Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Ondokuz Mayıs Üniversitesi Tıp Fakültesi Tıbbi Onkoloji
HÜCRE SİKLUSU Ondokuz Mayıs Üniversitesi Tıp Fakültesi Tıbbi Onkoloji Dr. Müge Karaoğlanoğlu 06/2008

2
HÜCRE SİKLUSUNUN EVRELERİ
M evresi: mitoz G1 evresi: mitoz ile DNA replikasyonu arasındaki süreç S evresi: DNA sentezi G2 evresi: mitoza hazırlık dönemi

3
MİTOZ EVRESİ Mitoz evresinde kromozomlar birbirinden ayrılır ve hücre bölünmesi ( sitokinez) gerçekleşir. Bu evre 1 saat sürer

4
Profaz: kromatin fibrillerinin daha fazla kısalıp kalınlaşması sağlanır. Çekirdekçik ortadan kalkmaya başlar,mikrotubul polimerizasyonuyla mitotik iğcikler oluşur. Prometafaz: Çekirdek zarı parçalanır. Kinetokor mikrotubulleri herbir kromozoma sentromerinin her iki tarafındaki kinetokorlardan tutunur. Metafaz: Sentriol çiftleri hücrenin zıt kutuplarına ulaşır. Kromozomlar mitotik iğcikler tarafından metafaz düzlemine doğru çekilirler. Anafaz: Herbir kromozomun kardeş kromatitleri kinetokor mikrotubullerince zıt kutuplara doğru çekilir (Kinetokor mikrotubulleri kısalır). Böylece herbir kromozom takımı birbirinden ayrılıp, kutuplara doğru çekilir. Telofaz : Profazın tüm olayları tersine döner. Çekirdek zarı yeniden birleşir, çekirdek oluşur. DNA’lar kromatin şeklinde gevşeyerek yeniden şekillenir. Çekirdekçik ortaya çıkar. Sitoplazma bölünmesi : Mikroflamentlerin oluşturduğu kasılma halkasıyla, sitoplazma boğumlanır ve ikiye bölünür
. Böylece herbir kromozom takımı birbirinden ayrılıp, kutuplara doğru çekilir. Telofaz : Profazın tüm olayları tersine döner. Çekirdek zarı yeniden birleşir, çekirdek oluşur. DNA’lar kromatin şeklinde gevşeyerek yeniden şekillenir. Çekirdekçik ortaya çıkar. Sitoplazma bölünmesi : Mikroflamentlerin oluşturduğu kasılma halkasıyla, sitoplazma boğumlanır ve ikiye bölünür.")
5
Hücre döngüsünün %95’i interfazda (mitozlar arası dönem) geçer.
 geçer.")
6
‘G1’ EVRESİ G1 evresi: ilk bölünmede oluşan eş hücrelerin tekrar hücre bölünmesine girmeden önceki evresidir. Bu evrede DNA replikasyonu olmaz; RNA ve protein sentezi devam eder; evre 3-4 saat sürer ve hücre S evresine hazırlanır.

7
‘S’ EVRESİ S evresi: DNA replikasyonu, kromozomun çiftlenmesi, RNA ve protein sentezinin olduğu evredir. Bu evre 6-8 saat sürer.

8
DNA SENTEZİ DNA ve RNA’nın yapısını nükleotidler oluşturur.
Bu nükleotidlerin yapısında pürin ve pirimidin denilen azotlu bazlar bulunur. Pürin bazları: adenin, guanin Pirimidin bazları: sitozin, urasil, timin Pürin ve pirimidin inhibitörleri, nükleotid ve dolayısıyla DNA-RNA sentezini bozarak etki gösterirler.

9
DNA SENTEZİ DNA; çok sayıda monodeoksiribonükleotidin 3 5 fosfodiester bağlarıyla bağlanmasıyla oluşur. Bu fosfodiester bağları kimyasallar ile ayrılabilir veya nükleazlar tarafından enzimatik olarak hidrolize edilebilirler. Çift sarmalda, iki iplik arasında dar ve geniş girinti oluşur. Daktinomisin dar girintiye etki ederek DNA ve RNA sentezini bozar (iplikleri birbirine yapıştırır).
.")
10
DNA SENTEZİ DNA’nın replikasyonu: DNA moleküllerinin doğru kopyalarının yapılmasıdır Bir DNA molekülünün iki kolundan her biri, yeni bir DNA kolu sentezi için kalıp olarak görev görür. Sonuçta iki yeni DNA molekülü meydana gelir. DNA replikasyonu, kromatin üzerinde binlerce yerde birden başlar ve devam eder

11
DNA SENTEZİ DNA replikasyonu, orijin diye adlandırılan bir başlama noktasında başlar genellikle iki yöndeki replikasyon çatallarında 3 yönünde ilerler kalıp olarak görev gören kol 3 5 yönünde okunur

12
DNA helikaz: tek iplikli DNA’ya bağlanır, yakındaki çift iplikli bölgeye ilerleyerek ipliklerin açılmasını sağlar. Bu iplikler açılırken süper kıvrımlar olabilir. DNA topoizomerazlar : bu kıvrımları ortadan kaldırlar. Tip I DNA topoizomeraz: reversible olarak çift sarmalın tek ipliğini keser. Hem nükleaz (kesen) hem de ligaz (bağlayan) aktivitesi vardır. Tip II DNA topoizomeraz: her iki iplikte de kırılmalar yapar. Bunda da nükleaz ve ligaz aktivitesi vardır.
 hem de ligaz (bağlayan) aktivitesi vardır. Tip II DNA topoizomeraz: her iki iplikte de kırılmalar yapar. Bunda da nükleaz ve ligaz aktivitesi vardır.")
13
DNA SENTEZİ DNA polimeraz III: DNA zincir uzamasını sağlar (5 3 yönünde). ilk nükleotid alıcısı olarak RNA primerini kullanır. Ek olarak 35 kontrol okuması da yapar. Yanlış okunan kısmı çıkarır ( ekzonükleaz aktivitesi). DNA polimeraz III, bir RNA primerine yaklaşana kadar DNA sentezler. Bu durumda RNA çıkarılır ve boşluk DNA polimeraz I ile doldurulur ( polimeraz ve endonükleaz aktiviteleri).
. ilk nükleotid alıcısı olarak RNA primerini kullanır. Ek olarak 35 kontrol okuması da yapar. Yanlış okunan kısmı çıkarır ( ekzonükleaz aktivitesi). DNA polimeraz III, bir RNA primerine yaklaşana kadar DNA sentezler. Bu durumda RNA çıkarılır ve boşluk DNA polimeraz I ile doldurulur ( polimeraz ve endonükleaz aktiviteleri).")
14
DNA SENTEZİ DNA replikasyonunda DNA polimerazların etkisi, kalıp kolun karşısında büyüyen DNA koluna uygun deoksinükleozid trifosfatlardan (dNTP) deoksinükleozid monofosfatların (dNMP) girişini sağlamaktır. Yeni bir DNA şeridinin sentezi tamamlandıktan sonra DNA giraz, replike olmuş DNA’nın tekrar doğal haline kıvrılmasına yardımcı olur .
 deoksinükleozid monofosfatların (dNMP) girişini sağlamaktır. Yeni bir DNA şeridinin sentezi tamamlandıktan sonra DNA giraz, replike olmuş DNA’nın tekrar doğal haline kıvrılmasına yardımcı olur .")
15
RNA SENTEZİ RNA sentezi (transkripsiyon), DNA’da saklanan genetik bilgilerin bir RNA molekülü (mRNA, tRNA, rRNA) şeklinde kopyalanması veya yazılması olayıdır. Bir RNA molekülü, DNA’nın kalıp kolunun dizilişini bütünleyici ribonükleotidlerin ATP, GTP, CTP ve UTP’tan pirofosfatlar ayrılması suretiyle polimerizasyonu sonucunda, 5 3 yönünde sentezlenir.
, DNA’da saklanan genetik bilgilerin bir RNA molekülü (mRNA, tRNA, rRNA) şeklinde kopyalanması veya yazılması olayıdır. Bir RNA molekülü, DNA’nın kalıp kolunun dizilişini bütünleyici ribonükleotidlerin ATP, GTP, CTP ve UTP’tan pirofosfatlar ayrılması suretiyle polimerizasyonu sonucunda, 5 3 yönünde sentezlenir.")
16
RNA SENTEZİ RNA sentezi için RNA polimeraza, başlama ve sonlanma sinyallerine gereksinim vardır

17
RNA SENTEZİ RNA polimeraz kalıp kol tarafından yönetilen ve baz eşleşmesi kuralları tarafından yorumlanan spesifik bir diziliş içinde ribonükleotidleri polimerize ederken pirofosfatlar serbest bırakılır ve böylece RNA sentezlenir

18
RNA SENTEZİ Transkripsiyon sonunda oluşan RNA’lar primer RNA’lar diye adlandırılırlar ve genellikle hemen kullanılmazlar. RNA processing diye tanımlanan bazı işlemlerden geçtikten sonra işlev görebilecek olgun RNA’lar haline gelirler.

19
PROTEİN SENTEZİ Protein sentezi (translasyon), gen ifadesinin son aşamasıdır. Transkripsiyonla RNA’ya kopyalanan genetik bilgi son olarak bir protein veya polipeptit zinciri haline dönüştürülür .
, gen ifadesinin son aşamasıdır. Transkripsiyonla RNA’ya kopyalanan genetik bilgi son olarak bir protein veya polipeptit zinciri haline dönüştürülür .")
20
PROTEİN SENTEZİ Protein sentezinin üç komponenti: mRNA, tRNA ve ribozomlardır. mRNA; proteinin amino asit sırasını belirleyen kodu içerir. Protein sentezi başlayacağı zaman, sitoplazmada bulunan amino asitler, kendilerine özgü, Mg2+ gerektiren aminoaçil-tRNA sentetaz enzimleri yardımıyla tRNA’lara bağlanarak aminoaçil-tRNA şeklinde aktiflenirler.

21
Protein sentezi başlarken; ribozom alt üniteleri, mRNA , aminoaçil -tRNA, GTP ve başlama faktörleri (IF)’nin varlığı gereklidir. mRNA’nın 5′ ucuna yakın bir bölgesinde başlama kompleksi oluşur . mRNA üzerinde başlama kodonu ( AUG veya GUG ) özel bir başlatıcı tRNA ile tanınır. Bu tRNA prokaryotlarda N-formile metiyonin içerir.
 özel bir başlatıcı tRNA ile tanınır. Bu tRNA prokaryotlarda N-formile metiyonin içerir.")
22
Başlama kompleksi oluştuktan sonra, GTP’ın hidrolizi ve elongasyon faktörü (EF-Tu) sayesinde, bu kompleksteki A (amino) yerine, mRNA’nın buraya rast gelen kodonunu tamamlayan antikodonu içeren aminoaçil-tRNA gelir.
 sayesinde, bu kompleksteki A (amino) yerine, mRNA’nın buraya rast gelen kodonunu tamamlayan antikodonu içeren aminoaçil-tRNA gelir.")
23
Ribozomda bulunan peptidil transferaz enziminin katalitik etkisiyle P (peptid) yerindeki fmet-tRNA’da bulunan aminoaçil grubu, A (amino) yerindeki aminoaçil-tRNA’nın aminoaçilinin serbest amino grubuna peptit bağı ile bağlanmak üzere taşınır.
 yerindeki fmet-tRNA’da bulunan aminoaçil grubu, A (amino) yerindeki aminoaçil-tRNA’nın aminoaçilinin serbest amino grubuna peptit bağı ile bağlanmak üzere taşınır.")
24
GTP’nin hidrolizi ve EF-G (uzama faktörü) sayesinde P yerindeki tRNA kompleksten ayrılır
A yerindeki dipeptidil-tRNA, A yerinden P yerine yer değiştirirken ribozom, mRNA üzerinde 3 ucuna doğru bir kodon ilerler ve A yerine uygun aminoaçil-tRNA gelir.

25
PROTEİN SENTEZİ Son iki basamaktaki olayların tekrarı sonucunda polipeptit zinciri amino-terminal uçtan karboksil-terminal uca doğru uzar. Polipeptit zincirinin uzaması sonlandırılacağı zaman, A yerine UAG, UAA, UGA sonlandırma kodonlarından biri gelir; buraya terminasyon faktörü (RF) bağlanır ve önce polipeptidil-tRNA bağı hidroliz olur daha sonra diğer komponentler dissosiye olurlar.
 bağlanır ve önce polipeptidil-tRNA bağı hidroliz olur daha sonra diğer komponentler dissosiye olurlar.")
26
‘G2’ EVRESİ G2 evresi: S ve M evreleri arasındaki evredir.
Bu evrede DNA replikasyonu olmaz, RNA ve protein sentezi devam eder. evre 3-4 saat sürer ve hücre büyüklüğü iki katına ulaşır.

27
‘G0’ EVRESİ G0 evresi : Hücre döngüsünün dışında, son farklılaşmasını tamamlamış ve bölünmesi duran hücrelerin dinlenme evresidir. Birkaç saat, birkaç gün veya ömür boyu sürebilir. Bu evrede hücrede tüm biyokimyasal olaylar aktif bir şekilde devam etmektedir.

28
‘G0’ EVRESİ G0 dinlenme evresindeki hücre bölüneceği zaman, G1 evresinin başlangıcından itibaren hücre bölünme döngüsüne girer. Büyüme faktörleri, sitokinler ve tümör virüsleri gibi mitojenik iletiler, G0 evresindeki hücrenin G1 evresine girmesine yol açmaktadır.

29
NORMAL HÜCRE SİKLUSU Büyüme faktörü Büyüme faktörü reseptörü Sinyal iletimi Kopyalanmanın aktivasyonu Hücre bölünmesi

30
SİKLİNLER ve KONTROL NOKTALARI ile regüle edilirler.
Hücre siklusunda önemli 3 basamak 1) Hücre yüzey reseptörleri 2) sinyal iletim yolu 3) Nükleer transkripsiyon faktörleridir. Bu olaylar; SİKLİNLER ve KONTROL NOKTALARI ile regüle edilirler.
 Hücre yüzey reseptörleri. 2) sinyal iletim yolu. 3) Nükleer transkripsiyon faktörleridir. Bu olaylar; SİKLİNLER ve KONTROL. NOKTALARI ile regüle. edilirler.")
31
Uyarı iletim yolu ile alınan sinyaller nükleusa iletilir.
Nükleusta transkripsiyon gen düzeyinde yapılır ve transkripsiyon faktörleri ile kontrol edilir.

32
Hücre proliferasyonuna yol açan transkripsiyon
faktörleri; Protoonkogenler; c-myc Tümör süpresör gen; p53, Rb Transkripsiyon faktörleri fosforile olur, DNA’ya afinitesi değişir.

33
HÜCRE SİKLUSU KONTROL NOKTALARI
G1/S ve G2/M Hücre siklusu kontrol mekanizmalarındaki defekt Kanser hücrelerindeki genetik bozukluğun ana nedeni

34
G1/S kontrol noktası Restriksiyon noktası: DNA hasarı kontrol edilir
DNA hasarı var ise; Hücre siklusta tutulur DNA tamir mekanizmaları harekete geçer DNA hasarı tamir edilemez ise; Apoptozis harekete geçer S fazı; Geri dönüşü olmayan nokta Hücre çoğalmadan önce son hazırlıkların yapıldığı dönem

35
G2/M kontrol noktası DNA kopyalanmasının tamamlanması
Hücrenin emniyetli bir şekilde mitoza başlaması Kardeş kromatidlerin ayrılması Özellikle iyonize radyasyonla hasarlanan hücreler G2/M kontrol noktasını aktive eder ve hücre G2’de kalır Bu noktadaki defektler kromozomal anomalilere neden olur

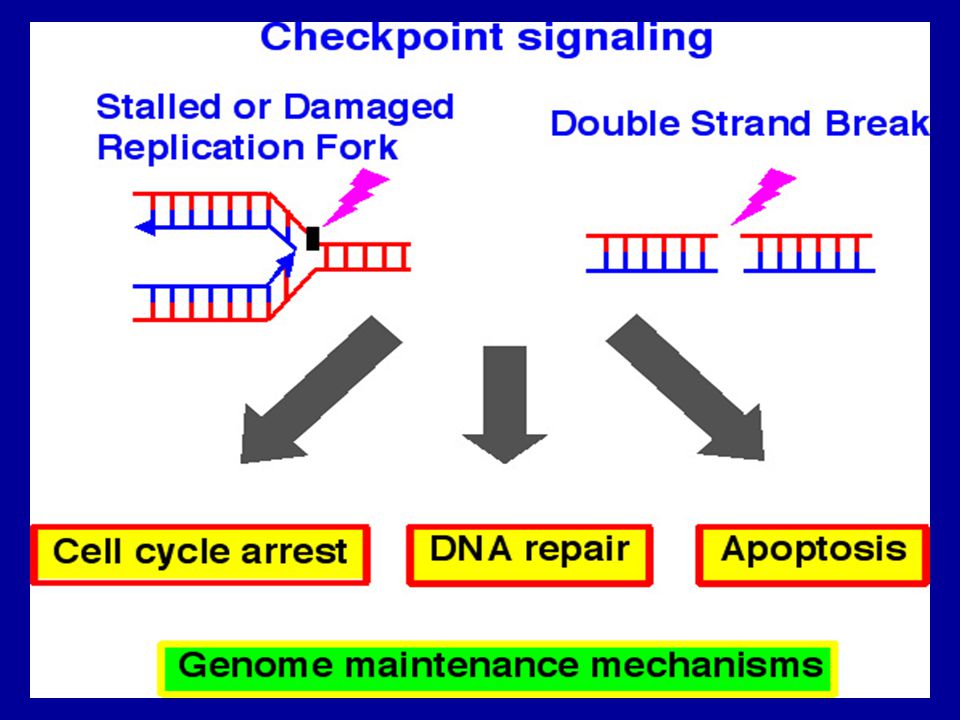
37
Hasar görmüş DNA’ya bağlanan protein komplekslerin hedefi ATM ve ATR olarak simgelenen birbiri ile ilintili iki tirozin kinazdır ve bunlar DNA hasarına cevaben aktifleşirler. ATM ve ATR, Chk2 ve Chk1 kinazları fosfatllayarak aktive ederler. Chk2 ve ATM tafından fosforillenen p53, G1/S kontrol noktasındaki duraksamadan sorumludur.

38
İĞ KONTROL NOKTASI Mitozun sonlarında yer alır.
Kromozomların yavru hücrelere tam ve doğru dağıtılması için mitotik iğ üzerindeki dizilimlerini gözlemler.

39
SİKLİNLER Hücre siklusunun spesifik fazlarında sentezlenirler.
Sırasıyla D, E, A, B ortaya çıkar. Siklin bağımlı kinazlar (CDK) ile kompleks oluşturunca etki gösterirler. CDK inhibitörleri ile veya proteazlarla inaktive edilirler.
 ile kompleks oluşturunca etki gösterirler. CDK inhibitörleri ile veya proteazlarla inaktive edilirler.")
40
SİKLİNLER Fonksiyonları CDK’ları aktive etmektir
CDK aktivasyonundan sonra siklin seviyeleri hızla düşer

41
SİKLİNLER CDK; hücre siklusu boyunca inaktif formda bulunur
Hücrenin diğer faza ilerlemesini, hedef proteinleri fosforize ederek sağlar Siklinlerle bağlandıktan sonra fosforilasyonla aktive olur

42
Sessiz hücreler büyüme faktörleri ile uyarıldığı zaman
Siklin D ve Siklin E konsantrasyon artımı SiklinD-CDK4 ve SiklinE-CDK2 aktivasyonu RB fosforilasyonu

43
hücre siklusunun moleküler açma-kapama merkezi
RB fosforilasyonu hücre siklusunun moleküler açma-kapama merkezi

44
E2F; transkripsiyon faktörlerindendir
E2F; transkripsiyon faktörlerindendir. Rb/E2F kompleksi, E2F tarafından kontrol edilen genlerin transkripsiyonunu engeller.

45
Kompleksten ayrılan hiperfosforilize RB, E2F kopyalanmasını aktive eder.

46
S fazına ilerlemek için SiklinE-CDK2 kompleksinin oluşması gerekir.
Hücre siklusunda G1 / S sınır noktasında ilerleme S fazına ilerlemek için SiklinE-CDK2 kompleksinin oluşması gerekir. E2F’in aktive olması ile siklinE sentezi başlamıştır

47
Hücre siklusunda G2 / M sınır noktasında ilerleme
M fazına ilerlemek için SiklinA-CDK2 kompleksinin oluşması gerekir. E2F’in aktive olması ile siklinA sentezi başlamıştır

48
Mitozdan çıkma SiklinB-CDK1 inaktivasyonunu gerektirir.
Yeni bölünen hücreler; Tekrar G1’e dönebilir Sessiz döneme girebilir

49
SİKLİN BAĞIMLI KİNAZLAR
CDK4 Siklin D ile kompleks oluşturur Kompleks RB’yi fosforile eder Hücre G1’de kontrol noktasına ilerler CDK2 G1 geç döneminde Siklin E ile kompleks oluşturur G1/S geçişinde rol alır S fazında Siklin A ile kompleks oluşturur G2/M geçişini kolaylaştırır CDK1 Siklin B ile kompleks oluşturur G2/M geçişinde rol oynar

50
HÜCRE SİKLUSU İNHİBİTÖRLERİ
Cip/Kip INK4/ARF * Tümör süpressör gibi davranırlar

51
Cip/Kip ailesi p21, p27, p57 İnaktivasyon:
Siklin-CDK kompleksine bağlanarak sağlanır

52
p21’in kopyalama aktivitesi :
p53’ün kontrolü altındadır Hücre siklusunda p53: Hasarlı hücrelerin ilerlemelerini durduran veya yavaşlatan kontrol mekanizmalarını tetikler Apopitozuna yol açar

53
INK4a/ARF Gen lokusu iki protein kodlar 1. p16INK4a 2. p14ARF
Hücre siklusunu bloke ederek tümör süpresör gen gibi davranır

54
p16INK4a Sonuç: Hücre siklusu G1 sonunda kalır
SiklinD ile CDK4’e bağlanmak için rekabete girer. SiklinD-CDK4 kompleksi oluşumunu inhibe eder. RB fosforilasyonu engellenir. Sonuç: Hücre siklusu G1 sonunda kalır

55
p14ARF p53 parçalanmasını önleyerek hücre siklusunu bloke eder.

56
HÜCRE SİKLUSU İNHİBİTÖRLERİ
Cip/Kip ailesi: P21, p27 Siklin-CDK kompleksine bağlanarak hücre siklusunu bloke eder p21, p53 tümör süpresörü ile uyarılır p21, TGF gibi büyüme süpressörlerine yanıt verir INK4/ARF ailesi: p16INK4A, p14 p16INK4A, siklinD-CDK4’e bağlanır. RB’un inhibitör etkisini kolaylaştırır p14ARF, MDM2 aktivitesini inhibe ederek p53 seviyesini arttırır

57
melanom, glioblastoma, NSCLC,
tumor suppressor p16 akut lenfoid lösemiler, T and B hücre non-Hodgkin lenfomalar onkogen cdk4 glioblastoma, melanom Meme kanseri, özefagus kanseri, onkogen hepatoselüler karsinom cyclin D retinoblastoma, osteosarkoma, tumor suppressor pRB NSCLC

58
SİTOTOKSİK AJANLARIN ETKİ YERLERİ Hücre siklusu düzeyinde
Antibiotikler Antimetabolitler S (2-6h) G2 (2-32h) Vinka alkaloidleri M (0.5-2h) Mitoz inhibitörleri Taksoidler Alkilleyiciler G1 (2-h) G0
 G2. (2-32h) Vinka alkaloidleri. M. (0.5-2h) Mitoz inhibitörleri. Taksoidler. Alkilleyiciler. G1. (2-h) G0.")
59
Kemoterapötiklerin Etki Yerleri
Faz Spesifikler G1 fazı: L-Asp., pred. S fazı: Antimetabolitler G2 fazı: Bleomisin, etoposid M fazı: Vinkristin, vinblastin, taxanlar Faz spesifik olmayanlar Alkilleyiciler Cisplatin Antititümör antibiyotikler Steroid hormonlar

60
SİTOTOKSİK AJANLARIN ETKİ YERLERİ hücresel düzeyde
DNA sentezi Antimetabolitler DNA Alkilleyiciler DNA transkripsiyonu DNA duplikasyonu Mitoz Topoizomeraz inhibitörleri Mikrotübül inhibitörleri

61
SİTOTOKSİK AJANLARIN ETKİ YERLERİ
PÜRİN SENTEZİ PİRİMİDİN SENTEZİ 6-MERKAPTOPURIN 6-THIOGUANIN METOTREKSAT 5-FLUOROURASİL HİROKSİÜRE SİTARABİN RNA DNA ALKİLLEYİCİLER ANTiBiYOTiKLER DNA ETOPOSİD RNA L-ASPARAGİNAZ VİNKA ALKALOİDİ TAKSOİDLER PROTEİNLER ENZİMLER MİKROTÜBÜLLER

Benzer bir sunumlar

>")








 ZM106 Biyokimya 11. Hafta.>")





>")


