Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Proteinlerin üç boyutlu yapısı

2
Bir proteinin kovalent iskeleti yüzlerce tek bağdan oluşmaktadır
Bir proteinin kovalent iskeleti yüzlerce tek bağdan oluşmaktadır. Bu bağların birçoğunun etrafında serbest rotasyon mümkün olduğu için proteinin sınırsız sayıda konformasyonda bulunduğu düşünülebilir. Ancak her proteinin özgün bir üç-boyutlu yapıya sahip olduğunu kuvvetle düşündürmektedir. Hemoglobin ve üreaz enzimi dahil pek çok protein kristallendirilmişti. Bir kristal yapı içinde moleküllerin düzenli bir diziliminin, genellikle sadece moleküler birimler tıpatıp aynı olduğunda, oluşabildiği bilinmektedir.

3
Bu bölümde, proteinlerin üç-boyutlu yapısını, beş temayı vurgulayarak inceleyeceğiz. Birincisi bir proteinin üç- boyutlu yapısı, proteinin amino asit dizilimi tarafından belirlenir. İkincisi, bir proteinin işlevi yapısına bağlıdır. Üçüncüsü, izole edilmiş bir protein eşsiz veya eşsize yakın bir yapıya sahiptir. Dördüncüsü, belirli bir proteinin özgül yapısının korunmasını sağlayan sabitleyici en önemli kuvvet kovalent olmayan etkileşimleridir. Sonuç olarak, çok sayıda özgün protein yapıları arasında, protein mimarisi konusundaki bilgilerimizi düzenlememize yardımcı olur. Protein işlevi çoğu kez iki veya daha fazla yapısal biçim arasında değişimi gerektirir

4
Protein yapısına bakış
Bir proteinin yapısında bulunan atomların uzaysal düzenlenimi proteinlerin konformasyonu olarak adlandırılmaktadır. Proteinin olası konformasyonları, kovalent bağları kırılmaksızın elde edilebilen herhangi bir yapısal durumudur. Örneğin, tek bağlar etrafında rotasyon olması ile konformasyonda değişim oluşabilir. Teorik olarak mümkün olabilen çok sayıdaki konformasyonların, biyolojik şartlar altında, genellikle bir veya ikisi egemen olarak bulunur. Belirli şartlar altında mevcut olan konformasyon, çoğunlukla termodinamik olarak en kararlı, yani en düşük Gibbs serbest enerjisine (G) sahip olan konformasyondur.
 sahip olan konformasyondur.")
5
Protein konformasyonu büyük ölçüde zayıf etkileşimlerle sabitlenir
Protein yapısıyla ilişkili olan karalılık terimi, doğal konformasyonu sürdürme eğilimi olarak tanımlanabilir. Fizyolojik şartlar altında tipik proteinlerdeki katlanmış ve katlanmamış hali ayıran ∆G; 20 ile 65kJ/mol sınırları arasındadır. Belirli bir polipeptit zincirinin teorik olarak sayısız farklı konformasyonu olduğu düşünülebilir ve bir proteinin katlanmamış hali, konformasyonal entropisnin en yüksek olduğu durum olarak nitelendirilebilir. Tek bir kovalent bağın kırılabilmesi için yaklaşık olarak 200 ile 460 kJ/mol enerji gerekirken zayıf etkileşimler, 4ile 30 kJ/mol arasındaki önemsiz düzeydeki enerjiyle bozulabilir

6
Disülfit bağları gibi kovalent bağlar, şüphesiz tek bir zayıf etkileşimden çok daha kuvvetlidir.
Genelde en düşük serbest enerjili protein konformasyonu (yani en karalı konformasyon) en fazla sayıda zayıf etkileşimi içeren konformasyonlardan biridir. Protein karalılığına zayıf etkileşimlerin katkısı suyun özellikleri göz önüne alınarak anlaşılabilir. Saf su hidrojen bağı yapmış H2O moleküllerinin bir ağını içermektedir. Başka hiçbir molekül suyun hidrojen bağlama potansiyeline sahip değildir ve sulu çözeltide bulunan diğer moleküller, su molekülleri arasındaki hidrojen bağlarının bozulmasına neden olur.
 en fazla sayıda zayıf etkileşimi içeren konformasyonlardan biridir. Protein karalılığına zayıf etkileşimlerin katkısı suyun özellikleri göz önüne alınarak anlaşılabilir. Saf su hidrojen bağı yapmış H2O moleküllerinin bir ağını içermektedir. Başka hiçbir molekül suyun hidrojen bağlama potansiyeline sahip değildir ve sulu çözeltide bulunan diğer moleküller, su molekülleri arasındaki hidrojen bağlarının bozulmasına neden olur.")
7
Entropi terimi, sulu çözeltilerde hidrofobik grupların birleşmesini yönlendiren başlıca termodinamik itici kuvvettir. Fizyolojik koşullarda bir proteindeki hidrojen bağlarının ve iyonik etkileşimlerin oluşumu büyük ölçüde aynı entropik etki tarafından yönlendirilir. Polar gruplar genellikle suyla hidrojen bağı yaparlar ve bundan dolayı suda çözünürler. En polar molekülün bile çözünürlük sınırı vardır, çünkü bu moleküllerin çözeltideki varlığı birim kütle başına düşen hidrojen bağında bir azalmaya neden olacaktır.

8
Bir makromoleküldeki iki polar grup arsındaki moleküliçi hidrojen bağları veya iyonik etkileşimlerin oluşum enerjisi, büyük ölçüde, aynı gruplar ve su arasındaki herhangi bir etkileşimin uzaklaştırılmasıyla ortadan kaldırılabildiği halde, moleküliçi etkileşimlerin oluşumu sırasında yapılanmış olan su tabakasının bozulması, katlanma için güdümleyici bir entropik güç sağlar. Hidrofobik etkileşimler bir proteinin konformasyonunun sabitlenmesinde şüphesiz önemlidir. Genellikle hidrofobik amino asitlerin yan zincirleri proteinin iç kısmında, yoğun olarak paketlenmiş bir merkez oluşturur. Proteinlerdeki gruplar arasında oluşan hidrojen bağları kooperatif olarak biçimlenir. Protein konformasyonunun sabitlenmesine hidrojen bağları ve diğer kovalent olmayan etkileşimlerin katkısı halen değerlendirilmektedir. İki basit kuralı yansıtmaktadır: (1) hidrofobik karakterli amino asitler sudan uzakta, daha çok proteinin iç kısmına gömülü bulunur ve (2) protein yapısı içindeki hidrojen bağı sayısı maksimum düzeye çıkarılır.
 hidrofobik karakterli amino asitler sudan uzakta, daha çok proteinin iç kısmına gömülü bulunur ve (2) protein yapısı içindeki hidrojen bağı sayısı maksimum düzeye çıkarılır.")
9
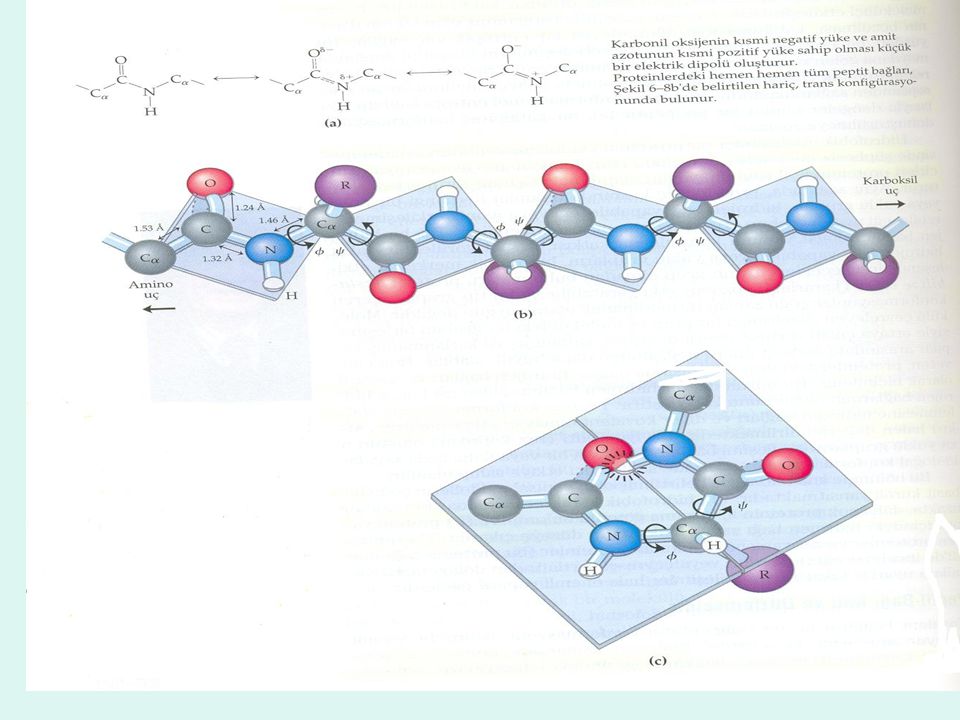
Peptit bağı katı ve düzlemseldir
1930’ların sonlarında, Linus Pauling ve Robert Corey protein yapısı hakkında şu andaki bilgilerimizin temelini oluşturan seri bir çalışma başlattılar.

12
Cα üzerindeki rotasyondan kaynaklanan bağ açıları N-- Cα için Ø (phi) ve Cα—C bağı için ψ (psi) olarak belirtilmektedir. Ø ve ψ değerlerinin ikisin inde Oo olduğu konformasyon bu sebepten dolayı engellenmektedir ve bu konformasyon sadece rotasyonun açılarını açıklamak için bir referans noktası olarak kullanılmaktadır. Ramachandran grafiğinde Ø ve ψ’in alabileceği değerler grafik olarak ortaya çıkar.

13
Proteinlerin ikincil yapısı
İkincil yapı terimi, polipeptidin belirli kısımlarının yerel konformasyonlarını ifade eder. İkincil yapının özellikle birkaç tipi karalıdır ve protein yapısında yaygın olarak bulunur. En yaygın olarak bulunanılar α heliks (α sarmal) ve β konformasyonlarıdır.1952’de bu ikincil yapının varlığını öngörmüşlerdir
 ve β konformasyonlarıdır.1952’de bu ikincil yapının varlığını öngörmüşlerdir.")
14
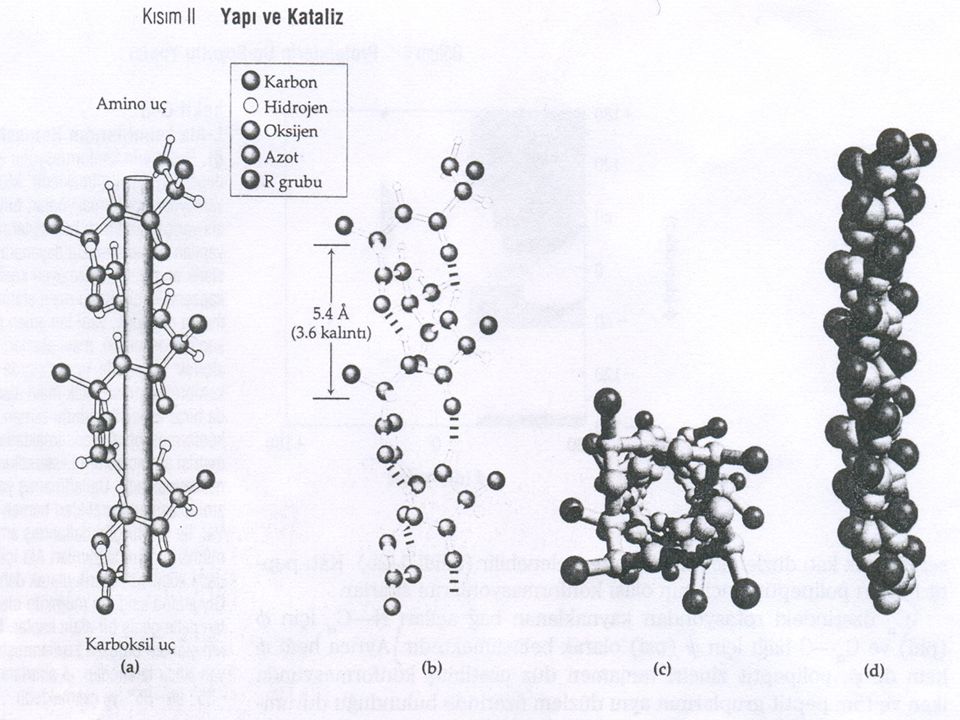
α Heliks yaygın bir protein ikincil yapısıdır
Pauling ve Corey William Astbury’in (1930) deneysel sonuçlarına sahiplerdi. Katı Peptit bağlarıyla (rotasyonu serbest olan tek bağlar dışında) polipeptit zincirinin varsayılabilen en basit düzlenimi Pauling ve Corey’in α heliks olarak adlandırıldıkları helezonik yapıdır. Tekrarlayan birim, eksen boyunca yaklaşık 5.4 Å uzunluğunda olan, heliksin tek bir dönüşüdür. Bir α helikste amino asitler ψ = -45o ile -50o ve Ø = -60o olduğu konformasyonlara sahiptir. Ve her bir heliks dönüşü 3.6 amino asit kalıntısı içermektedir.
 deneysel sonuçlarına sahiplerdi. Katı Peptit bağlarıyla (rotasyonu serbest olan tek bağlar dışında) polipeptit zincirinin varsayılabilen en basit düzlenimi Pauling ve Corey’in α heliks olarak adlandırıldıkları helezonik yapıdır. Tekrarlayan birim, eksen boyunca yaklaşık 5.4 Å uzunluğunda olan, heliksin tek bir dönüşüdür. Bir α helikste amino asitler ψ = -45o ile -50o ve Ø = -60o olduğu konformasyonlara sahiptir. Ve her bir heliks dönüşü 3.6 amino asit kalıntısı içermektedir.")
15
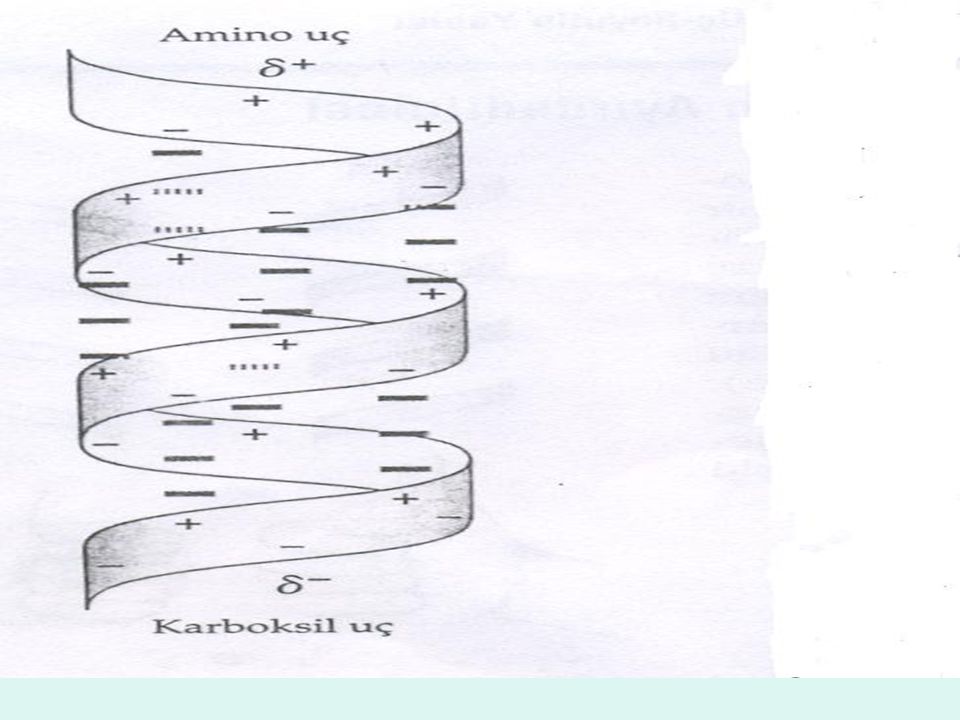
α Heliks yapısı niçin diğer birçok olası konformasyondan daha kolaylıkla oluşur? Yanıt, kısmen α heliks yapısının iç hidrojen bağlarının en uygun kullanımını sağlamasına bağlıdır. Bu yapı, Peptit bağının elektronegatif azot atomuna bağlı hidrojen atomu ile bu Peptit bağının amino terminal ucu kısmında bulunan dördüncü amino asidin elektronegatif karbonil oksijen atomu arasında bulunan bir hidrojen bağıyla sabitlenmektedir. Doğal olarak bulunan L- amino asitler gerek sağ-el dönüşlü, gerekse sol- el dönüşlü heliks yapısını oluşturabilir

17
Amino asit dizisi α heliksin kararlılığını etkiler
Polipeptitlerin hepsi kararlı bir α helliks yapısı oluşturamaz. Amino asitlerin yan zincirleri arasındaki etkileşimler, bu yapıyı stabilize veya destabilize edebilir.eğer bir polipeptit zinciri Glu’lardan oluşmuş uzun bir bölüm içeriyorsa, zincirin bu bölümü pH 7.0 ‘de α heliks yapısı oluşturmayacaktır.

18
Aynı nedenle eğer çok sayıda bitişik, pH 7
Aynı nedenle eğer çok sayıda bitişik, pH 7.0’de pozitif yüklü R grubuna sahip olan Lys ve / veya Arg amino asitleri varsa; bu amino asitlerde birbirini itecek ve α heliks oluşumunu önleyeceklerdir. Zincirde birbirlerine yakın oldukları zaman α heliksi detabilize edebilirler. Pozitif yüklü amino asitler genellikle negatif yüklü amino asitlerden üç amino asit kalıntısı uzakta bulunur ve bu yerleşim bir iyon çifti oluşumuna olanak verir. İki aromatik amino asit de hidrofobik etkileşimler nedeniyle sıklıkla benzer aralıkta bulunur

19
Prolin amino asidindeki azot atomu katı halka yapısının bir kısmını oluşturduğundan N-Cα bağının etrafında rotasyon mümkün değildir. Bu yüzden, Pro kalıntısı α heliks yapısının kararlılığını bozan bir kıvrım oluşturur. Bu nedenlerden dolayı prolin α heliks yapısında çok nadir olarak bulunmaktadır. Glisin faklı bir nedenle α heliks yapısında sık bulunmaz. Daha fazla konformasyonal esnekliğe sahiptir. Glisin polimerlerinin oluşturduğu sarmal yapı α heliks yapısından oldukça farklıdır. Α heliks yapısının kararlılığını etkileyen son faktör; α –helezonik parçanın sonuna yakın bölgede bulunan amino asitlerin özelliğidir.

23
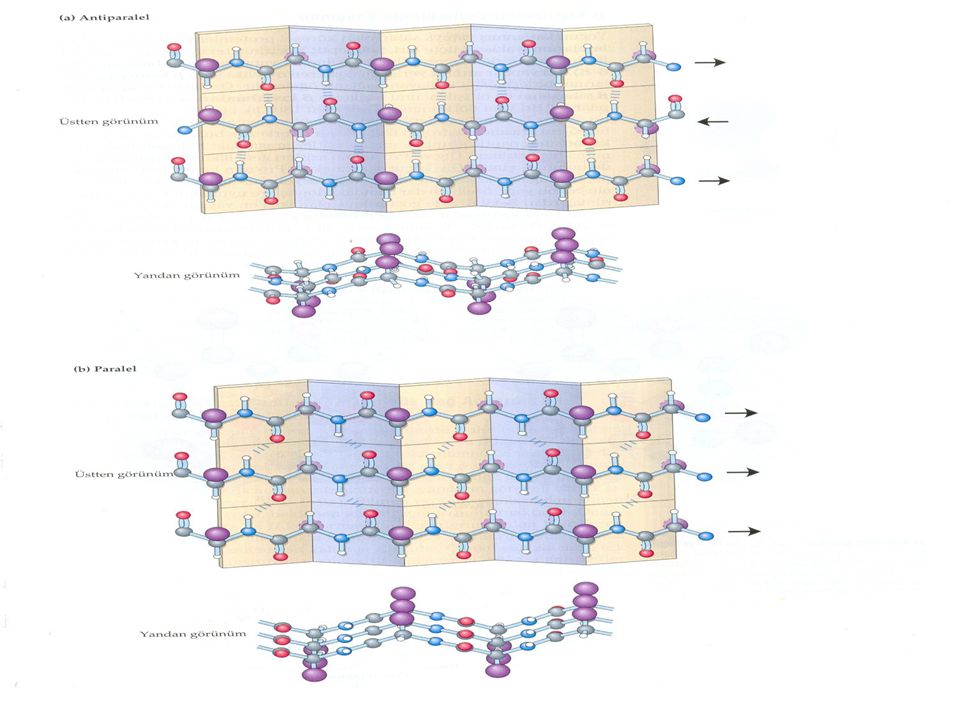
β konformasyonu polipeptit zincirini tabakalar şeklinde düzenler
İpek fibroini ve örümcek ağı fibroini gibi β- keratinlerin, en küçük R gruplarına sahip iki amino asit olan Gly ve Ala içerikleri çok yüksektir.

Benzer bir sunumlar





 ile karakterize edilir. Verilen bir elementin tüm atomlarında.>")











>")





