Sunuyu indir
1
Protein Sentezi (Translasyon)
Dr. Hatice Mergen 1 1 1 1

2
Protein Metabolizması
Proteinler birçok informasyon yolunun son ürünüdür. Tipik bir hücrede binlerce farklı protein vardır. Bu proteinler hücrenin ihtiyaçlarına göre sentezlenir ve uygun hücresel hedeflere yönlendirilirler. Protein biyosentezi en kompleks biyosentetik işlemdir. Ökaryotik protein sentezine 70’in üzerinde ribozomal protein, 20 veya daha fazla aktive olmuş amino asit öncülü, bir düzineden daha fazla yardımcı enzim ve faktör başlama, uzama ve polipeptid sonlanması için gereklidir.

3
Protein Metabolizması
Ayrıca, farklı proteinlerin final işlenmesi için ek olarak 100 kadar enzim gereklidir. Sonuç olarak, 300’den fazla sayıda farklı makromolekül protein sentezinde görev alır. Protein biyosentezi hücredeki tüm biyosentetik reaksiyonlarda kullanılan enerjinin % 90’ını kullanır. Tipik bir bakteri hücresinde ribozom, ilişkili protein faktörü ve enzim, tRNA hücrenin kuru ağırlığının % 35’inden fazlasını oluşturur. Olayın karmaşıklığına rağmen proteinler oldukça hızlı sentez edilirler. Bir E. coli hücresinde 100 amino asitlik bir polipeptit yaklaşık 5 saniyede sentez edilir.

4
Ribozomlar ve Endoplazmik Retikulum

5
Genetik Kod Amino asidler, amino açil-tRNA sentetaz enzimi ile aminoaçil-tRNA’ları oluştururlar. Crick’in adaptör molekülü hipotezinde adaptör molekül bir uçta amino asidleri bağlarken diğer uçta amino asit ile ilişkili mRNA dizisi ile bağlantı kurar. Bilgi böylece 4 bazlı nükleik asit dizilerinden 20 amino asitli proteinlere çevrilir (Translasyon).
.")
6
Crick’in adaptör hipotezi
Amino asidler kovalent olarak tRNA’nın 3′ ucuna bağlanır. tRNA’daki üçlü kodon, mRNA’daki bazlar ile komplementer olarak hidrojen bağı ile bağlanır.

7
Genetik kod sentetik mRNA’lar kullanılarak çözülmüştür
Dört nükleotidin ikili olarak değişik şekillerde bir araya gelme olasılığı 42=16 Dört nükleotidin üçlü olarak değişik şekillerde bir araya gelme olasılığı 43=64 Kodon: Spesifik bir amino asidi kodlayan üç nükleotid’ten oluşan dize Başlama (initiasyon) kodonu (AUG), tüm hücrelerde bir polipeptidi başlatan sinyal kodonu (Bir polipeptidin içinde sinyal ayrıca Met’i kodlar) Sonlanma-DUR (terminasyon) kodonları (UAA, UAG ve UGA), hiçbir amino asidi kodlamazlar. Bu kodonlar polipeptid sentezinin bittiğinin sinyalini verirler (Stop veya nonsense kodonlar) Dejenerasyon: Bir amino asidin birden fazla kodon ile kodlanmasıdır.
 kodonu (AUG), tüm hücrelerde bir polipeptidi başlatan sinyal kodonu (Bir polipeptidin içinde sinyal ayrıca Met’i kodlar) Sonlanma-DUR (terminasyon) kodonları (UAA, UAG ve UGA), hiçbir amino asidi kodlamazlar. Bu kodonlar polipeptid sentezinin bittiğinin sinyalini verirler (Stop veya nonsense kodonlar) Dejenerasyon: Bir amino asidin birden fazla kodon ile kodlanmasıdır.")
8
mRNA amino asit kodları

9
Genetik kodun dejenerasyonu

10
Kodon ve antikodon eşleşmesi

11
Kodon ve antikodon eşleşmesi
tRNA’da bulunan inosinat (I), U, C ve A ile hidrojen bağı yapabilir
, U, C ve A ile hidrojen bağı yapabilir.")
12
Wobble bazı Wobble bazı tRNA’ların birden fazla kodonu tanımalarını sağlar Antikodon: tRNA’da mRNA’ya karşılık gelen kodon. Antikodon’daki ilk baz (5′ 3′ yönünde) wobble bazdır.
 wobble bazdır.")
13
Wobble Hipotezi Bir mRNA kodonundaki ilk iki baz, tRNA’daki antikodon ile her zaman güçlü Watson-Crick baz eşleşmesi yapar. Antikodondaki ilk baz (5′ 3′ yönünde okunur) kodondaki 3. bazın karşısındaki bazdır. tRNA tarafından tanınan kodonların sayısını belirler (Bkz. Tablo) Tüm 61 kodonun translasyonu için minumum 32 tRNA gereklidir.
 kodondaki 3. bazın karşısındaki bazdır. tRNA tarafından tanınan kodonların sayısını belirler (Bkz. Tablo) Tüm 61 kodonun translasyonu için minumum 32 tRNA gereklidir.")
14
Antikodondaki Wobble bazı tRNA’nın tanıyabildiği kodon sayısını nasıl belirler

15
Genetik kod’daki doğal değişiklikler

16
Protein Sentezi için mRNA Ribozomlar tRNA Aminoasitler gereklidir

17
Ribosomes are necessary for translation
Fig.14.1

18
Ribozomal altüniteler
S (Svedberg ünitesi)
")
19
Bakteriyel rRNA

20
tRNA’nın karakteristik yapısal özellikleri bulunur
Birçok tRNA’nın 5′ ucunda guanilat (PG) rezidüsü ve 3′ ucunda, CCA (3′) sekansı vardır. Aminoasit kolu spesifik bir amino asidi taşır. Antikodon kolu, antikodon içerir. TψC kolu, ribotimidin (T), pseudouridin (ψ) içerir. D ve TψC kolları tRNA’nın katlanması için önemlidir.
 rezidüsü ve 3′ ucunda, CCA (3′) sekansı vardır. Aminoasit kolu spesifik bir amino asidi taşır. Antikodon kolu, antikodon içerir. TψC kolu, ribotimidin (T), pseudouridin (ψ) içerir. D ve TψC kolları tRNA’nın katlanması için önemlidir.")
21
tRNA contains unusual bases
Fig.14.2

22
Tüm tRNA’ların genel yonca yaprağı şeklinde sekonder yapıları

23
Cloverleaf Model of tRNA
Transcribed as large precursor cleaved to 4S 5’-Gp pCpCpA-3’ stem and loop structure TyC dihydrouracil Fig.14.3

24
Maya tRNAAla’nın nükleotid sekansı

25
3-D Structure of tRNA Fig.14.4 TyC

26
Maya tRNAPhe’nin 3 boyutlu yapısı

27
Maya tRNAPhe’nin 3 boyutlu yapısı

28
Protein Sentezi Amino asidlerin aktivasyonu İnitiasyon Elongasyon
Terminasyon ve salınım Katlanma ve posttranslasyonel işlemler

29
Protein Sentezi

30
Her bir amino asit için özgül amnioaçil-tRNA sentetaz enzimi vardır
Her bir tRNA’ya doğru aminoasitin tanınıp, bağlanması gerekir. Ayrıca, bu süreç enerji gereksinen bir sentez sürecidir. Bu nedenle, her hücrede her bir aminoasite özgü aminoaçil tRNA sentetaz enzimleri bulunur. Enzim katalizli tepkimede ATP hidrolizi ile aktive edilen amino asit, tRNA’nın 3’ ucundaki adenin bazına ait riboz şekerin 3’-OH grubuna bağlanır.

31
Amino açil tRNA’ların genel yapısı

32
Protein Sentezinde Amino asitlerin aktivasyonu
İki tip amino açil tRNA sentetaz enzimi bulunur. Her amino asidin karbonil grubu peptid bağı oluşumunu kolaylaştırmak için aktive edilir. Her amino asit ile şifrelenen bilgi eşleştirilir. Bu enzimin hata düzeltme görevi de vardır.

33
tRNA doğru aminoasiti tanımalıdır
aminoacyl tRNA synthetases tarafından katalizlenir amino asitler aktive edilir (aminoacyladenylic acid) ATP gerekir Mg 2+ Amino acid + tRNA + ATP aminoacyl-tRNA + AMP + PPi Fig. 14.5
 ATP gerekir. Mg 2+ Amino acid + tRNA + ATP aminoacyl-tRNA + AMP + PPi. Fig")
34
N-formilmetionin

35
Protein Sentezinin Basamakları
Başlama (Initiation) Zincir Uzaması (Elongation) Zincir Sonlanması (Termination)
 Zincir Uzaması (Elongation) Zincir Sonlanması (Termination)")
36
Protein Sentezinin Basamakları-Başlama
Sentezin başlaması için: GTP, IF1, IF2, IF3 gereklidir. İlk olarak SSU rRNA (16S) özgül DNA dizilerini bulur: Shine-Dalgarno sequence (AGGAGG)
 özgül DNA dizilerini bulur: Shine-Dalgarno sequence (AGGAGG)")
37
Başlama için gerekli protein faktörler

38
Shine-Dalgarno dizesi
(5′) AUG ile initiasyon mRNA’da Shine-Dalgarno dizesi ile doğru pozisyondan başlatılır.
 AUG ile initiasyon mRNA’da Shine-Dalgarno dizesi ile. doğru pozisyondan başlatılır.")
39
Shine-Dalgarno dizisi

40
Sentezin başlaması Amino açil veya A bölgesi, P bölgesi,
Bakteriyel ribozomda amino açil tRNA’ların bağlandığı üç bölge vardır: Amino açil veya A bölgesi, P bölgesi, E (çıkış) bölgesi 1. İlk olarak mRNA başlama faktörleri ile birlikte (IF1, 2, 3) küçük alt birime bağlanır. 2. Başlatıcı fMet-tRNA P bölgesindeki mRNA kodonuna bağlanır; IF3 ayrılır. 3. Büyük alt birim komplekse bağlanır; IF1 ve IF2 ayrılır; EF-Tu, tRNA’ya bağlanarak, A bölgesine girişi kolaylaştırır.
 bölgesi. 1. İlk olarak mRNA başlama faktörleri ile birlikte (IF1, 2, 3) küçük alt birime bağlanır. 2. Başlatıcı fMet-tRNA P bölgesindeki mRNA kodonuna bağlanır; IF3 ayrılır. 3. Büyük alt birim komplekse bağlanır; IF1 ve IF2 ayrılır; EF-Tu, tRNA’ya bağlanarak, A bölgesine girişi kolaylaştırır.")
41
Zincir uzaması: İlk basamak (ikinci amino açil tRNA’nın bağlanması”)
İkinci yüklü tRNA, EF-Tu’nun yardımı ile A bölgesine girer; uzamanın ilk basamağı başlar.

42
Uzama, İkinci basamak: Peptid bağı oluşumu
A bölgesinde Dipeptit bağı oluşur (Peptidil transferaz aktivitesi); Yüksüz tRNA, E bölgesine hareket eder ve ribozomu terk eder. Yeni oluşan dipeptit P bölgesine hareket eder. mRNA 3 baz kayar; EF-G translokasyon basamağını kolaylaştırır, uzamanın ilk basamağı tamamlanır.
; Yüksüz tRNA, E bölgesine hareket eder ve ribozomu terk eder. Yeni oluşan dipeptit P bölgesine hareket eder. mRNA 3 baz kayar; EF-G translokasyon basamağını kolaylaştırır, uzamanın ilk basamağı tamamlanır.")
43
Uzama, üçüncü basamak: translokasyon
EF-Tu +GTP + tRNA A bölgesine yerleşir, EF-Tu girişi kolaylaştırır; Peptid bağı oluşumu ile tripeptid elde edilir. yüksüz tRNA, E bölgesine hareket eder. mRNA 3 baz kayar; EF-G translokasyon basamağını kolaylaştırır, uzamanın ilk basamağı tamamlanır.

44
Bakterilerde protein sentezinin terminasyonu
Basamak 1- DUR kodonlarına gelindiğinde, GTP bağımlı salınma faktörlerine “harekete geçme” sinyali verilirve polipeptit zinciri ile tRNA arasındaki bağı kırarak zincirin translasyon kompleksinden ayrılmasını sağlar. Basamak 2- Bu kırılmadan sonra tRNA ribozomdan salınır ve ribozom alt birimlerine ayrılır.

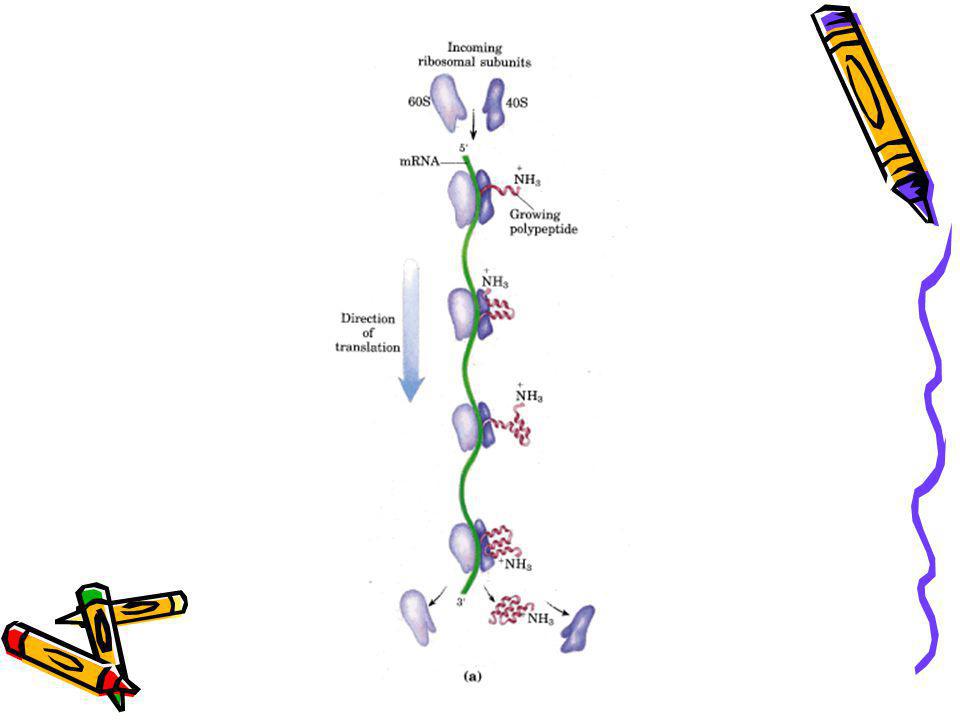
45
Polizom Hem ökaryotik, hem de prokaryotik hücrelerde protein sentezinde 10 ile 100 ribozom aynı anda aktiftir. Bunlara polizom denir.

46
Polizom: Elektron mikroskobu

47
Bakteride transkripsiyon ve translasyonun eşleşmesi

49
Ökaryotlarda Protein Sentezi
Daha komplekstir mRNA stoplazmaya taşınır mRNA 1-2 saatte yıkılır Kozak sequence 5’-ACCAUGG….. AUG Met kodlar Daha fazla sayıda ribozom görev alır Ribozomlar E.R’a tutunur.

50
Ökaryotik initiasyon kompleksinin oluşumundaki protein kompleksler

51
Polipeptitteki aminoasitlerin linear dizilimi primer yapıyı verir.
Fig ; R-groups (radical)
")
52
Amino asitler N terminalden C-terminale doğru peptid bağı ile birleştirilirler

53
Sekonder yapıda, polipeptit zincirinde birbirine komşu olan amino asitlerin oluşturduğu, düzenli ve tekrarlayan bir konfigürasyon bulunur.

54
Proteinin tersiyer yapısı zincirin uzaydaki 3 boyutlu konformasyonunu ifade eder.
Bu yapı uzayda ….. Sisteinler arasındaki disülfid bağları Hidrofilik polar R grupları Hidrofobik nonpolar R grupları ile stabilize edilir

55
Quaternary Structure Birden çok polipeptit zincirinin biraraya gelmesi ile kuaterner yapı oluşur.

57
Translasyon Sonrası Modifikasyonlar
N-ucu ve C-ucundaki a.a ler çoğunlukla uzaklaştırılır yada değişime uğrar. Bazen tek bir a.a değişime uğrayabilir. Bazen karbohidrat yan zincirleri takılabilir Polipeptit zincirlerinde kırpılma yapılabilir. Sinyal dizileri proteinden uzaklaştırılır. Polipeptit zincirleri çoğu kez metallerle kompleks yapar Folding: chaperonins

58
Post-translasyonel modifikasyonlar
Amino terminal ve karboksi terminal uçta modifikasyonlar: Tüm polipeptidler bakterilerde N-formilmetionin, ökaryotlarda ise metionin ile başlar. Formil grubu, bu amino asidler final peptid’den uzaklaştırılır. Ökaryotik proteinlerin % 50’sinde amino terminal uçtaki amino grubu N-asetil’lenir.

59
Post-translasyonel modifikasyonlar
Bazı amino asitlerin modifikasyonu: Ser, Thr ve Tyr rezidülerinin hidroksil gruplarının enzimatik olarak ATPile fosforillenmesi. Böylece diğer moleküllerle iyonik bağ yapabilen eksi yüklü a.a ler oluşur. Bazı proteinlerde Glu rezidüsüne ekstra karbonil grubu eklenmesi (Örn.protrombin’de K vitamininin rolü) Bazı kas proteinlerinde ve sitokrom c’de monometil ve dimetillizin rezidüleri (Örn. calmodulin’de trimetil lizin rezidüsü bulunmakta)
 Bazı kas proteinlerinde ve sitokrom c’de monometil ve dimetillizin rezidüleri (Örn. calmodulin’de trimetil lizin rezidüsü bulunmakta)")
60
Amino asid rezidülerinde modifikasyonlar

61
Amino asid rezidülerinde modifikasyonlar

62
Amino asid rezidülerinde modifikasyonlar

63
Post-translasyonel modifikasyonlar
Karbohidrat yan zincirinin eklenmesi: Polipeptid zincirinin sentezi sırasında veya sonra glikoproteinin karbohidrat yan zinciri eklenir. Bazı glikoproteinlerde karbohidrat yan zinciri enzimatik olarak Asn rezidülerine bağlanır (N-linked oligosakkaridler), diğerlerinde Ser ve Thr residülerine bağlanır (O-linked oligosakkaridler). Birçok protein ekstrasellüler olarak fonksiyon görür.
, diğerlerinde Ser ve Thr residülerine bağlanır (O-linked oligosakkaridler). Birçok protein ekstrasellüler olarak fonksiyon görür.")
64
Post-translasyonel modifikasyonlar
Sinyal sekansının kaybı: Amino terminal uçtaki rezidülük kısım proteinin hücre içi hedeflere yönlendirilmesinde önemli rol oynar. Daha sonra bu kısım çıkartılır.

65
Post-translasyonel modifikasyonlar
Polipeptitlerin metallerle kompleks oluşturması : Proteinlerin tersiyer ve kuaterner yapılarında metal atomları vardır ve bu yapıları kazanmaları çoğu bu kez metallere bağlıdır.

66
Post-translasyonel modifikasyonlar
Prostetik grupların eklenmesi: Birçok protein aktiviteleri için kovalant bağlı prostetik gruplara gereksinim duyarlar. Örn. Asetil-CoA karboksilaz’ın biyotin molekülü ve sitokrom c’nin hem grubu Proteolitik işlemler: Birçok protein sentez sonrası kısaltılır. Örn. İnsulin Disülfid çapraz bağlarının oluşumu: İki sistein rezidüsü arasında çapraz bağ oluşur. Bu proteini hücre içinde ve dışında etkilerden korur.

67
Protein Sentezinin İnhibisyonu

68
Peptid bağ oluşumunu promisin ile blokajı
Translokasyona uğramadığı için polipeptid sentezinde erken terminasyon

69
Peptidil promisin

70
Tetrasiklin, bakterilerde protein sentezini ribozomun A bölgesini
bloke ederek sağlar.

71
Siklohekzimid Sikloheksimid, 80 S ökaryotik
ribozomlarda peptidil transferaz aktivitesini bloke eder fakat 70 S Bakteriyel (ayrıca mitokondrial ve kloroplast) üzerine etkis yoktur.
 üzerine etkis yoktur.")
72
Kloramfenikol Bakterilerde protein sentezini peptidil transferaz blakajı ile yapar. sitozolik ökaryotik protein sentezi üzerine etkisi yoktur.

73
Streptomisin Basit bir trisakkaridtir. Düşük konsantrasyonlarda genetik kodun hatalı okunması, yüksek dozda ise initiasyonun inhibisyonuna sebep olur.

74
Toksinlerle protein sentez inhibisyonu
Difteri toksininin, ökaryotik elongasyon faktörü üzerine etkisi vardır. Risin, ökaryotik ribozomlarda 60 S alt üniteyi inaktive eder (23S rRNA’da spesifik bir adenozin’in depurinasyonu ile).
.")



>")





 ZM106 Biyokimya 11. Hafta.>")








>")


