Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Introduction to Genetic Analysis
Anthony Griffiths • Susan Wessler • Richard Lewontin William Gelbart • David Suzuki • Jeffrey Miller Introduction to Genetic Analysis Eighth Edition Chapter 9: Proteins and Their Synthesis Copyright © 2005 by W. H. Freeman & Company

2
Gen dizisi ve protein ilişkisi Genetik kod neden dejeneredir?
Doğru aa nasıl mRNAdan okunur? Neden doğru aa tRNA bağlanması protein sentezi için önemlidir? rRNA neden anahtar rol oynar? Translasyon pro ve ökaryotlarda neden farklıdır? Translasyon sonrası modifikasyon neden proteinlerin işlevi için önemlidir? X-ışını kristalografi çalışmalarından elde edilen Haloarcula marismourti bakteri ribozomu yüzeyi. Mvi:RNA içeren yapı, mor:protein, E,P, A bölgeleri beyaz kırmızı, sarı

3
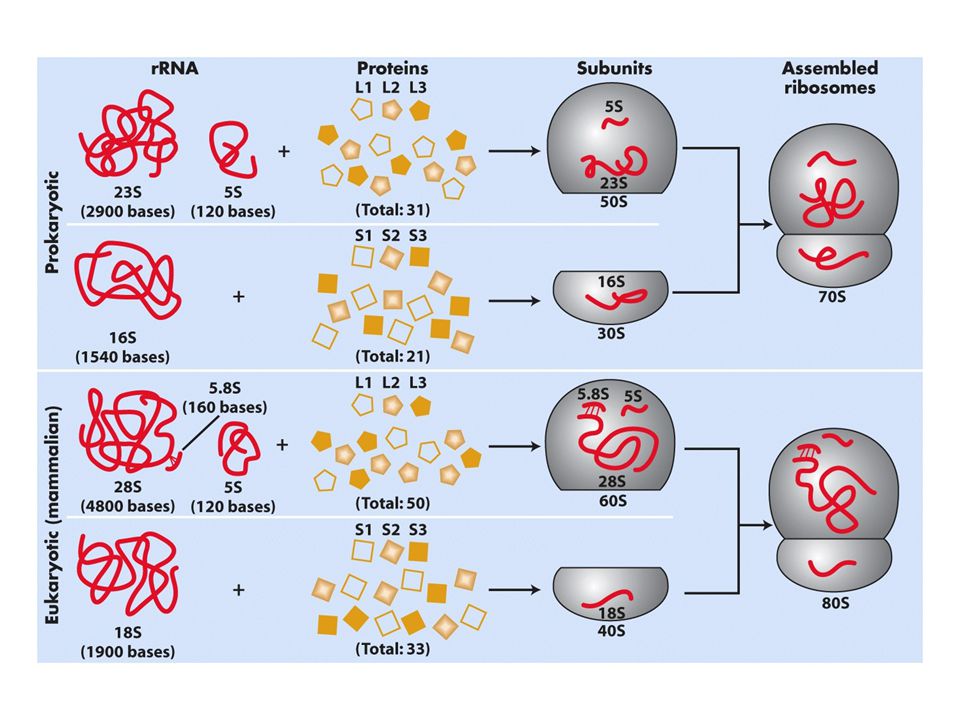
Giriş Bu bölümde bilgi transferinin son evresi incelenecek.translasyon (RNA dizisinden protein sentezi Fonksiyonel RNA molekülleri translasyon sırasında kilit rol oynarlar. tRNA (transfer RNA): adaptör moleküllerdir mRNAda bulunan 3lü nt kodonlarına karşılık gelen amino asitleri ribozoma taşırlar translasyon makinesinin temel bileşenidir rRNA (ribozomal RNA) ribozomun en büyük bileşenidir. Ribozomlar büyük makromoleküler komplekslerdir ve aminoasitleri biraraya getirerek proteinleri oluştururlar. Bölünen bir hücrede rRNA ve tRNA toplam RNA’nın %95ini oluşturur mRNA ise sadece %5.İki etken bu bolluğu açıklayabilir 1.mRNA’dan daha stabildirler bunedenle uzun süre hücrede kalırlar 2.aktif bir ökaryotik hücrede toplam çekirdek transkrpsiyonunun yarısından fazlası rRNA ve tRNA genlerinden oluşur
: adaptör moleküllerdir mRNAda bulunan 3lü nt kodonlarına karşılık gelen amino asitleri ribozoma taşırlar translasyon makinesinin temel bileşenidir. rRNA (ribozomal RNA) ribozomun en büyük bileşenidir. Ribozomlar büyük makromoleküler komplekslerdir ve aminoasitleri biraraya getirerek proteinleri oluştururlar. Bölünen bir hücrede rRNA ve tRNA toplam RNA’nın %95ini oluşturur mRNA ise sadece %5.İki etken bu bolluğu açıklayabilir. 1.mRNA’dan daha stabildirler bunedenle uzun süre hücrede kalırlar. 2.aktif bir ökaryotik hücrede toplam çekirdek transkrpsiyonunun yarısından fazlası rRNA ve tRNA genlerinden oluşur.")
4
Pro ve Ökaryotlarda Translasyon: genel görünüm
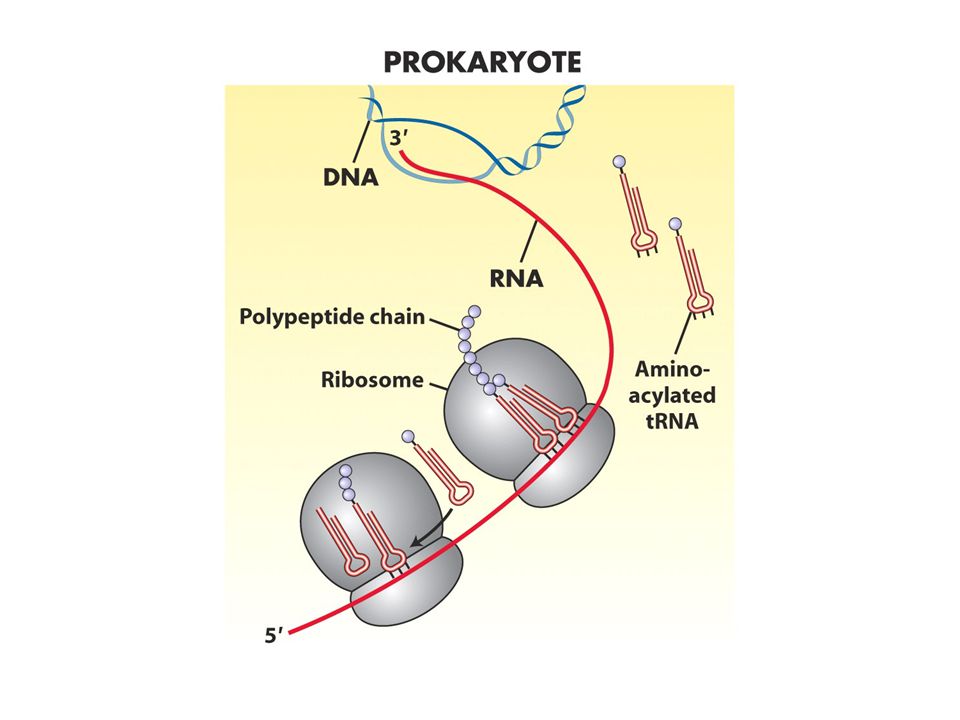
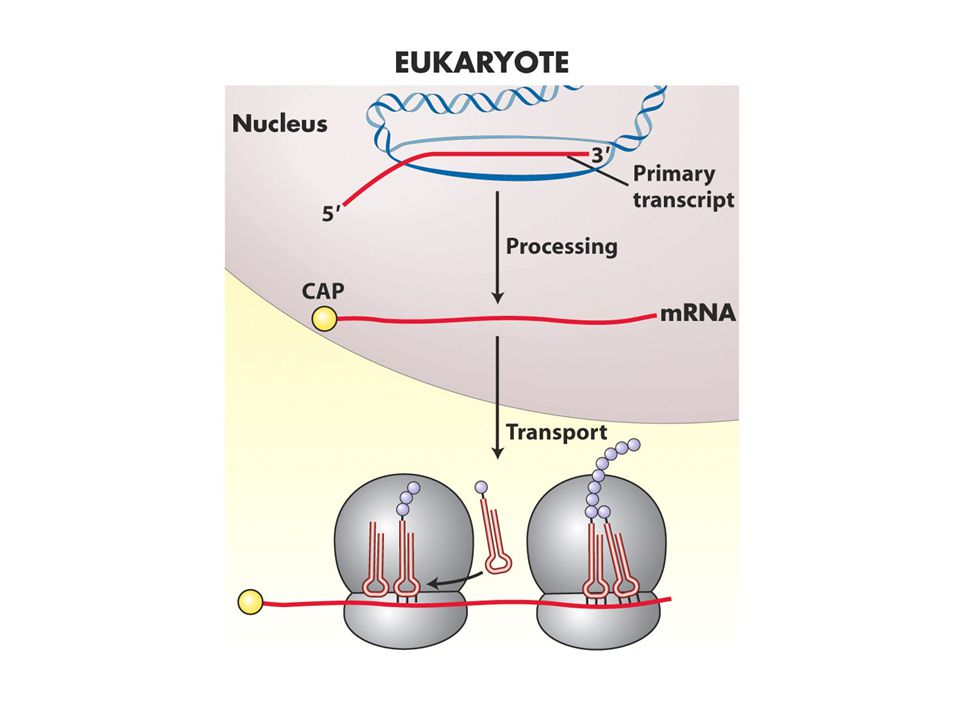
Translasyonel aygıt heriki hücre tipinde birbirine benzer ama ayrıldıkları noktalar vardır Prokaryotlarda transkripsiyon ve translasyon ardıldır: transkripsiyon bir yandan sürerken translasyon 5’ ucunda başlar. Ökaryotlarda ise hücrenin farklı yerinde olur transkript önce işlenir sonra sitoplazmaya geçer

7
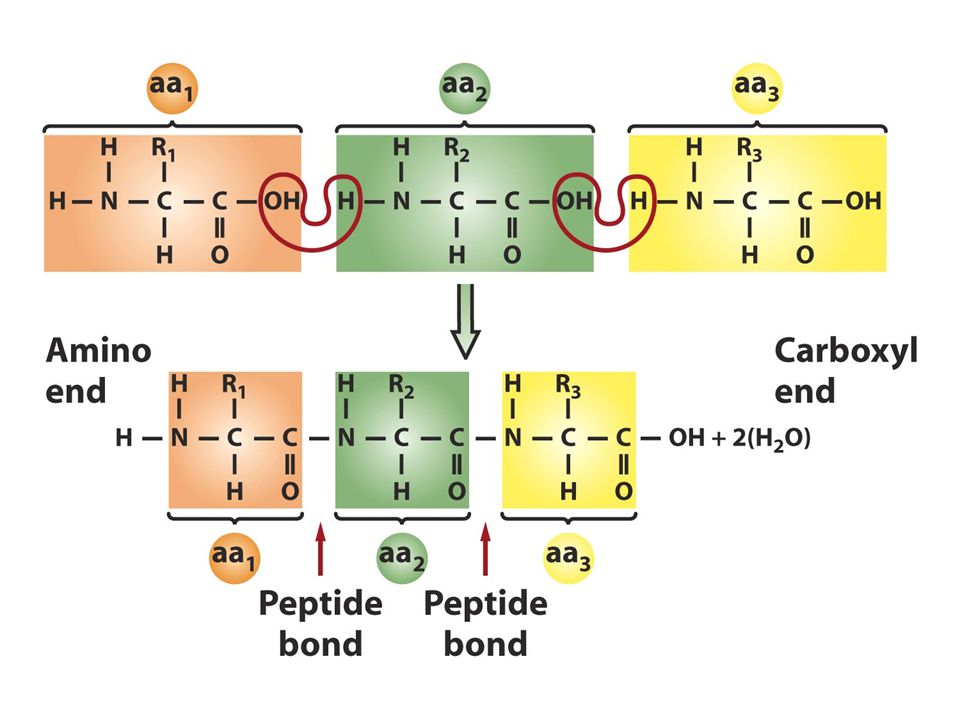
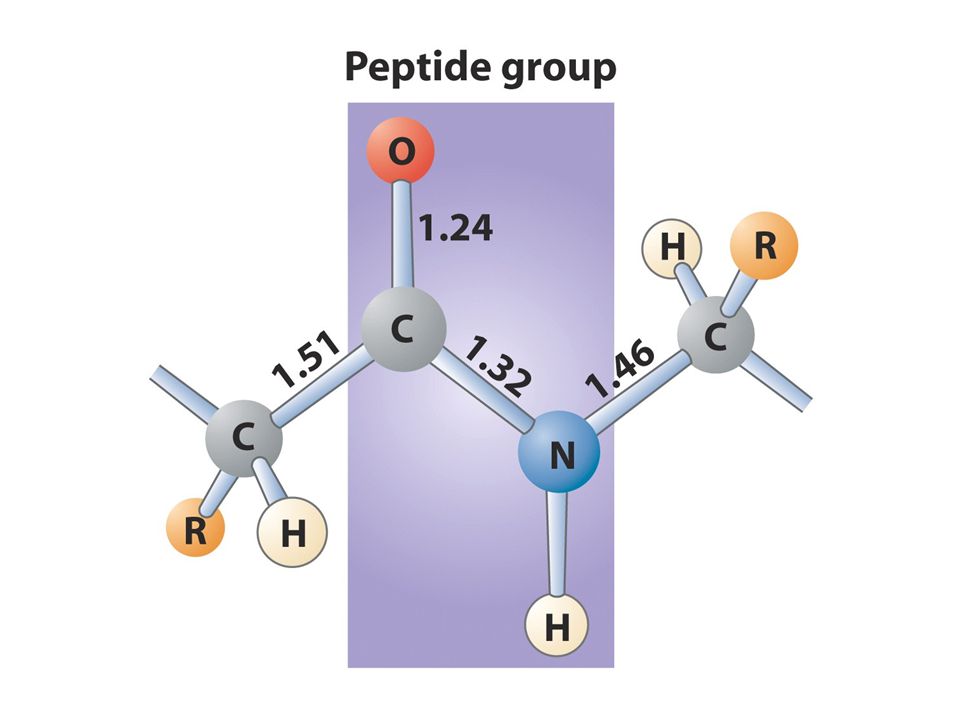
Protein yapısı Primer transkript tamamen olgun mRNAya dönüştüğünde protein sentezi başlar Proteinler amino asit polimerleridirler yani polipeptidlerdir. Tüm aminoasitler genel bir yapıya sahiptir ve 20 tanesi protein yapısında bulunurlar Peptid bağı kovalent bir bağdır ve 1. aminoasitin COOH ucu ile 2. aminoasitin amino ucu arasından bir molekül su çıkmasıyla elde edilir

10
Proteinler 4 düzeyde organize olurlar
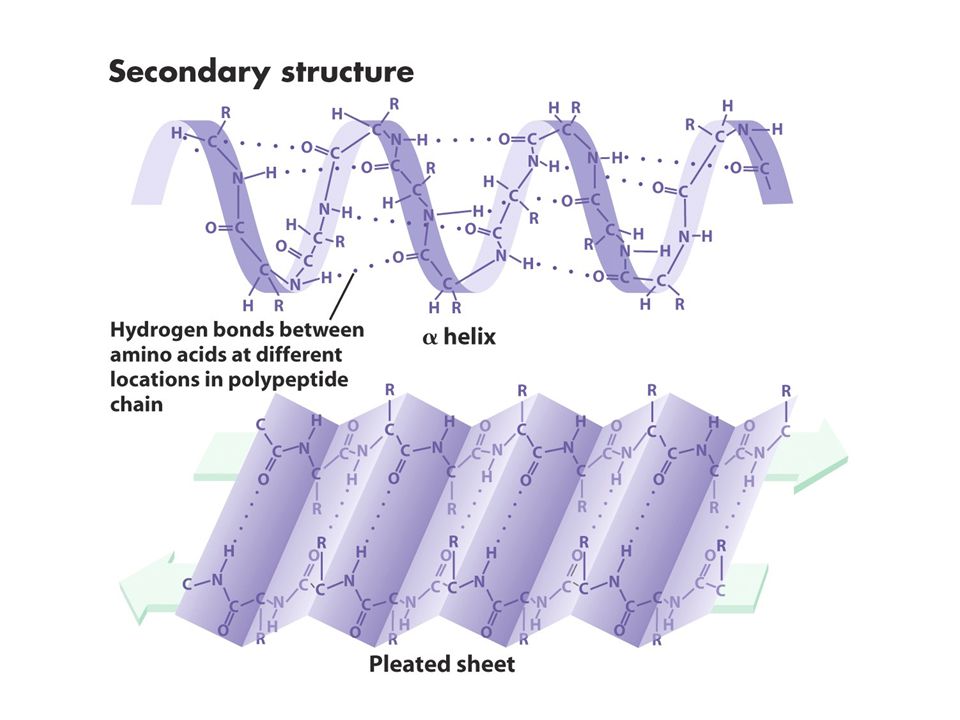
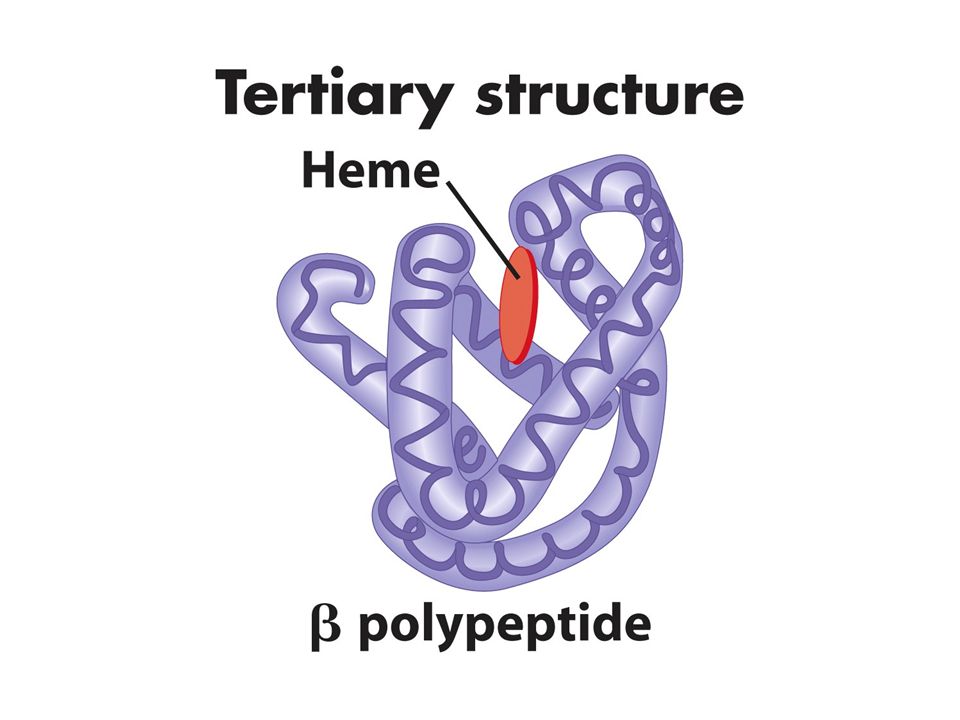
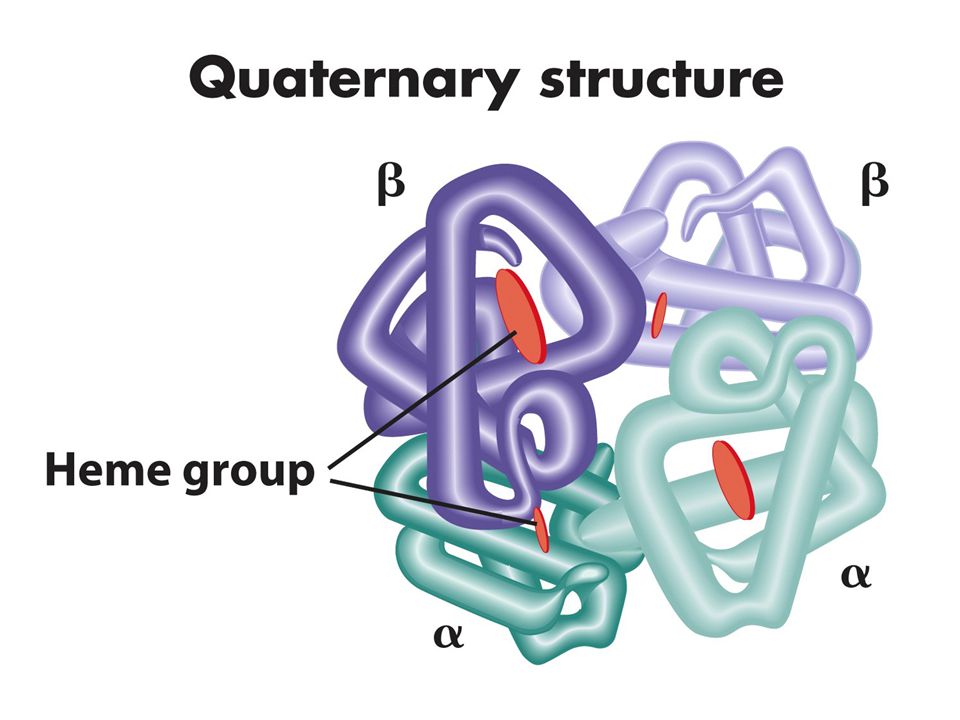
Primer yapı düz lineer biçimde aa dizilimidir Sekonder yapı polipeptid zincirinin katlanmasıyla oluşan özel bir yapıdır bu yapı lineer yapıdaki yakın aa arasında bağ güçlerinden oluşur: zayıf bağlar (H bağları, elektrostatik güçler ve van der waals bağları). En genel tipleri ise alfa helix beta pleated sheet. Aynı protein bu yapılardan birini yada herikisini içerebilir. Tersiyer yapı sekonder yapının katlanmasıyla oluşur Quaterner yapı ise bir yada birkaç polipeptidin katlanarak altbirimleriyle oluşturduğu kompleks yapıdır.Bu yapı farklı polipeptidlerden oluşursa heterodimer aynı tip polipeptidler arasında olursa homodimer oluşturur. Bazı proteinler kompakt yapıdadır enzim ve antikorlar globüler yapıdadır lineer yapıdaki proteinlere ise fibröz proteinler denir
. En genel tipleri ise alfa helix beta pleated sheet. Aynı protein bu yapılardan birini yada herikisini içerebilir. Tersiyer yapı sekonder yapının katlanmasıyla oluşur. Quaterner yapı ise bir yada birkaç polipeptidin katlanarak altbirimleriyle oluşturduğu kompleks yapıdır.Bu yapı farklı polipeptidlerden oluşursa heterodimer aynı tip polipeptidler arasında olursa homodimer oluşturur. Bazı proteinler kompakt yapıdadır enzim ve antikorlar globüler yapıdadır lineer yapıdaki proteinlere ise fibröz proteinler denir.")
16
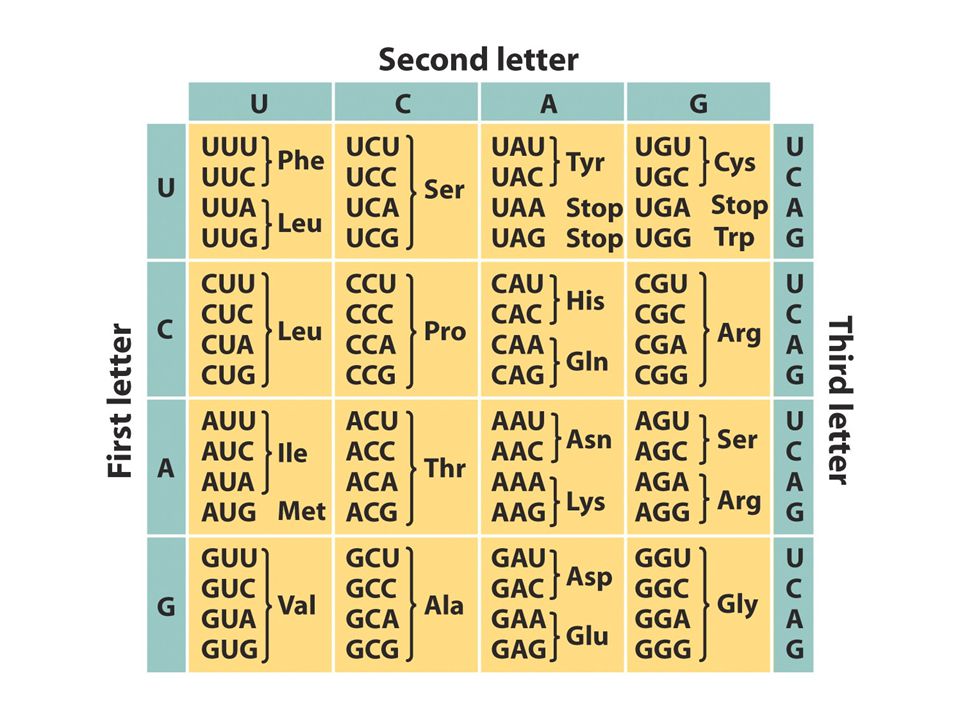
Genetik kod Eğer genler DNA segmenti ise ve DNA zinciri nt dizilimi ise nt dizisinin proteinlerin aa dizisini dikte etmesi gerekir.DNA dizisi protein dizisini nasıl belirler? Eğer nt ler bir koddaki harfler ise harflerin kombinasyonu farklı aminoasitleri temsil eden kelimeleri oluşturur. Kod nasıl okunur?Üst üste çakışırmı çakışmazmı? mRNA da bulunan Kaç tane harf kelimeyi yada kodonu oluşturur ve hangi kodon hangi aa’ i şifreler.

17
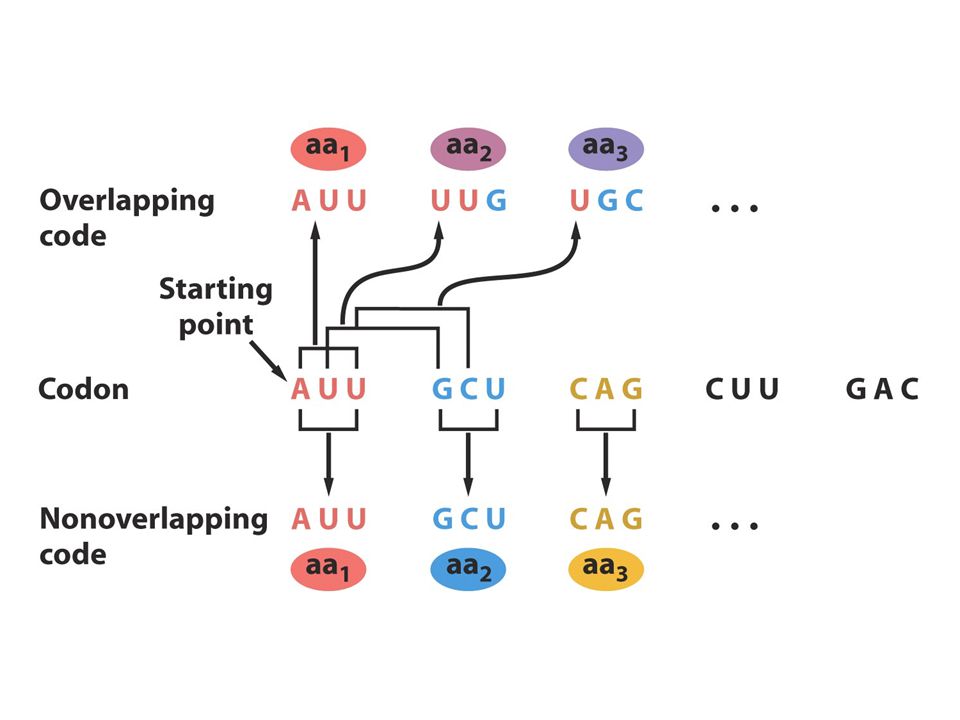
Çakışan-çakışmayan kod
Şekil 9.6 çakışan ve çakışmayan kodonlar arasındaki farkı göstermektedir. Örnek 3 harfli yada triplet kodu göstermektedir.çakışmayan kodda ardıl aa ler ardıl kodonlarca temsil edilir Çakışan kodda ise ardıl aa’ler bazı ardıl bazlarca temsil edilir örneğin bir kodonun son iki bazı sonraki kodonun ilk iki bazı olabilir. 1961 yılında genetik kodun çakışmayan olduğu bulunmuştur.

19
Kodondaki harf sayıları
Eğer mRNA baştan sona okunursa 4 A,G,C,U)bazdan biri herbir pozisyonda bir kez bulunur. Eğer kelime bir harf uzunluğundaysa sadece 4 kelime olasıdır.Sözlük genetik kod olamaz çünkü protein yapısındaki 20 aa için bir kelime olması gerekir.Eğer kelime 2 harf uzunluğunda olsaydı 4x4=16 kelime olası olurdu (AU, CU, yada CC). Bu sözlük yeterince uzun değildir. Eğer 3 harften oluşsaydı 4x4x4=64 kelime olası olurdu.Bu sayı tüm aa leri kapsayacak kadar uzundur.
bazdan biri herbir pozisyonda bir kez bulunur. Eğer kelime bir harf uzunluğundaysa sadece 4 kelime olasıdır.Sözlük genetik kod olamaz çünkü protein yapısındaki 20 aa için bir kelime olması gerekir.Eğer kelime 2 harf uzunluğunda olsaydı 4x4=16 kelime olası olurdu (AU, CU, yada CC). Bu sözlük yeterince uzun değildir. Eğer 3 harften oluşsaydı 4x4x4=64 kelime olası olurdu.Bu sayı tüm aa leri kapsayacak kadar uzundur.")
20
Genetik kodda dejenerasyon
Her pozisyon için 4 harf varsa 3 harfli kodon 64 kelime oluşturur. Sadece 20 kelime 20 aminoasit için gerekliyse diğer kodonlar ne işe yaramaktadır? Crick’in çalışmaları genetik kodun dejenere olduğunu ortaya koymuştur buna göre bazı aa ler birden fazla kodonca temsil edilmektedir. Herhangi bir aa kodlamayan kodonlar stop kodonlardır UAG-amber kodon UGA-opal kodon UAA ochre kodon

22
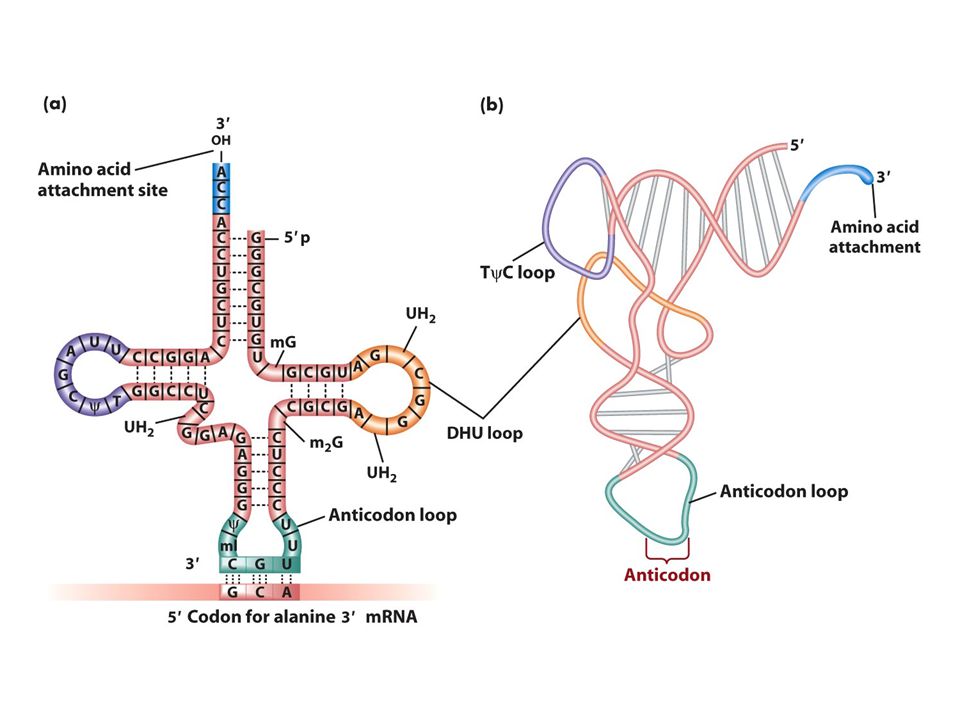
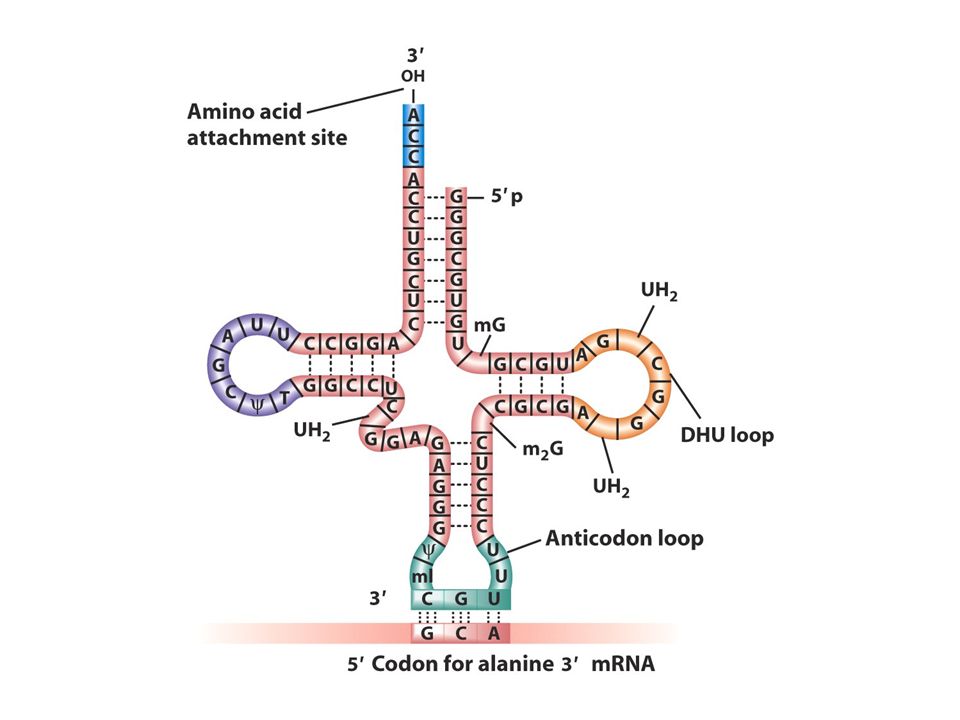
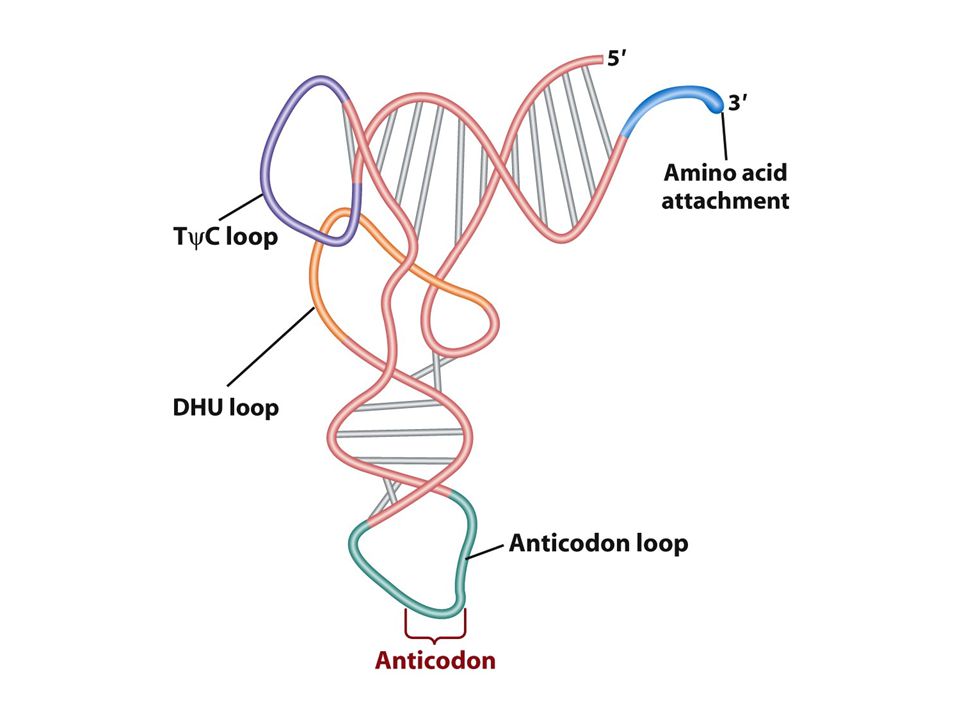
tRNA: adaptör Herbir aa için bir tane tRNA molekülü vardır herbir aa spesifik bir tRNA molekülüne bağlanır mRNA kodonları ile tRNA antikodonu RNA-RNA baz çifti oluşturur.Kodon antikodona komplementerdir. Aa ler tRNAlara amino açil tRNA sentetaz denen enzimlerce takılır ve tRNA böylece aa ile yüklenir. Her aa tRNA’nın 3’ CCA ucuna takılır.

26
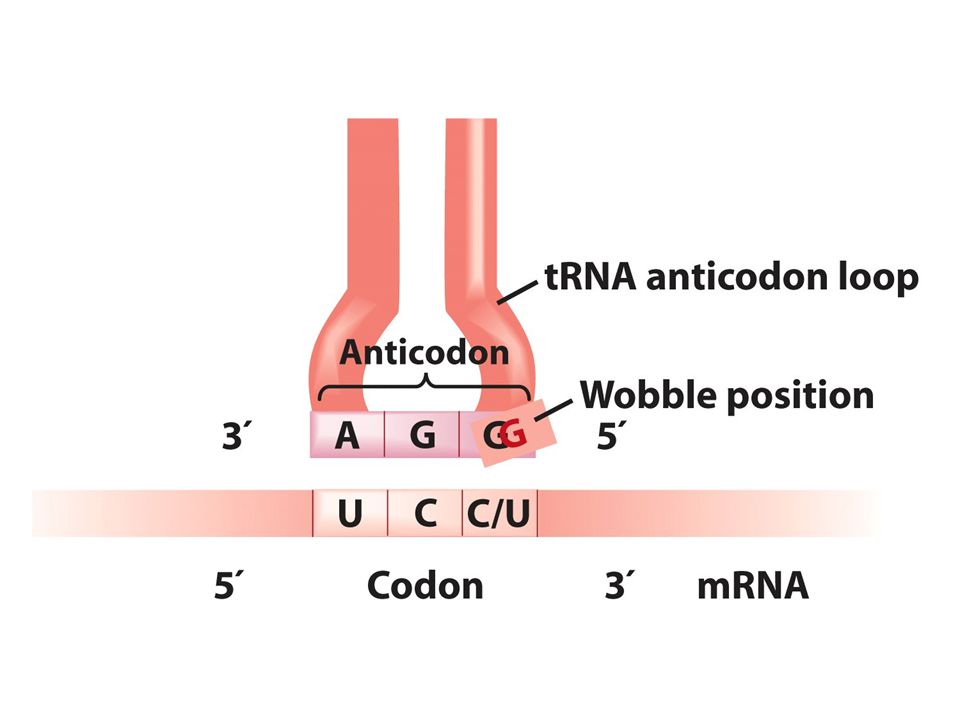
Wobble pozisyonu Bir aa için kodon sayısı değişir.1-6arasında kodon bazan bir aa için gereklidir örneğin serin için 6 tane kodon vardır. Neden genetik kod bu varyasyonu gösterir 1. çoğu aa ler ribozomlara alternatif tRNAlar ile taşınır herbir tip farklı bir antikodonla baz çifti oluşturur. 2.bazı yüklü tRNAlar spesifik aa lerini birkaç kodondan birine getirir ve bu tRNAlar sadece kendisine komplementer olan kodona değil birkaç alternatif kodonu tanır. Gevşek bir bağlantıyla kodon antikodon eşleşmesi gerçekleşir bu eşleşme kodonun 3’ ucunda antikodonun ise 5’ ucunda olur buna wobble baz eşleşmesi denir. Wobble pozisyonunda antikodonun 3. nükleotidi iki biçimde eşleşir biri komplementer olduğu bazla diğeri komplementer olmayan bazla İnosin tRNA yapısında bulunan ender bazlardan birirdir.

30
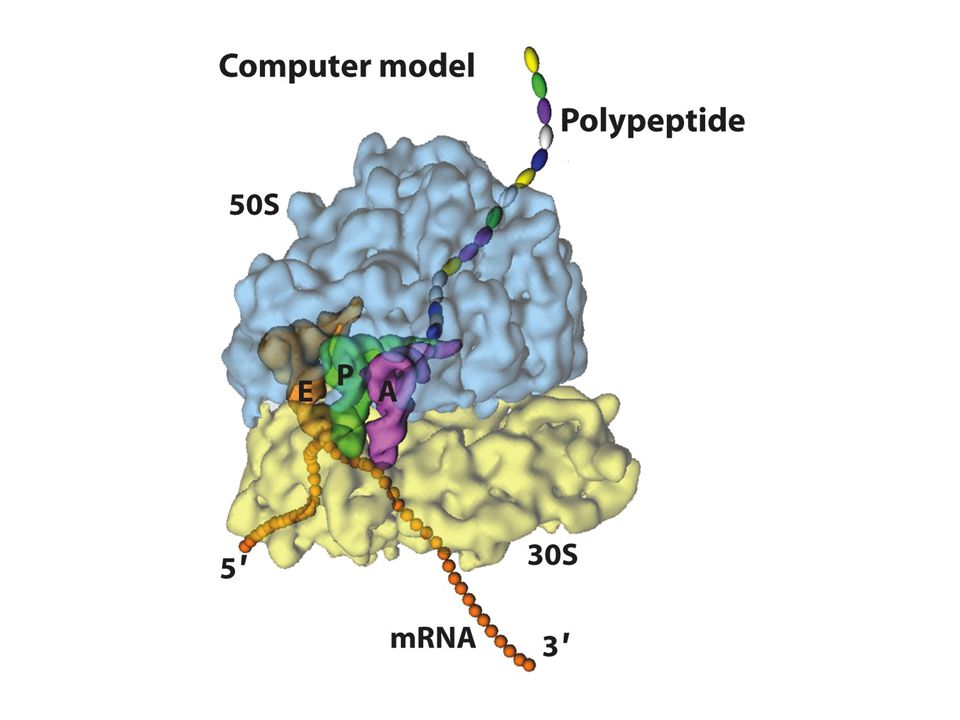
ribozomlar Protein sentezinin gerçekleştiği organellerdir
mRNAnın bağlandığı yer küçük altbirimdir ve tRNA için üç tane bağlanma bölgesi vardır A: aminoaçil tRNA bağlanır P: peptidil tRNA bağlanır E:çıkış kanalı (deaçile(yüksüz- tRNA bağlanır) Deşifre (decoding) merkezi:30S altbiriminde bulunur ve sadece kodonla komplementer tRNA A bölgesine kabul edilmesini sağlar Peptidil transferaz merkezi:50S biriminde bulunur ve peptid bağının oluşumunu katalizler
 Deşifre (decoding) merkezi:30S altbiriminde bulunur ve sadece kodonla komplementer tRNA A bölgesine kabul edilmesini sağlar. Peptidil transferaz merkezi:50S biriminde bulunur ve peptid bağının oluşumunu katalizler.")
34
translasyon Üç evreden oluşur Başlangıç:initiation Uzama :elongation
Terminasyon:sonlanma

35
Amino asitlerin aktivasyonu
Amino-açil tRNA sentetaz enzimi iki aşamada amino asitleri tRNA molekülüne bağlar. Enerji ATP hidrolizinden sağlanır Önce aa karboksil grubu ile AMP’ye bağlanır ve adenile amino asit oluşur. Bu kompleks enzimden ayrılmadan AMP’ye bağlı karboksil ucundan aa tRNA molekülünün 3’ ucunda bulunan CCA üçlü nükleotidinin Adeninine bağlı olan şekerinin OH grubuna transfer edilir. Bu bağ ester bağıdır ve sonuçta aminoaçil-tRNA oluşturulur.

36
Aa aktivasyonu Amino açil-tRNA sentetaz enzimleri aa birimlerini tRNA üzerine aktarırlar

37
Aa aktivasyonu

38
Aa aktivasyonu

39
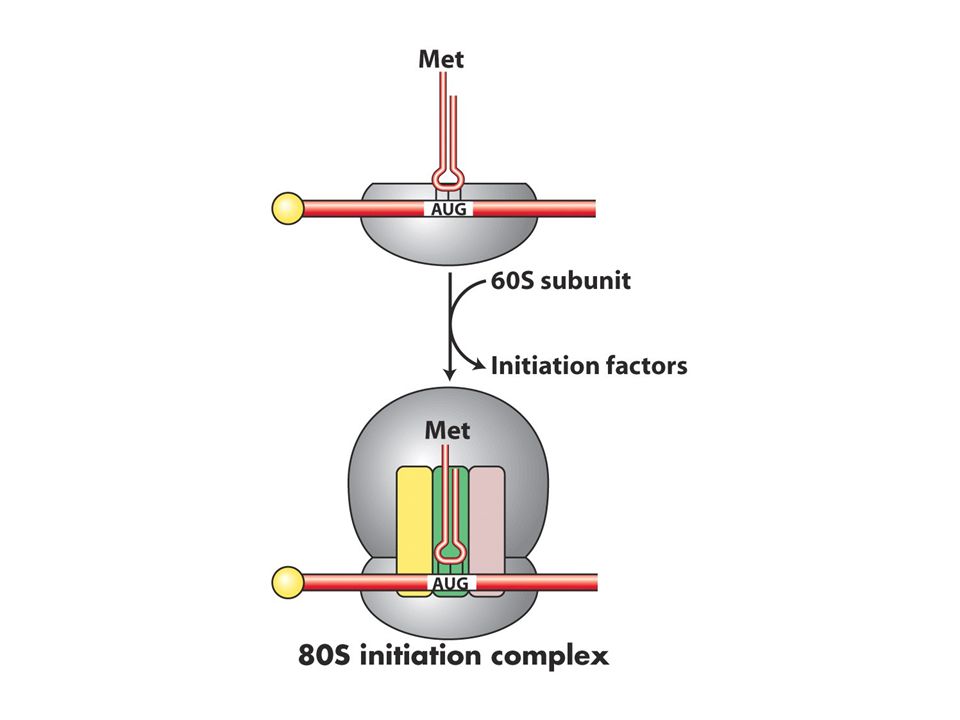
Translasyon başlangıcı
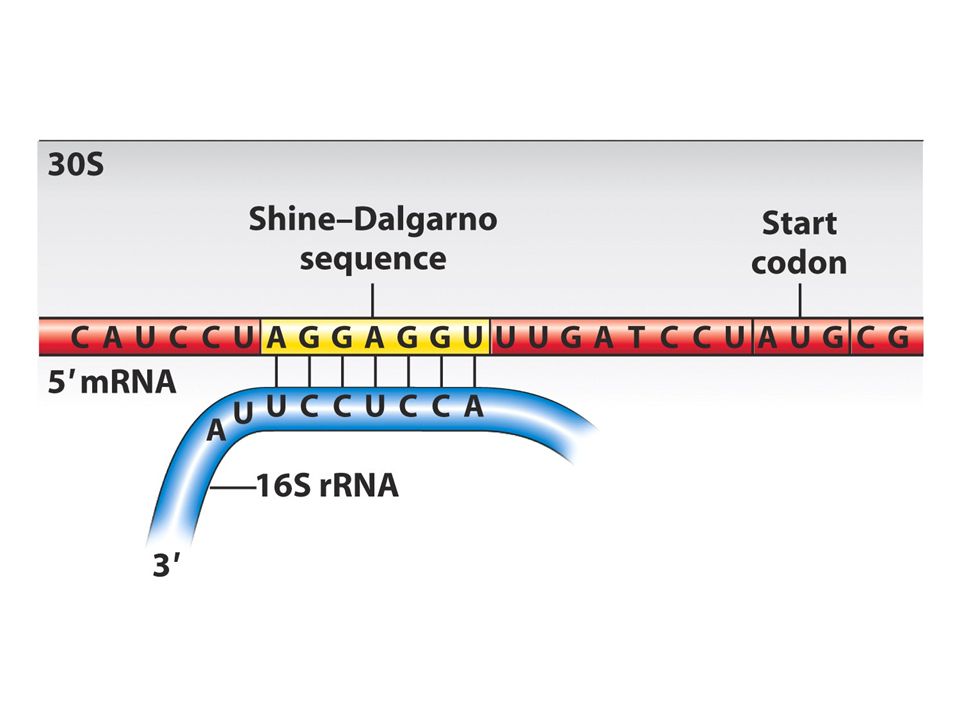
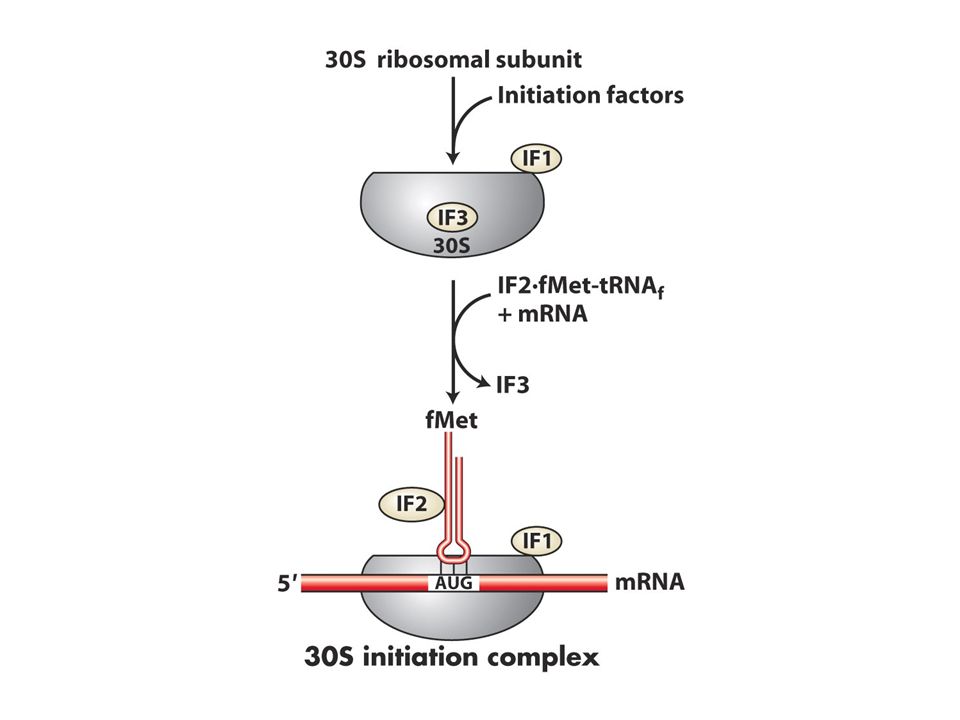
En önemli aşama ilk amino açil tRNA’nın A bölgesine yerleşmesidir dolayısıyla doğru okuma çerçevesi sağlanır. Hem pro hem ökaryotlardaki ilk aa metiyonindir ve özel bir tRNA tarafından mettRNA olarak adlandırılır ve AUG kodonunu tanır.Bu tRNA initiator tRNA olarak da bilinir.Bakterilerde bu ilk metiyonin formillenir ve bu grup daha sonra çıkarılır. Bilindiği gibi pro ve ökaryotlarda 5’UTR bölgeleri vardır bu bölge transkripsyon başlangıcı ve translasyon başlangıç noktaları arasındadır. Prokaryotlarda başlangıç kodonundan önce özel bir dizi vardır buraya Shine dalgarno dizisi adı verilir bu dizi 30S ribozomunda bulunan 16S rRNA ile baz çifti oluşturur bu eşleşme ile P bölgesini öylesine bir pozisyona sokarki başlangıç tRNAsı buraya girer.

41
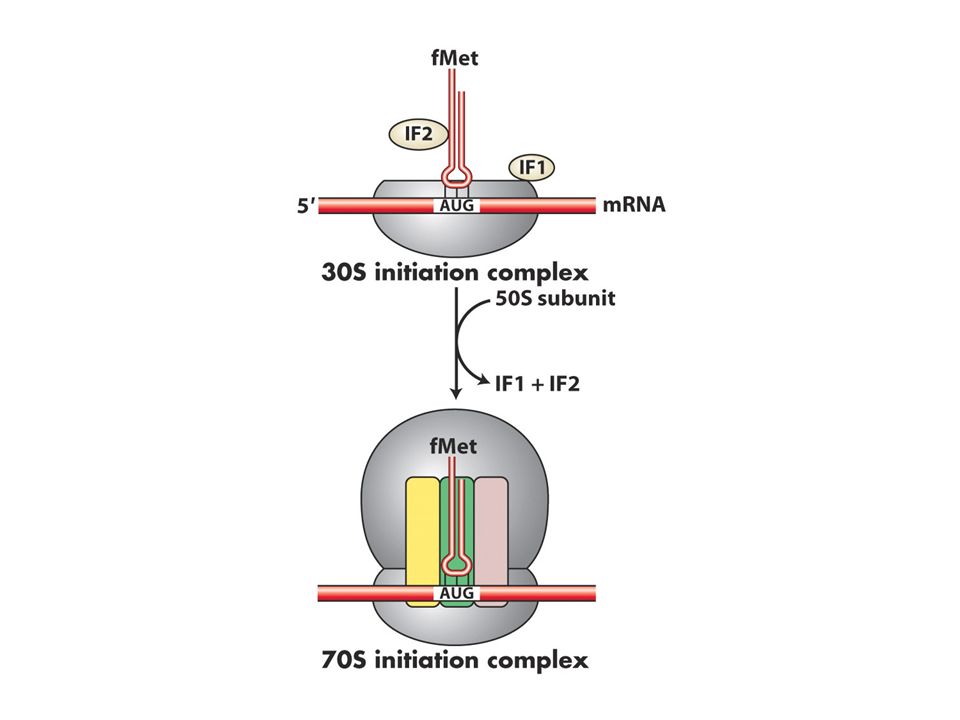
Bakterilerde üç tane başlangıç proteini (IF1,2 ve 3 doğru başlama için gereklidir.
IF3 50 ve 30S altbirimlerini ayrı tutar 1 ve 2 ise sadece başlangıç tRNAsının P bölgesine girmesini sağlar.30S altbirimi mRNA ve initiator tRNA başlangıç kompleksini oluşturur. 50S buraya katılınca başlangıç proteinleri ayrılır Prokaryotlarda çekirdek olmadığı için mRNA sentezlenirken translasyon shine dalgarno bölgesinden başlar.

44
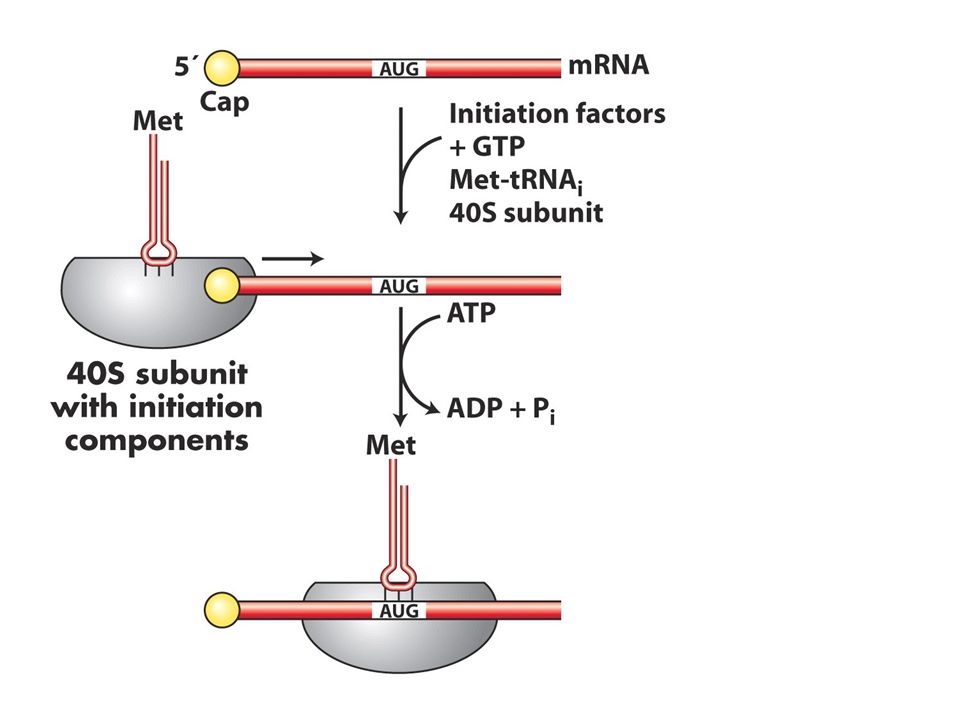
ökaryotlarda bu iki süreç farklı kompartmanlarda olduğu için mRNA olgunlaştıktan sonra sitoplazmaya geçer ve üzerinde birçok protein taşır ayrıca ikincil yapılarda oluşturur ve bu yapıların arasından AUG kodonunu açığa çıkarmak gerekir. Bunu gerçekleştiren proteinler eIF4A, B ve G dir.Bu proteinler cap, 40S altbirimi ve initiator tRNA ile etkileşir ve 5’-3’ yönünde kaymaya başlar ve baz çifti oluşturan yerleri açar. AUG bulunur ve sonra 60S birimi bağlanır ve başlama kompleksi oluşur

47
elongation Bu aşamada ribozom bir fabrika gibi çalışır.herbir aa büyüyen polipeptid zincirine eklenirken deaçile tRNA başka bir aa taşımak üzere geri dönüşüm döngüsüne girer. Uzama sırasında elongasyon faktör Tu ve EF-G kullanılır.aminoaçil tRNA EF-Tu ile bağlanır ve ternary kompleks oluşur. Elongasyon döngüsü başlangıç tRNA’sı P bölgesinde ve aminoaçil tRNA A bölgesine girince başlar. Buraya tRNA girince ribozom biçim değiştirir ve EF-Tu ternary kompleksten ayrılır ve iki aa büyük altbirimde bulunan peptidil transferaz merkezinde birbirine yakınlaşır. Burada peptid bağı oluşur ve dipeptid A bölgesine kayar. Devreye EF-G girerek A bölgesindeki tRNA P bölgesine P bölgesindeki boş tRNA E bölgesine kayar ve mRNA ribozom içinde hareket ederek bir sonraki kodon A bölgesine karşılık gelir ve EF-G A bölgesinden çıkarak buraya yeni bir aa yüklü tRNA gelmesi için burayı boşaltır. Sonraki aşamada ise A bölgesine yeni tRNA girince E bölgesindeki boş tRNA sitoplazmaya döner ve bu döngü durdurucu kodona kadar sürer.

49
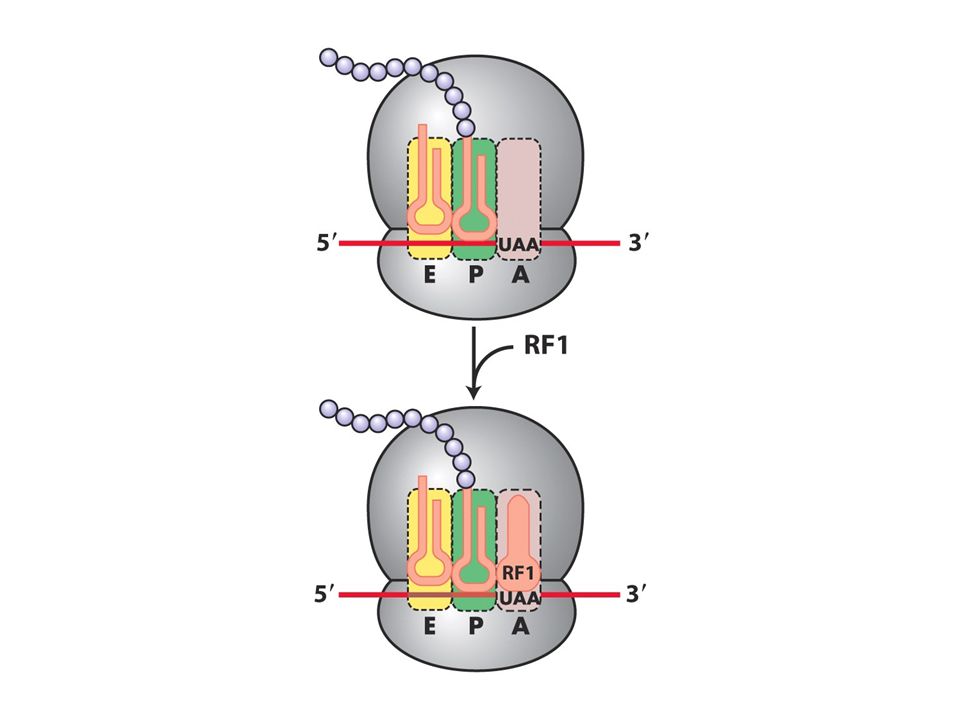
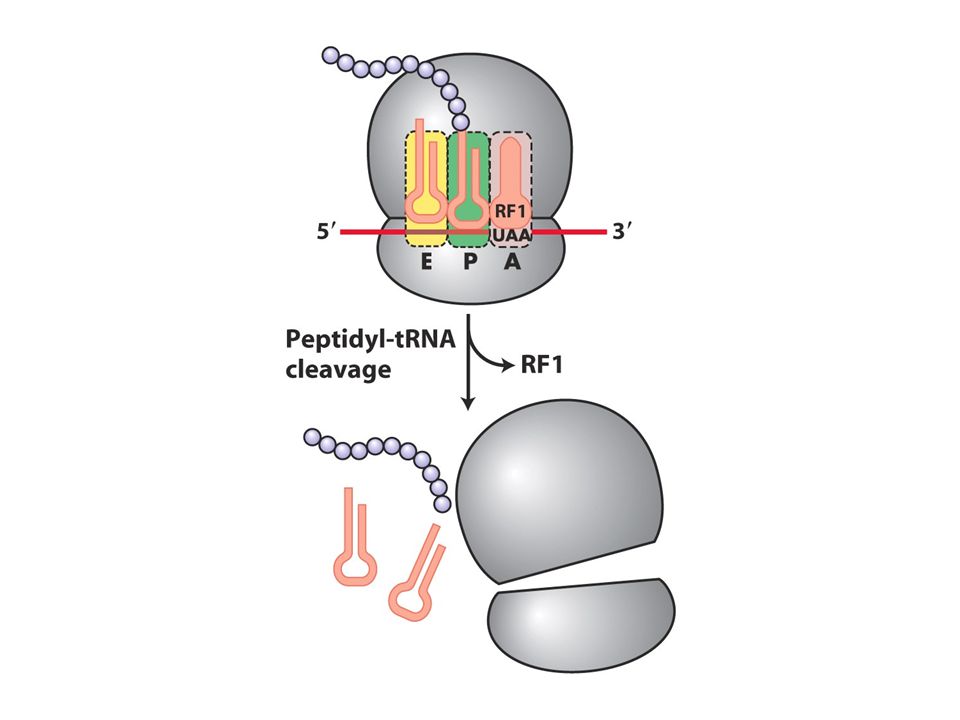
Translasyon terminasyonu
A bölgesi üç tane stop kodona gelinceye kadar elongasyon sürer. Buraya release faktörler (RF1, RF2 ve 3) bağlanır.Bakterilerde RF1 UAA yada UAGyı tanır RF2 UAA yada UGAyı tanır. RF3 herikisine yardımcı olur.RF1 ve 2 tarafında A bölgesinin tanınması ternary komplekten farklıdır. Stop kodonlar RF proteindeki tripeptid tarafından tanınır antikodondan değil.Ayrıca RFler 30Sdeki A bölgesine uyarlar ve peptid bağı oluşumunda görev almazlar.Bunun yerine su molekülü peptidiltransferaz merkezine yerleşir ve P bölgesindeki tRNAda bulunan polipeptid zinciri salınır.
 bağlanır.Bakterilerde RF1 UAA yada UAGyı tanır RF2 UAA yada UGAyı tanır. RF3 herikisine yardımcı olur.RF1 ve 2 tarafında A bölgesinin tanınması ternary komplekten farklıdır. Stop kodonlar RF proteindeki tripeptid tarafından tanınır antikodondan değil.Ayrıca RFler 30Sdeki A bölgesine uyarlar ve peptid bağı oluşumunda görev almazlar.Bunun yerine su molekülü peptidiltransferaz merkezine yerleşir ve P bölgesindeki tRNAda bulunan polipeptid zinciri salınır.")
52
Moleküler benzerlik (mimicry)
EF-G şekli ternary komplekse benzer (aa tRNA+ EF-Tu) RF şekli ise deaçile tRNAya benzer. Bir molekülü diğerine benzerliğinden dolayı yapısını tahmin etmeye moleküler mimikri denir.EF-Gnin ternary komplekse benzemesi onun 30S ve 50S altbirimleriyle etkileşime girmesini işaret eder ve bu interaksiyonda ternary kompleksin yerinin EF-G tarafından doldurulduğu anlamına gelir.
 RF şekli ise deaçile tRNAya benzer. Bir molekülü diğerine benzerliğinden dolayı yapısını tahmin etmeye moleküler mimikri denir.EF-Gnin ternary komplekse benzemesi onun 30S ve 50S altbirimleriyle etkileşime girmesini işaret eder ve bu interaksiyonda ternary kompleksin yerinin EF-G tarafından doldurulduğu anlamına gelir.")
53
Poliribozomlar Birçok proteinin sentezi 20sn ile birkaç dk arasında gerçekleşir. Buda poliribozomlar aracılığıyla gerçekleşir İlk ribozom belirli bir noktaya geldiğinde başka bir ribozom AUG kodonuna bağlanır ve sentezi sürdürür. 2. ribozoma yeterince yer açılınca. Ökaryot mRNA’larının 5’ ve 3’ uçları birbiriyle bağlantılıdır. Dolayısıyla mRNA’nın 3’ ucunda işini bitirip ayrılan ribozom altbirimleri hemen yeniden kompleks oluşturarak 5’ ucundan senteze başlar

Benzer bir sunumlar






>")





 ZM106 Biyokimya 11. Hafta.>")


>")




>")



 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")