Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Proteinlerin Moleküler Mimarileri
- Şekilsel olarak proteinin anlaşılması - Oluşum süreci (sarılması) Proteinlerin 3 boyutlu Yapıları
 Proteinlerin 3 boyutlu Yapıları.")
2
Bölümde konu başlıkları
Protein nedir, çeşitli proteinler Peptid bağının oluşması (birincil yapı) Peptid bağının düzlem özelliği Proteinin ana zincirinin belli biçimsel mimarilere kıvrılması (proteinin ikincil yapıları) Polipeptid zincirinin alacağı son biçimsel mimari (üçüncül yapı oluşumu) Proteinlerin dördüncül yapısı ve 4. yapıların x, y z eksen 3 boyut ile ilgisi yoktur
 Peptid bağının düzlem özelliği. Proteinin ana zincirinin belli biçimsel mimarilere kıvrılması (proteinin ikincil yapıları) Polipeptid zincirinin alacağı son biçimsel mimari (üçüncül yapı oluşumu) Proteinlerin dördüncül yapısı ve 4. yapıların x, y z eksen 3 boyut ile ilgisi yoktur.")
3
Protein çeşitleri Proteinler çok çeşitli şekillerde olabilirler
1-Globüler proteinler: Pek çoğu suda çözünür, sıkı bir şekilde sarılmış yumak olarak kabaca küreye benzerler. Globüler (yuvarlağımsı) proteinlerin genel olarak iç kısımları hidrofobik dış kısımları ise hidrofilik özelliktedir. Genel olarak yüzeylerinde oyuk veya çukurluklar vardır ki bunlar özgün olarak başka molekülleri bağlamaya eşlik ederler. Bu sayede biyolojik faaliyetin dinamik etmenleri olarak işlev görürler Globüler proteinlerin çoğu enzim olarak görev yapar E.coli deki proteinlerin %31 metabolik enzim olarak görev yapmaktadır. Diğer globüler proteinler taşıma ve yönetim gibi farklı işlevler üstlenmişlerdir ör. E coli deki proteinlerin %12 si bu tip proteinlerdir
 proteinlerin genel olarak iç kısımları hidrofobik dış kısımları ise hidrofilik özelliktedir. Genel olarak yüzeylerinde oyuk veya çukurluklar vardır ki bunlar özgün olarak başka molekülleri bağlamaya eşlik ederler. Bu sayede biyolojik faaliyetin dinamik etmenleri olarak işlev görürler. Globüler proteinlerin çoğu enzim olarak görev yapar. E.coli deki proteinlerin %31 metabolik enzim olarak görev yapmaktadır. Diğer globüler proteinler taşıma ve yönetim gibi farklı işlevler üstlenmişlerdir ör. E coli deki proteinlerin %12 si bu tip proteinlerdir.")
4
İpliksi Proteinler İpliksi yapıdaki proteinler hücre veya organizmaya mekanik destek sağlayan proteinlerdir. İpliksi proteinler kablo ve ilmikler gibi inşa edilmiş büyük oluşumlardır Ör: saç veya tırnakların temel bileşiği ɑ-keratin Ör: deri, kemik, diş ve tendonların temel bileşiği kollajen

5
Membran Proteinleri 1- Globüler proteinler 2- İpliksi proteinler
Membran proteinleri E coli de en azından polipeptidlerin %16 sıdır Ökaryotlarda daha yüksek oranda membran proteinleri bulunmaktadır.

6
Temel başlıklar Peptid bağının mimarisi
Alfa-Sarmal ve beta-yapraklı biçimler olarak peptid bağı ile oluşturulan yaygın ikincil yapılar Daha üst seviyede protein mimarisi Protein sarılması ve kararlılık Protein mimarisinin protein işlevi ile ilişkisi Ör:Kollajen, hemoglobin, ve antibodiler

7
Protein mimarisi Proteinde yüzlerce aminoasit olabilir
Her bir aminoasidin bir diğerine göre sayısız farklı konumlarda bulunabilir. Sonuçta her bir protein astronomik sayılarda farklı biçimler sergileyebilir. Fakat hücresel koşullar altında (doğal ortam) her bir protein zinciri tek bir biçimsel duruş yada mimariye kıvrılır buna doğal mimari (native conformation, native state)
 her bir protein zinciri tek bir biçimsel duruş yada mimariye kıvrılır buna doğal mimari. (native conformation, native state)")
8
Protein mimarisi Protein zincirindeki aminoasitlerin yan grupları arasındaki H bağları ve diğer zayıf etkileşimler nedeniyle protein zincirinin yerleştiriminde bir çok sınırlamalar vardır. Bir proteinin biyolojik işlevi tamamen onun doğal biçimsel mimarisine (conformation) bağlıdır.
 bağlıdır.")
9
Protein mimarisi Protein tek bir polipeptid zincirinden oluşabileceği gibi birkaç polipeptid zincirinden de oluşabilir Proteini oluşturmak için birkaç polipeptid zinciri bir araya geliyorsa bunlar arasında sadece zayıf bağlar söz konusudur Kural olarak, her bir polipeptid tek bir genden oluşur Genin büyüklüğü ile proteinin büyüklüğü birbirinde 10 katı farklı olabilir. Bazı polipeptidler yanlızca 100 aminoasitten oluşabileceği ve yaklaşık molekül ağırlığı olabileceği gibi bazıları 250, , 000 gibi olabilir.

10
Protein mimarisi Bazı türlerde her bir polipeptidin büyüklüğü ve aminoasit sıralaması genomdan belirlenebilir. Escherichia coli bakterisinde yaklaşık 4000 farklı polipeptid zinciri vardır ve her bir polipeptidin ortalama 300 aminoasidi vardır Meyve sineği, Drosophila melanogaster, yaklaşık farklı ve 300 aminoasitlil polipeptid molekülü içerir İnsan ve diğer memeliler yaklaşık farklı polipeptid içerir Bir hücrenin oluşturacağı tüm polipeptidleri inceleyen alan Proteomiks olarak anılmaktadır.

11
Protein mimarisi: Neden önemli
Aminoasit zinciri doğrusal olarak birbirlerine bağlanan düz zincirli bir yapı Ama bu düz zincir düz uzanıp durmuyor. Kıvrılıp yumak oluşturuyor, Yumak ama herhangi bir yumak değil özgün bir yumak; Bu yumak nasıl oluşuyor? Bana ne? Nasıl oluştuğunu anlamak neden önemli?

12
Protein mimarisi: Neden önemli
Proteine atfedilen tüm özellikler onun biçimsel mimarisi ile ilgilidir. Biçimsel mimarideki değişme ölümle yaşam arasındaki canlı ile cansız arasındaki fark kadar önemlidir. Çiğ yumurta ile haşlanmış yumurta arasındaki tek fark proteinlerin biçimsel mimarilerinin değişmesidir. Haşlanmış yumurtadan civciv çıkar mı?

13
Protein mimarisini oluşturan bağlar
Protein moleküler mimarisinde sadece peptid bağı ile birbirine bağlanan proteinler arasında kovalent bağ vardır. Protein mimarisini kararlı halde tutan güç veya bağların hepsi kovalent olmayan bağlardır. (disülfit bağları hariç) 2., 3. ve 4. yapıların oluşturulması çoğunlukla zayıf etkileşimlere bağlıdır.
 2., 3. ve 4. yapıların oluşturulması çoğunlukla zayıf etkileşimlere bağlıdır.")
14
Birincil yapı Bir polipeptid zincirindeki aminoasitlerin peptid bağı ile oluşturdukları zincirin diziliş sırası ve sayısı proteinin birincil yapısıdır.

15
İkincil mimari Peptid bağının bir diğer peptid bağı ile oluşturduğu H bağları ile oluşturulan bölgesel özgün biçimler

16
Üçüncül mimari

17
3.cül mimari Üçüncül yapılar tamamıyla sarılmış sıkı yumak hali ifade eder Bir polipeptidin sarılmış halinde çeşitli bölümler birbirinden ayrı bölümler gösterir bunlara domain denir. Üçüncül yapılar aminoasit zincirinde birbirlerine komşu olmayan aminoasit yan gruplarının etkileşimi ile kararlı hale getirilir. Üçüncül yapının oluşumu esnasında aminoasit zincirinin farklı bölümleri ve çeşitli ikincil yapılar bir araya getirilir

18
Dördüncül mimari 1. polipeptid 2. polipeptid

19
Dördüncül mimari Hemoglobin: 4 polipeptid

20
Dördüncül mimari Hemoglobin: 4 polipeptid

21
Protein Moleküler mimarisini belirleme yöntemleri
1.cil yapı-kimyasal yöntemle Edman ajanı ile veya gen ile belirlenebilir Protein biçimsel mimarisi ise genellikle X-ışınları yardımıyla belirlenir.

22
X-ışınları Yansımaları
Bu yöntemde paralel X ışınları protein kristaline gönderilir Kristale çarpan elektronlar yansır ve yansıyan elektronalr bir film üzerinde belirir Atomu saran elektronların yoğunluğundan oluşan yansımalar matematiksel analiz edilir Atomu saran elektron yoğunluğu molekülün şekli ve farklı atomların birbirlerine göre konumları hakkında bilgiler verir Bu bilgiler kimyasal bağ bilgileri ışığında değerlendirilir. Bu sayede Molekülün üç boyutlu biçimi ve bağları çıkarılır Bu yöntemle aminoasit zincir yapısı bilinmeksizin proteinin biçimsel mimarisi elde edilebilir.

23
X-ışınları Yansımaları
X-ışınları yansımaları ile önceleri sadece basit tekrarlanan ünitelerden oluşan iplikis proteinler ve küçük biyolojik moleküllerin biçimleri aydınlatıldı. Enzimlerde globüler proteinlerde karmaşık yapılardan dolayı atomların konumlarını hesaplamak oldukça zor idi. Yapılması gereken hesaplamaları yapmak insan yeteneklerini aşıyor, bilgisayalrlar gerekiyordu. Bilgisayar teknolojisi ilerledikçe protein yapısı daha kolay ve detaylı aydınlatılır oldu. 1962 yılında Miyoglobin ve Hemoglobin’in yapısı aydınlatıldı

24
İnsülin kristali

25
Protein kristali Bir molekül yeterince saf ise birbirlerine göre kendilerini konumlandırarak yüzlercesi bir araya gelerek düzenli belli bir biçimde bir araya gelirler. Saflık, Sulu çözeltiden yavaş bir şekilde çökmeye zorlama Kristaller birbirlerine kovalent olmayan bağlarla bağlanırlar X ışınları ile protein kristallerinin biçimsel mimarileri aydınlatılabilir

26
Protein mimari kararlılığı
Protein zinciri var olan ortamında en düşük enerjili mimari biçime kıvrılmaya çalışır. En düşük enerjili mimari biçim? -kendi içinde azami sayıda bağ oluşturma -çözgenle (suyla) azami sayıda olumlu etkileşim (hidrojen bağı, iyon-dipol) -su sevmeyen grupların sudan uzaklaştırılması (hidrofobik etkileşim)
 azami sayıda olumlu etkileşim (hidrojen bağı, iyon-dipol) -su sevmeyen grupların sudan uzaklaştırılması (hidrofobik etkileşim)")
27
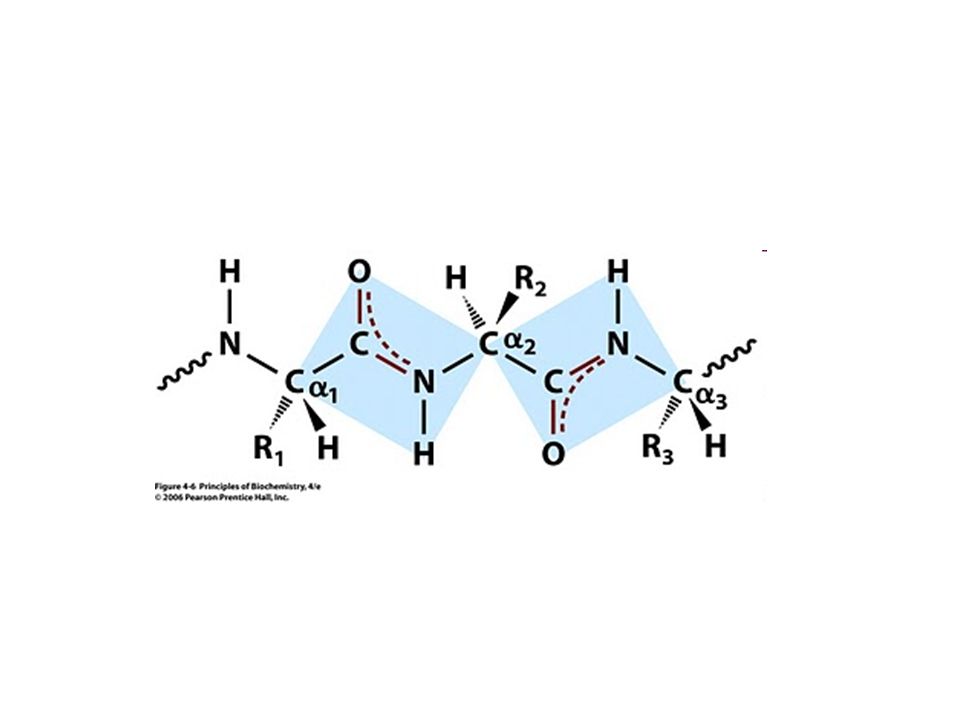
Protein mimarisi: Peptid Bağının özelliği
Proteinin mimarisini anlamak için peptid bağının özelliklerini anlamak gereklidir. X-ışınları yansımalarından protein yapısını çözmek zordu.. (globüler proteinler) İnsanlar çeşitli modeller üzerinde düşünerek protein molekülünün nasıl durabileceğini canlandırmaya çalışmışlar
 İnsanlar çeşitli modeller üzerinde düşünerek protein molekülünün nasıl durabileceğini canlandırmaya çalışmışlar.")
28
Protein mimarisi: Peptid Bağının özelliği
Protein zinciri mevcut ortamında en düşük enerjili mimari biçime kıvrılmaya çalışır. Protein zincirindeki atomlar istediği yere dönme özgürlüğüne sahip mi? Tüm gruplar tekli kovalent bağ

29
Protein mimarisi: Peptid Bağının özelliği
1-Çift bağa göre uzun Beklenenden uzun Linus Pauling and Robert Corey, deneyleri Aminoasit, dipeptid, tripeptid kristallerinin X ışınları yansımaları 2-Tekli bağa göre kısa Beklenenden kısa

30
Protein mimarisi: Peptid Bağının özelliği
1-Karbonil C=O bağı ve C-N amid bağı kısmi çift bağ özelliği göstermektedir. 2-Trans biçim. Tüm bunlar ne anlama geliyor.. - 2,8 Ao + Peptid bağının trans biçim olduğunu göstermektedir Kısmi çift bağ

31
Protein mimarisi: Peptid Bağının özelliği
Kısmi çift bağ demek kısmi iyon oluşturma yani yüklü olma demektir. Çift bağın dönme özgürlüğü yoktur. Trans biçimli duran O, C, N ve H atomlarının konumları sabittir, bir düzlem oluştururlar. Düzlemin iki köşesinde kısmi + ve – yükler vardır bunlar ne anlama geliyor.. - 2,8 Ao + Peptid bağının trans biçim olduğunu göstermektedir Kısmi çift bağ

32
Protein mimarisi: Peptid Bağının özelliği

33
Protein mimarisi: Peptid Bağının özelliği
Protein zincirindeki peptid düzlemleri bir bütün olarak hareket eder. Düzlemler sadece ɑ-karbon atomları etrafında dönebilirler. Peptid düzlemleri yanyana veya üstüste geldiklerinde kendi aralarında H-bağı oluşturabilirler. Bu sayede protein kendi içinde oluşturduğu zayıf bağlarla kararlılık kazanır. 2,8 Ao Normal VDW mesafesi 3,5 A Peptid bağının trans biçim olduğunu göstermektedir Kısmi çift bağ

34
Protein mimarisi: Peptid Bağının özelliği

36
Linus Pauling’in çizimi

38
Pauling and Corey Peptid düzlemleri ile protein zincirinden H-bağlarını azamiye çıkaracak yani en kararlı duruşları oluşturacak yapıların modellerini oluşturmaya başladılar. Düşündükleri şekillerden biri, peptid düzlemlerinin kendi içinde kıvrılıp dönerek oluşturdukları yapıydı. Alfa-sarmal yapıyı elde ettiler.

39
Tüm peptid düzlemleri H bağı yapıyor

40
Protein mimarisi: Peptid Bağının özelliği
Kağıt üzerinde elde edilen biçimleri, şayet doğruysa X-ışınları yansımalarında bulmaya çalıştılar ve pek çok olduğunu gördüler… X-ışınları yansıması filmi elde edilmiş bir çok proteinde -sarmal yapının olduğunu görmüşlerdir.

41
Linus Pauling:-sarmal yapı
-sarmal biçimi açıklaması nedeniyle Linus Pauling 1954 yılında Nobel ödülüne layık görüldü.

42
Hemoglobin: %80 -sarmal biçim

43
-Sarmal Biçimin özellikleri
Her dönüşte ortalama 3,6 aminoasit içerir Her peptid düzlemi sarmalın eksenine paralel yerleşmiştir. Her peptid düzlemi zincirin her iki yönünde 3.aminasitle H-bağı yapmıştır. H-bağları sarmal eksenine paraleldir. Sarmalın içine hiçbir molekül sığmayacak ölçüde dardır. Aminoasitlerin yan grupları sarmalın dışında kalmaktadırlar

44
-Sarmal Biçimin özellikleri
-sarmal biçimde çok sıkı bir paketleme söz konusudur Proteine esnek olmayan, dar, uzun, çubuksu bir özellik kazandırır. Proteinlerde %5-80 arasında -sarmal yapılar bulunur. Proteinlerdeki oranı ortalama %26 dır Ör: miyoglobinde aminoasitlerin %80’i 8 tane sarmal yapı içerisinde yer alır. Kas: miyosin %80 sarmal biçim içerir. Sarmal yapıda 4-5 – 40 arasında ortalama 12 aminoasit içerir

45
-Sarmal Biçimin özellikleri
-sarmal biçim kendi içinde H-bağı oluşturduğundan peptid bağının çözücü ile etkileşimi sınırlandırılmıştır. -sarmal yapının oluşmaya başlayabilmesi için en azından ardışık 3 aminoasidin yan gruplarının birbiri ile uyumlu olması lazım. Kolaylıkla oluşmaya başlamaz, ama oluştuktan sonra oldukça kararlı biçimsel bir mimaridir.

46
Sarmal yapı ve aminoasitler
Bazı aminoasitler daha sıklıkla -sarmal biçimde yer alırlar Ör.Alanin, küçük ve yüksüz yan grubu ile biçime kolaylıkla uyar, ve sarmal yapıda oldukça yaygındır. Tirozin ve asparajin gibi büyük yan gruba sahip aminoasitler pek sık bulunmaz. Glisin, yan grubunda H olduğundan her yöne kolaylıkla döner o nedenle sarmal yapıyı teşvik etmez, hatta bozar, pek çok sarmal yapı glisinden sonra başlar veya glisinle biter Prolin, yan grup peptid zincirine bağlandığından Amino grubu H bağı yapamaz, sarmal yapıda bulunma olasılığı en düşük aminoasittir. Hatta doğrudan sarmal yapıyı bozucu etkiye sahiptir. Prolin sıklıkla sarmal yapının bitiminde yer alır, proteini dönmeye zorlar.

47
Sarmal yapı özellikleri
Çoğu Sarmal biçimde, polar yan gruplar sarmalın bir yüzünde, hidrofobik aminoasitler diğer yüzünde toplanma eğilimi göstermektedir ki böyle sarmallara, amfoter sarmal denir. Amfoter sarmalın polar yüzü çözücüyle etkileşim halinde iken, hidrofobik yüzü protein içine doğru yönelmiştir. Tüm zincirin yönünü etkilediğinden buna sarmal direksiyonu denir.

48
Proteinde sağ el yönünde dönen sarmal yapı bulunur

49
İkincil Mimari: -biçimleri (pileli şerit) biçimi (-pleated sheat)
Pauling ve Corey, -sarmal yapıdan sonra, beta-pileli zigzag biçimlerinin modellerini önermişlerdir. Düz bir biçimde uzanmış bir aminoasit zincirleri arasında H bağı oluşumunu ile oluşan biçimlerdir. peptid düzlemleri arasında azami sayıda H-bağı oluşmaktadır.

50
Paralel -biçimleri Zıt yönlü -biçimleri

51
Zıt yönlü -biçimleri

52
Zıt yönlü -biçimleri

53
Paralel -biçimleri

54
-biçimleri Proteinlerde ikinci yaygın biçimsel mimarilerdir.
Her zaman birkaç tanesi beraber bulunur. Yani tek başlarına uzanmış bir -biçimi bulunmaz. (Tek başına uzanmış bir aminoasit zinciri, zincire kararlılık kazandırmaz. H bağı yapılabilmesi için daima birkaç tanesi beraber bulunmalıdır) Tipik bir -pileli biçim 2 ile15 şeritin yan yana gelmesiyle oluşur Her bir şerit, genelde 6 aminoasit (3-10 arasında) uzunluğundadır Paralel olmayan - pileleri daha kararlıdır (paralellere kıyasla) muhtemelen doğrusal olmayan H bağlarından dolayı. Burada kaldık
 Tipik bir -pileli biçim 2 ile15 şeritin yan yana gelmesiyle oluşur. Her bir şerit, genelde 6 aminoasit (3-10 arasında) uzunluğundadır. Paralel olmayan - pileleri daha kararlıdır (paralellere kıyasla) muhtemelen doğrusal olmayan H bağlarından dolayı. Burada kaldık.")
55
-biçimleri Bazı proteinler tamamen -şeritli yapılardan oluşur,
Genelde proteinlerdeki oranları düşüktür. Şeritin proteinin içine bakan tarafı genelde hidrofobik iken dışa bakan tarafı hidrofiliktir Paralel -pilelerinin genelde her iki tarafı da hidrofobiktir ve proteinin iç kısımlarında gömülmüş durumdadır

56
Polar yan gruplar yüzeye doğru yönelimiş
Şeritin proteinin içine bakan tarafı genelde hidrofobik iken dışa bakan tarafı hidrofiliktir İç kısımda bir araya gelmiş Hidrofobik yan gruplar Polar yan gruplar yüzeye doğru yönelimiş

57
Paralel -şeritleri (1ADF)
At karaciyeri alkol dehidrojenaz enzimi [PDB 1ADF].

58
Paralel -şeritleri At karaciyeri alkol dehidrojenaz enzimi
[PDB 1ADF].

59
At karaciyeri alkol dehidrojenaz enzimi [PDB 1ADF].
Glisin Lisin Triptofan Treonin
![At karaciyeri alkol dehidrojenaz enzimi [PDB 1ADF].](http://slideplayer.biz.tr/slide/8846057/26/images/59/At+karaciyeri+alkol+dehidrojenaz+enzimi+%5BPDB+1ADF%5D..jpg "Glisin. Lisin. Triptofan. Treonin.")
60
Diğer biçimler: Göze ve Dönüş
Proteinlerde tekrarlanmayan belli biçimlere göze ve dönüş (loop, turn) gibi adlar verilmektedir Bu bölümlerde de peptid bağı çeşitli bağlanmalar yapmaktadır, sıkışıktır Genelde sarmalı takiben görülür Göze ve dönüşler -sarmalları veya -biçimlerinin birinin bitip diğerinin başladığı bölümlere verilen adlardır. Yani onları birbirlerine bağlarlar Bu sayede yumağımsı bir sarılma elde edilir Genelde proteindeki aminoasitlerin 1/3-1/10’u bu tür biçimlerin içinde yer alır Gözeler genelde hidrofilik aminoasitler içerir, ve proteinin yüzeyine çıkarlar, ve su ile H bağı oluştururlar Gözeler şayet birkaç tane (en fazla 5) aminoasit içeriyorlarsa ve proteinde aniden dönüşe neden oluyorlarsa Dönüş olarak adlandırılır. Dönüşlerin en yaygınları zıt yönlü dönüşlerdir ki -Dönüşleri olarak adlandırılır.
 gibi adlar verilmektedir. Bu bölümlerde de peptid bağı çeşitli bağlanmalar yapmaktadır, sıkışıktır. Genelde sarmalı takiben görülür. Göze ve dönüşler -sarmalları veya -biçimlerinin birinin bitip diğerinin başladığı bölümlere verilen adlardır. Yani onları birbirlerine bağlarlar. Bu sayede yumağımsı bir sarılma elde edilir. Genelde proteindeki aminoasitlerin 1/3-1/10’u bu tür biçimlerin içinde yer alır. Gözeler genelde hidrofilik aminoasitler içerir, ve proteinin yüzeyine çıkarlar, ve su ile H bağı oluştururlar. Gözeler şayet birkaç tane (en fazla 5) aminoasit içeriyorlarsa ve proteinde aniden dönüşe neden oluyorlarsa Dönüş olarak adlandırılır. Dönüşlerin en yaygınları zıt yönlü dönüşlerdir ki -Dönüşleri olarak adlandırılır.")
61
Tip I ve II -Dönüşler Her ikisinde 4 aminoasit içerir
Tip II -Dönüş Tip I -Dönüş Her ikisinde 4 aminoasit içerir İlk aminoasit ile 4. arasında H bağı vardır Protein zincirini aninden (180o) döndürürler Genellikle prolin ve/veya glisin içerirler
 döndürürler. Genellikle prolin ve/veya glisin içerirler.")
62
Protein mimarisi: Üçüncül Biçim

63
Protein mimarisi: Üçüncül Biçim
Zaten oluşmuş ve oluşmakta olan belli ikincil yapılar etrafında bir polipeptid zincirinin kıvrılıp, sarılıp son halini aldığı biçim, üçüncül mimari olarak adlandırılır. Üçüncül mimari proteinin doğal mimarisidir, doğal olarak sahip olduğu moleküler mimaridir. Doğal mimari: bir polipeptid zincirinin her bir atomunun uzaydaki konumu (koordinatları) belirlidir. Aminoasit dizilişinde birbirinden çok uzaklarda bulunan aminoasitler üçüncül yapıda yan yana getirilmiş olabilir. İkincil yapılar peptid düzlemleri arasında oluşturulan H bağları ile elde edilen belli biçimli kararlı yapılar iken, Üçüncül yapılarda yan gruplar arası etkileşimin esastır,
 belirlidir. Aminoasit dizilişinde birbirinden çok uzaklarda bulunan aminoasitler üçüncül yapıda yan yana getirilmiş olabilir. İkincil yapılar peptid düzlemleri arasında oluşturulan H bağları ile elde edilen belli biçimli kararlı yapılar iken, Üçüncül yapılarda yan gruplar arası etkileşimin esastır,")
64
Üçüncül Mimari: Yan Gruplar…
Üçüncül biçimin oluşumunun sürücü gücü, en düşük enerjili mimari duruşu sağlamaktır. Bunun için, su seven grupları suyla etkileme getirme ve H-bağı oluşturmalarını sağlama, su sevmeyen grupları sudan uzaklaştırma ki su kendi arasında H bağı oluşturabilsin ve kendi aralarında VDW bağları oluşturabilsinler Sonuçta protein oldukça sıkı ve yoğun bir yumak oluşturur

65
Üçüncül Mimari: Yan Gruplar…
Protein zincirini bir arada tutan ikincil ve üçüncül yapıların hepsi zayıf etkileşimlerdir. Tek birisi hariç… Protein üçüncül mimarisinde kovalent olan tek bağ, Disülfit köprüleridir. Disülfit köprüleri proteinin birincil yapısının parçası değildir, protein sarılırken veya sarıldıktan sonra oluşur.

66
Üçüncül Mimari: Yan Gruplar…
Proteinin iç kısımlarında su vb bulunmaz Yüklü yan gruplar genelde yüzeyde yer almışlardır. Şayet iç kısımlarda bulunuyorlarsa, ya iyon köprüsü vb bir yapı içerisindedirler. Yüklü grupları suda uzaklaştırıp protein içinde sürükleyecek enerji genelde molekülde yoktur Proteinin yüzeyindeki grupların suyla olan etkileşimlerinin sayısı ve gücü proteinin çözünürlülüğünü belirler

67
Örme kazaktaki desenlerle protein arasında bir ilişki var mı?

68
Protein mimarisindeki daha büyük desenler: Motifler
Daha geniş ikincil yapılar, motifler Bunlar -sarmallar ve -biçimleri ve gözelerin karışımdan oluşan biçimlerdir. Bazen bu tür yapıların bazı proteinlerde özel görevleri vardır. Sarmal-Göze-Sarmal motifi

69
Protein mimarisindeki daha büyük desenler: Motifler

70
Protein mimarisindeki daha büyük desenler: Motifler

71
Protein mimarisindeki daha büyük desenler: Motifler

72
Protein mimarisindeki daha da BÜYÜK bölümler: Domain

73
Protein mimarisindeki daha da BÜYÜK bölümler: Domain

74
Protein mimarisindeki daha da BÜYÜK bölümler: Domain

75
Protein mimarisindeki daha da BÜYÜK bölümler: Domain
Proteinler birbirinden ayrı, bağımsız sarılan, sıkı biçimler içerirler bunlara Domain denir. Bunlar motiflerin kombinasyonundan oluşabilir. Domain aa den 300 aa e kadar olabilirler Domainler genelde gözelerle birbirine eklenmiştir.

76
Protein mimarisindeki daha da BÜYÜK bölümler: Domain

77
Domain: biçim ~ işlev Sıklıkla bir domainin belli bir işi vardır
Ör: bir molekülü bağlama, tek bir tepkimeyi katalizleme Çok işlevli enzimlerde, katalizler birkaç domainden biri ile gerçekleştirilir. Genellikle küçük bir molekül bağlanması veya aktif merkezin oluşturulması iki domainin arafazında gerçekleştirilir. Buralar genelde oyuklar,cepler, çukurluklar gibi protein yüzeyindeki yerlerdir. Bağlanma yeri proteinin içinde doğru yönlenmiştir ve sudan uzaktır. Enzime substrat bağlandığı zaman , o kadar uyum sağlanır ki kalan birkaç su molekülü de uzaklaşır.

78
Çeşitli proteinler: üçüncül biçimleri ve işlevleri

79
Çeşitli proteinler: üçüncül biçimleri ve işlevleri
Tek bir -helix domaini

80
Çeşitli proteinler: üçüncül biçimleri ve işlevleri
İki adet -barrel biçiminden oluşur Karonhidrat bağlayan bir proteindir. Tek bir domain ve -sandeviç biçiminden oluşur

81
Çeşitli proteinler: üçüncül biçimleri ve işlevleri
Geniş bir -barreli ve içinde tek bir -sarmal biçim var

82
GFP

83
GFP

84
Green Fluorescent Protein GFP
GFP 238 amino asitten oluşan bir protein, mavi ışık (395 nm) verildiğinde yeşil floresan (509 nm) ışık veriyor. Hücre biyolojisinde işaretleyici protein olarak hücre içi olayları incelemekte kullanılmaktadır. Gen aktarımı ve aktarılan genin çalışıp çalışmadığını gösterici işaretleyici etken olarak kullanılmaktadır Herhangi bir canlı, veya belli hücrelere aktarılabilmekte. Bir çok canlı türüne aktarıldı (bacteria, yeast and other fungi, fish (such as zebrafish), plant, fly, and mammalian cells, including human. Martin Chalfie, Osamu Shimomura, and Roger Y. Tsien were awarded the 2008 Nobel Prize in Chemistry on 10 October 2008 for their discovery and development of the green fluorescent protein.
 verildiğinde yeşil floresan (509 nm) ışık veriyor. Hücre biyolojisinde işaretleyici protein olarak hücre içi olayları incelemekte kullanılmaktadır. Gen aktarımı ve aktarılan genin çalışıp çalışmadığını gösterici işaretleyici etken olarak kullanılmaktadır. Herhangi bir canlı, veya belli hücrelere aktarılabilmekte. Bir çok canlı türüne aktarıldı (bacteria, yeast and other fungi, fish (such as zebrafish), plant, fly, and mammalian cells, including human. Martin Chalfie, Osamu Shimomura, and Roger Y. Tsien were awarded the 2008 Nobel Prize in Chemistry on 10 October 2008 for their discovery and development of the green fluorescent protein.")
85
GFP

86
GFP

87
GFP

88
Çeşitli proteinler: üçüncül biçimleri ve işlevleri

89
Protein Mimarisi: 4.cül biçim
Birden fazla polipeptidin varlığında elde edilen biçime verilen addır. Oligomer de denir Her bir polipeptide alt birim (subunit) veya monomer denir. Oligomerik bir proteinde monomerler birbirinin aynı olabildikleri gibi farklı da olabilir. Aynı alt birimlerden oluşuyorlarsa genelde ikili (dimer) veya 4 lü (tetramer) oluşumları yaygındır Monomerler ayrı birer polipeptid zincirleridir, ayrı ayrı sarılırlar sonrada birleşirler. Oligomer, monomerlerin birleşimi ile kararlılık kazanır.
 veya monomer denir. Oligomerik bir proteinde monomerler birbirinin aynı olabildikleri gibi farklı da olabilir. Aynı alt birimlerden oluşuyorlarsa genelde ikili (dimer) veya 4 lü (tetramer) oluşumları yaygındır. Monomerler ayrı birer polipeptid zincirleridir, ayrı ayrı sarılırlar sonrada birleşirler. Oligomer, monomerlerin birleşimi ile kararlılık kazanır.")
90
Protein Mimarisi: 4.cül biçim
Monomerler birbirlerinden farklılarsa genelde farklı işlevleri vardır Alt birimler yunanca harflerle adlandırılır Ör: 2 olarak adlandırılan bir oligomerik proteinde 2 tane aynı 2 farklı 4 monomer olduğu anlaşılır Monomerler arasında tıpkı 3. biçimde olduğu gibi yan gruplar arasında zayıf bağlarla bağlanmışlardır. Genelde: hidrofobik etkileşim söz konusudur, ayrıca elektrostatik çekim de olabilir. Aradaki bağlar zayıf olduğundan laboratuarda monomerler birbirinden ayrılabilir.

91
Oligomerik Proteinler
İki aynı monomer

92
Oligomerik Proteinler
İki aynı altbirim tamamen -biçimlerinden oluşmuş, simetrik Sadece gözelerinden zayıf bağlarla birbirine bağlanmış

93
Oligomerik Proteinler
İki aynı monomer (dimer) Membran proteini
 Membran proteini.")
94
Oligomerik Proteinler
Trimer aynı subunit Gözelerden birbirine tutunmuşlar

95
Oligomerik Proteinler
Tetramer iki çeşit monomer içeriyor 22

96
Protein Mimari biçimleri: Özet
1.cil yapı: peptid bağı ile kovalent bağlı düz aminoasit zinciri 2.cil yapı: peptid düzlemleri arası azami sayıda H bağı oluşturulması ile elde edilen belli biçimler 3.cül yapı: Yan gruplar arası etkileşim -H bağları -hidrofobik etkileşim -dipol-iyon etkileşimleri -İyon-iyon etkileşimleri -VDW bağları -Disülfit bağları 4.cül yapı: Oligomerik proteinleride monomerler arası etkileşimler

Benzer bir sunumlar


 ile karakterize edilir. Verilen bir elementin tüm atomlarında.>")






 Elementler :>")










>")