştirerek düzenleyebilir. Enzim proteinin sentezi ihtiyaca göre artabilir (indüklenme) ya da azalabilir (represyon, baskılanma). İndüksiyon ile enzim sentezi artar, represyon ile enzim sentezi azalır. Derepresyon ile enzim sentezi baskıdan kurtulup tekrardan artar.
 miktarının değişimi. 2-Enzimin katalitik etkinliğinin değişimi. 1-ENZİMİN MUTLAK MİKTARININ DEĞİŞİMİ. Organizma varolan enzim miktarını, enzim sentez hızlarını değiştirerek düzenleyebilir. Enzim proteinin sentezi ihtiyaca göre artabilir (indüklenme) ya da azalabilir (represyon, baskılanma). İndüksiyon ile enzim sentezi artar, represyon ile enzim sentezi azalır. Derepresyon ile enzim sentezi baskıdan kurtulup tekrardan artar.")
104
Enzim sentezi indükleyici bir maddeye cevap olarak tetiklenebilir.
E. coli isimli mikroorganizma glukozun bulunduğu ortamda glukozu kullanır. Ancak bir disakkarid olan laktozu, β-galaktozidaz isimli enzimi bulunmadığından dolayı galaktoz ve glukoza hidrolizleyemez. Ortamdan glukoz çıkarılıp yerine laktoz ilave edildiğinde, mikroorganizma laktozu kullanır hale gelir. Laktoz (indüktör) protein yapısında olan β-galaktozidazın sentezlenmesini indüklemiştir. İndükleyici maddelerin çoğu, enzimlerin substratlarıdır. Ancak substrata yapısal bakımdan benzeyen bileşikler de indükleyici madde olabilirler fakat substrat olamazlar!
 protein yapısında olan β-galaktozidazın. sentezlenmesini indüklemiştir. İndükleyici maddelerin çoğu, enzimlerin substratlarıdır. Ancak substrata yapısal bakımdan benzeyen bileşikler de indükleyici. madde olabilirler fakat substrat olamazlar!")
105
Salmonella isimli mikroorganizma histidin bulunan ortamda, kendisi
histidin sentezleyemez ( represyon, baskı altına alma). Histidin korepresör’dür, yani kendi sentezi ile ilgili enzimlerin sentezini engellemektedir. Histidin represyon etkisini bir proteine bağlanarak yapar. Bu proteine aporepresör denir. Aporepresör-korepresör, enzim sentezini kontrol altına alır. Ortamdan histidin çıkarılması yahut bu maddenin tükenmesi, enzim biosentezinin tekrardan başlamasını sağlar (derepresyon, baskının ortadan kaldırılışı).
. Histidin korepresör’dür, yani kendi sentezi ile ilgili enzimlerin sentezini engellemektedir. Histidin represyon etkisini bir proteine bağlanarak yapar. Bu proteine aporepresör denir. Aporepresör-korepresör, enzim sentezini kontrol altına alır. Ortamdan histidin çıkarılması yahut bu maddenin tükenmesi, enzim biosentezinin tekrardan başlamasını sağlar (derepresyon, baskının ortadan kaldırılışı).")
106
2-ENZİMİN KATALİTİK ETKİNLİĞİNİN DEĞİŞİMİ
1. Enzim molekülünün aktivasyon-inhibisyonu (Allosterik enzimler) 2. Kovalent Modifikasyon 3. Zimojen Aktivasyonu 1.ALLOSTERİK ENZİMLER Allosterik “başka yere ait” anlamına gelir. Aktif bölgeleri dışında bir yere nonkovalent olarak bağlanan efektör (modülatör) isimli moleküller tarafından düzenlenirler. Allosterik efektörün enzime bağlanması sonucunda, enzimin substratına olan ilgisi değişir. Efektör, enzim aktivitesini inhibe ettiğinde negatif efektör, aktiviteyi arttırdığında pozitif efektör denir.
 2. Kovalent Modifikasyon. 3. Zimojen Aktivasyonu. 1.ALLOSTERİK ENZİMLER. Allosterik başka yere ait anlamına gelir. Aktif bölgeleri dışında bir yere nonkovalent olarak bağlanan. efektör (modülatör) isimli moleküller tarafından düzenlenirler. Allosterik efektörün enzime bağlanması sonucunda, enzimin. substratına olan ilgisi değişir. Efektör, enzim aktivitesini inhibe ettiğinde negatif efektör, aktiviteyi arttırdığında pozitif efektör denir.")
107
Birden fazla alt üniteden meydana gelmişlerdir (oligomerik enzimler).
HIZ DÜZENLEYİCİ ENZİMLERİN ÇOĞU ALLOSTERİK ENZİMLERDİR. Birden fazla alt üniteden meydana gelmişlerdir (oligomerik enzimler). Çoğunlukla metabolik yolun ilk basamağına etki ederler. Enzim moleküllerinin üzerinde bir katalitik, bir de düzenleyici bölge bulunur. Düzenleyici bölgeye efektör, nonkovalent olarak bağlanır.
. Çoğunlukla metabolik yolun ilk basamağına etki ederler. Enzim moleküllerinin üzerinde bir katalitik, bir de düzenleyici. bölge bulunur. Düzenleyici bölgeye efektör, nonkovalent olarak bağlanır.")
108
II. İNHİBİTÖR I. SUBSTRAT (negatif efektör) (pozitif efektör)
KATALİTİK BÖLGE II. İNHİBİTÖR AKTİVATÖR I. (negatif efektör) (pozitif efektör) DÜZENLEYİCİ BÖLGE
 (pozitif efektör) DÜZENLEYİCİ BÖLGE.")
109
İki alt birimli bir enzimin 1
İki alt birimli bir enzimin 1. alt ünitesine efektörün bağlanması sonucunda, 2. alt birimin substrat bağlanması etkilenmektedir. Efektör, 2. alt birime substrat bağlanmasını hızlandırabilir ya da yavaşlatabilir. Bu olaya kooperativite adı verilir. Ortamda bulunan allosterik aktivatör enzimin etkinliğini arttırır, allosterik inhibitör ise enzimin etkinliğini azaltır. Allosterik enzimlerde, substrat ile hız arasındaki hiperbolik eğri, sigmoidal karakter kazanır. Bu grafik lineer olarak çizildiğinde ise kırık şekil alır. Böyle bir grafik enzimin 2 veya daha fazla alt üniteden meydana geldiğini gösterir.

110
V Sigmoidal eğri S 1/V kırık eğri 1/S

111
Substratın kendisi aktivatör ya da inhibitör olarak davrandığında
homotropik etki denir. Bu olaya 4 altüniteden meydana gelmiş enzimlerde rastlanır. Bir allosterik enzim kendi substratından başka bir efektör tarafından aktive ya da inhibe edilmekte ise heterotropik etkiden bahsedilir.

112
Bir ara ürün veya son ürün de enzim etkinliğinin düzenlenmesinde
görev yapabilir. Bu ürün kullanılmıyor birikiyor ise, bu yolda görevli 1. veya 2. enzimi “negatif feed back” başa tepki şeklinde inhibisyona uğratır. A B C D D’nin konsantrasyonu, sentezlendiği kadar tüketilmediğinden dolayı artacak olursa, metabolik yoldaki ilk enzim inhibe olur. Düzenleyici enzimler sayesinde son ürünün birikimi engellenmiş olur! E1 E2 E3 Hız düzenleyici enzim Feed back İnhibisyon

113
2. KOVALENT MODİFİKASYONA UĞRAYAN SİSTEMLER
Bazı enzimlerin katalitik etkileri kovalent modifikasyonlarla değişebilir. Aktivitelerinde kovalent modifikasyona uğrayan enzimler biribirine dönüşebilen enzimlerdir. Bu enzimler iki aktivite halinde bulunurlar: Yüksek ve düşük aktivite En sık rastlanan modifikasyon şekli, enzim molekülünün yapısındaki belirli serin, treonin veya tirozin isimli aminoasidlere bir fosfat grubunun eklenmesi veya bir fosfat grubunun çıkarılmasıdır.

114
O OH Fosforilasyon Enzim Enzim O-P-O- O- Defosforilasyon
Bazı enzimlerde fosfoenzim, bazılarında ise defosfo enzim şekil daha aktif olabilmektedir. Metabolik yolun gereksinimine uygun olarak, fosfor grubunun enzime ilavesi ya da enzimden ayrılması sonucunda, enzim iki faklı şekilde çalışmaktadır. Organizmanın glukoza gereksinimi olduğu esnada, glikojen sentataz fosforillenerek aktivitesini kaybeder. Aynı esnada glikojen fosforilaz bir fosfat grubu bağlayarak aktif şekle dönüşür. Bu sayede depo maddesi glikojenden glukoz sağlanmış olur. Fosforilasyon ve defosforilasyon sırası ile protein kinaz ve protein fosfataz ismi verilen enzimler tarafından gerçekleştirilmektedir. Buenzimler ise hormonal ve sinirsel kontrol altında tutulmaktadır. O OH Fosforilasyon Enzim Enzim O-P-O- serin serin O- Defosforilasyon

115
Kinaz ve Fosfataz isimli enzimler kovalent modifikasyonun reversibl
(geri dönüşebilir) oluşunu sağlayan enzimlerdir. ATP ADP Mg+2 Kinaz Enzim-Serin Enzim-Serin O-PO3-2 Fosfataz Mg+2 Pi H2O ENZİMİN KOVALENT MODİFİKASYONU
 oluşunu sağlayan enzimlerdir. ATP. ADP. Mg+2. Kinaz. Enzim-Serin. Enzim-Serin. O-PO3-2. Fosfataz. Mg+2. Pi. H2O. ENZİMİN KOVALENT MODİFİKASYONU.")
116
3.ZİMOJEN AKTİVASYONU Hücre dışında görev yapan bazı enzimler, bulundukları yere zarar vermemeleri için aktif olamayan öncül moleküller şeklinde sentez edilirler. Bu moleküllere proenzim veya zimojen adı verilmektedir. Zimojen aktivasyonu bir veya birkaç peptid bağının koparılması (yarılması, kırılması) ile olur. Pankreasta sentezlenen tripsinojen (inaktif) etki gösterdiği barsaklara salgılandığında peptid bağları kırılır ve aktif şekle dönüşür.
 ile olur. Pankreasta sentezlenen tripsinojen (inaktif) etki gösterdiği. barsaklara salgılandığında peptid bağları kırılır ve aktif şekle. dönüşür.")
117
Kimotripsinojen Kimotripsin
Tripsinojen Tripsin (İnaktif) (Aktif) Kimotripsinojen Kimotripsin Kan plazmasında: Fibrinojen Fibrin Enterokinaz Tripsin Proteinlerdeki aromatik aminoasidleri ayırır. (Tirozin,fenilalanin, triptofan) Trombin
 (Aktif) Kimotripsinojen Kimotripsin. Kan plazmasında: Fibrinojen Fibrin. Enterokinaz. Tripsin. Proteinlerdeki aromatik. aminoasidleri ayırır. (Tirozin,fenilalanin, triptofan) Trombin.")