Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
LİPİD METABOLİZMASI BBP108-Bitki Biyokimyası 10. Hafta
Doç.Dr. Yıldız Aka Kaçar Ç.Ü.Z.F.Bahçe Bitkileri Bölümü

2
Bitkilerde yağlar enerji kaynağı olarak nerede önemli olabilir???
Hayvanlar yağları enerji deposu olarak kullanmalarına karşılık bitkiler genellikle karbon deposu olarak kullanırlar. Bitkilerde yağlar enerji kaynağı olarak nerede önemli olabilir??? Katı ve sıvı yağlar, pamuk, fıstık, ayçiçeği ve soya fasulyesi gibi tarımsal olarak önemli türleri içeren pek çok tohumda indirgenmiş karbonun önemli depolama şeklidir. Sıvı yağlar, küçük tohumlara sahip yabani bitkilerde en büyük depolama görevi görürler. Ayrıca zeytin avokado gibi bazı meyveler de katı ve sıvı yağları depolarlar.

3
Bu bölümde gliserolipidlerin iki tipinin sentezi açıklanacaktır.
Triaçil gliseroller (tohumlarda depolanan katı ve sıvı yağlar) Polar gliserolipidler (hücresel zarların ikili tabakasını oluşuran lipidler) Triaçilgliserol ve polargliserolipidlerin biyosentezinde plastidler ve endoplazmik retikulum olmak üzere iki organel ortak rol oynamaktadır.
 Polar gliserolipidler (hücresel zarların ikili tabakasını oluşuran lipidler) Triaçilgliserol ve polargliserolipidlerin biyosentezinde plastidler ve endoplazmik retikulum olmak üzere iki organel ortak rol oynamaktadır.")
4
Sıvı ve Katı Yağlar Büyük Miktarda Enerji Depolarlar
Lipidler karbonhidratlardan çok daha fazla indirgenmiş karbon formları içerir. Örneğin bir gram sıvı ve katı yağın tam oksidayonu sonucunda (yaklaşık 9.3 kcal’lik bir enerji), bir gram nişastanın oksidasyonundan (3.8 kcal) çok daha fazla ATP üretilir. Bununla birlikte sıvı ve katı yağlar ve zar fosfolipidlerin biyosentezi için çok daha fazla enerjiye ihtiyaç duyulur.
, bir gram nişastanın oksidasyonundan (3.8 kcal) çok daha fazla ATP üretilir. Bununla birlikte sıvı ve katı yağlar ve zar fosfolipidlerin biyosentezi için çok daha fazla enerjiye ihtiyaç duyulur.")
5
Triaçilgliserollar Oleozomlarda Depolanır
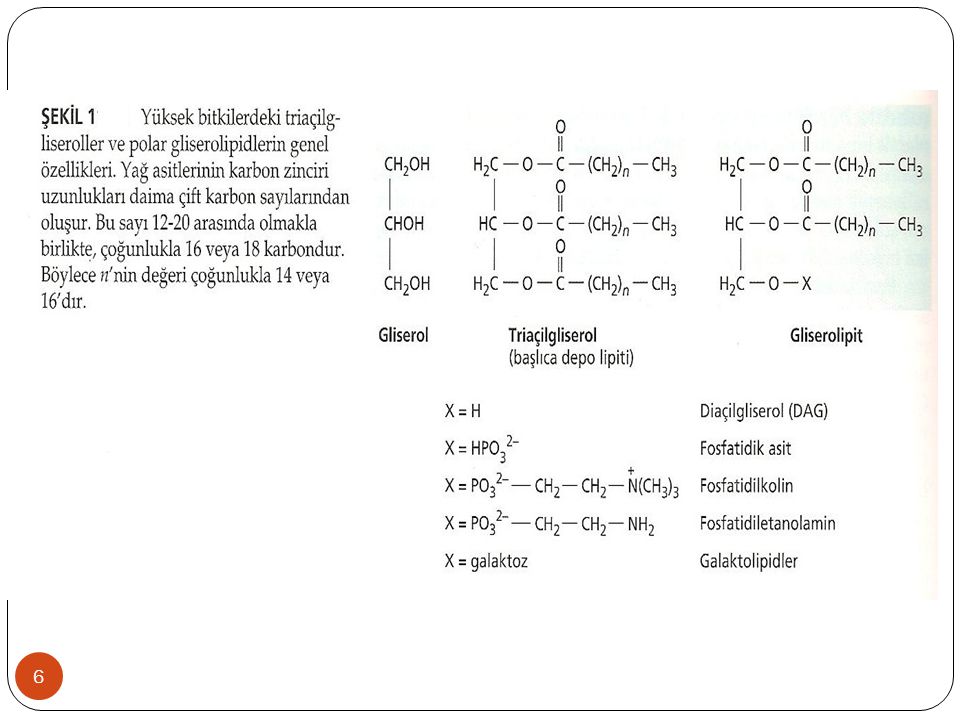
Sıvı ve katı yağlar esas olarak gliserolün 3 hidroksil grubuna ester bağları ile bağlanan yağ asidi moleküllerini içeren trigliserit veya triaçilgliserol (açil yağ asidi kısmını ifade eder) yapısında bulunurlar (Şekil 1). Bitkilerdeki yağ zincirleri çok sayıdaki karbon atomuna sahip düz zincirli karboksilik asitlerdir. Bu zincirlerin en uzunu 20 en kısası ise 12 karbon biriminden oluşur. Ancak çoğunlukla bitkilerde bulunan yağ asitleri 16 veya 18 karbonludurlar. Bitki lipidlerinde bulunan temel yağ asitleri Şekil 2 de gösterilmiştir.
 yapısında bulunurlar (Şekil 1). Bitkilerdeki yağ zincirleri çok sayıdaki karbon atomuna sahip düz zincirli karboksilik asitlerdir. Bu zincirlerin en uzunu 20 en kısası ise 12 karbon biriminden oluşur. Ancak çoğunlukla bitkilerde bulunan yağ asitleri 16 veya 18 karbonludurlar. Bitki lipidlerinde bulunan temel yağ asitleri Şekil 2 de gösterilmiştir.")
7
Şekil 2. Yüksek Bitki dokularındaki genel yağ asitleri
Bitki lipidlerin yapısında bulunan yağ asitlerinin bileşimi bitki türüne göre değişir. Örneğin fıstık yağı: %9 palmitik asit, %59 oleik asit ve %21 linoleik asitten, pamuk yağı: %20 palmitik asit, %30 oleik asit ve %45 linoleik asitten oluşur.

8
Birçok tohumda bulunan triaçilgliseroller ya kotiledon ya da endosperm hücrelerinin sitoplazmasında oleozomlar (sferozomlar ve yağ cisimleri olarak da bilinirler) olarak adlandırılan organeller içerisinde depolanırlar. Olezomlar sitoplazmadan trigliseritleri ayıran tekli bir zarla çevrilmişlerdir. Tek fosfolipid tabakası (yani ikili tabakanın yarısı) sitosol ile temasta olan fosfolipidlerin hidrofilik uçlarına sahip bu yağ cismini kuşatır ve hidrofobik açil karbon zincirleri triaçilgliserollerin iç kısmına doğru yönelir. Bu özelliklere sahip olan oleozom, yüzeyini kaplayan ve oleozin olarak adlandırılan özel proteinlerin varlığı ile kararlılık kazanır.
 sitosol ile temasta olan fosfolipidlerin hidrofilik uçlarına sahip bu yağ cismini kuşatır ve hidrofobik açil karbon zincirleri triaçilgliserollerin iç kısmına doğru yönelir. Bu özelliklere sahip olan oleozom, yüzeyini kaplayan ve oleozin olarak adlandırılan özel proteinlerin varlığı ile kararlılık kazanır.")
9
Şekil 3. A B Şekil 3. B’ de ER’nun fosfolipid çift tabakasının
içerisinde yağların sentezlenmesi ve Biriktirilmesi sonucu oleozomların oluştuğu görülmektedir. Olezomlar ER’den tomurcuklandıkatan sonra oleozin isimli bir protein tabakasıyla kuşatılmışlardır.

10
Polar Gliserolipidler Zarlardaki Yapısal Lipidlerdir
Zarlardaki esas yapısal lipidler, polar gliserolipidler olup iki alt grubu vardır. 1. Gliseroglikolipidler Ana grup olarak şekerlere sahiptir. 2. Gliserofosfolipidler Ana grup olarak fosfat grubu içerirler. Lipidlerin ayrıca fotosentezde ve diğer metabolik işlevlerde özel rolleri vardır. Fotosentetik dokulardaki zar lipidlerinin %70’ini bulunduran kloroplast zarları da gliserolipidlerden oluşurlar.Hücrenin diğer zarları ise gliserofosfolipidleri içerirler.

11
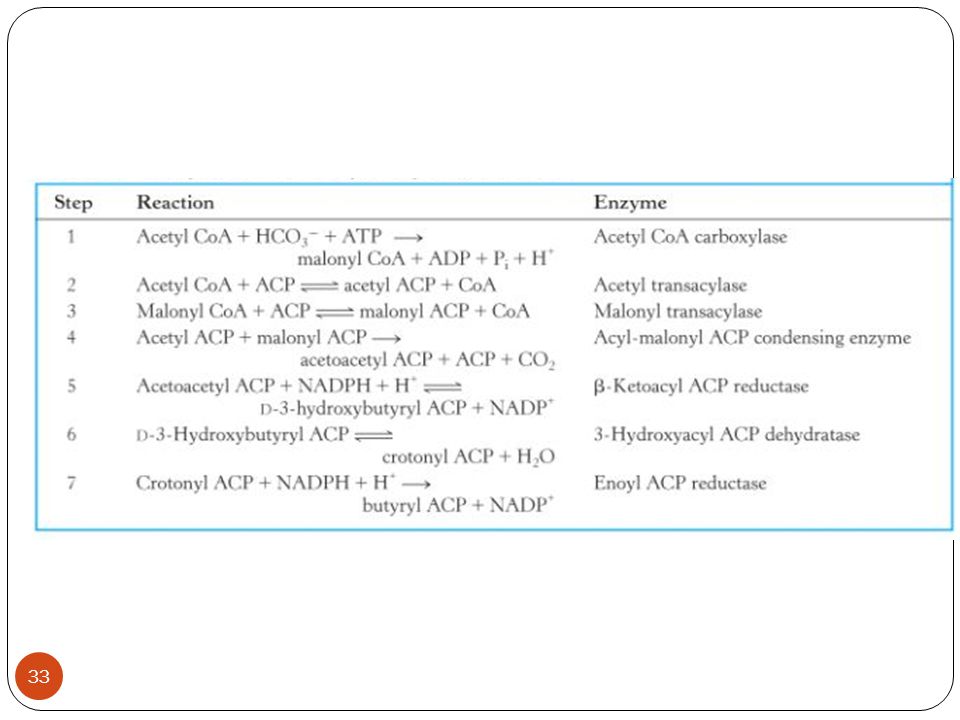
Yağ asidi Biyosentezi Bitkilerde yağ asitleri yüksek oranda plastidlerde sentezlenirken, hayvanlarda yağ asitleri genellikle sitoplazmada sentezlenir. Yağ asidi biyosentezinde rol oynayan enzimler kompleks halinde birarada tutulmakta ve ortak isimle yağ asidi sentaz olarak adlandırılmaktadır. Bu enzim kompleksi enzimlerin ayrı ayrı ayrı çalışmasından çok daha etkin bir biçimde reaksiyonu katalizler. Uzayan açil zincirleri açil taşıyıcı protein (ACP) olarak adlandırılan düşük molekül ağırlıklı asidik proteine kovalent bağ ile bağlanır. Açil taşıyıcı proteine bağlanan yağ asidi zinciri açil ACP olarak adlandırılır.
 olarak adlandırılan düşük molekül ağırlıklı asidik proteine kovalent bağ ile bağlanır. Açil taşıyıcı proteine bağlanan yağ asidi zinciri açil ACP olarak adlandırılır.")
12
yağ asidi sentaz enzimi

13
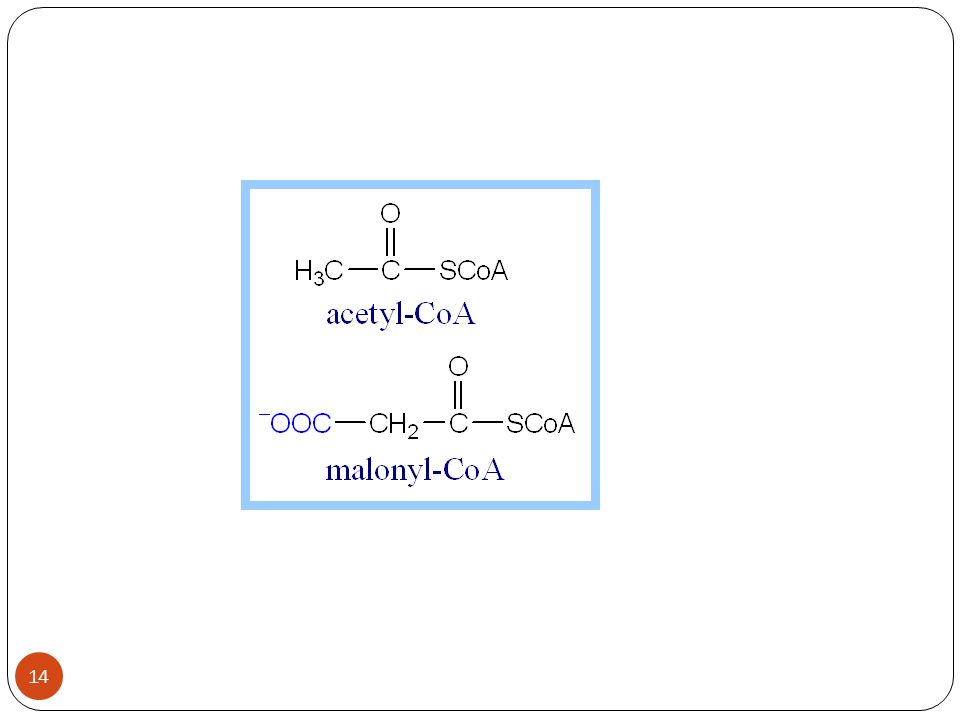
Yağ asidi biyosentezi Yağ asidi sentezi yolunda gerçekleşen ilk adım Asetil CoA karboksilaz enzimi yardımıyla asetil CoA ve CO2’den malonil CoA’nın sentezlenmesidir. Daha sonra Malonil CoA, malonil ACP’yi oluşturmak için ACP ile reaksiyona girer.

15
Yağ asidi biyosentezi için, asetil KoA ile birlikte NADPH, ATP, Mn2+, CO2 kaynağı olarak HCO3 ve biotin gereklidir.

16
Yağ asidi biyosentezinin ilk basamağı, asetil KoA’nın irreversibl (dönüşümsüz) bir reaksiyonla malonil KoA’ya karboksilasyonudur.
 bir reaksiyonla malonil KoA’ya karboksilasyonudur.")
17
Yağ asidi biyosentezi 1. Yağ asidi sentezinin birinci döngüsünde, AsetilCoA’dan asetat grubu kondanse edici enzimdeki belirli bir sisteine aktarılır ve daha sonra asetoasetil –ACP’yi oluşturmak için malonil-ACP ile birleşir. 2. Daha sonra karbon 3’teki keto grubu, üç enzimin etkisiyle 4 karbon uzunluğundaki yeni bir açil zincirini(bütiril-ACP) oluşturmak için uzaklaştırılır (indirgenir).
 oluşturmak için uzaklaştırılır (indirgenir).")
18
Yağ asidi biyosentezi 3. Bu 4 karbonlu asite kondanse edici enzimin katalizörlüğü ile diğer malonil –ACP molekülünden iki karbonlu birimin eklenmesi sağlanarak zincir uzunluğu karbon oluncaya kadar devam eder. 4. Bazı 16:0 –ACP’ler yağ asidi sentaz sisteminden ayrılır. Ancak 18:0-ACP’ye uzayan çoğu moleküller etkili bir biçimde desaturaz enzimi tarafından 18:1-ACP’ye dönüştürülür. Bu işlemin tekrarlanması ile plastidlerdeki yağ asidi sentezinin ara ürünleri oluşturulur.

19
Şekil 4. Bitki hücrelerini plastidlerindeki yağ asidi sentezi döngüsü

20
Çimlenen Tohumlardaki depo lipidler Karbonhidratlara dönüştürülürler
Çimlenme sonrası yağ içeren tohumlar, lipidleri sukroza dönüştürmek sureti ile triaçilgliserolleri metabolize ederler. Bitkiler tohumların endosperminde bulunan yağlarını çimlenen fidelerin kök ve gövdelerine taşıyamazlar. NEDEN??

21
Çimlenen Tohumlardaki depo lipidler Karbonhidratlara dönüştürülürler.
Çünkü; depolanmış lipidler daha hareketli bir karbon formuna (genellikle sukroz) dönüştürülmelidir. Bu işlem farklı hücresel bölümlerde yer alan birkaç reaksiyonla gerçekleştirilir.
 dönüştürülmelidir. Bu işlem farklı hücresel bölümlerde yer alan birkaç reaksiyonla gerçekleştirilir.")
22
Lipidlerin Sukroza dönüştürülmesi
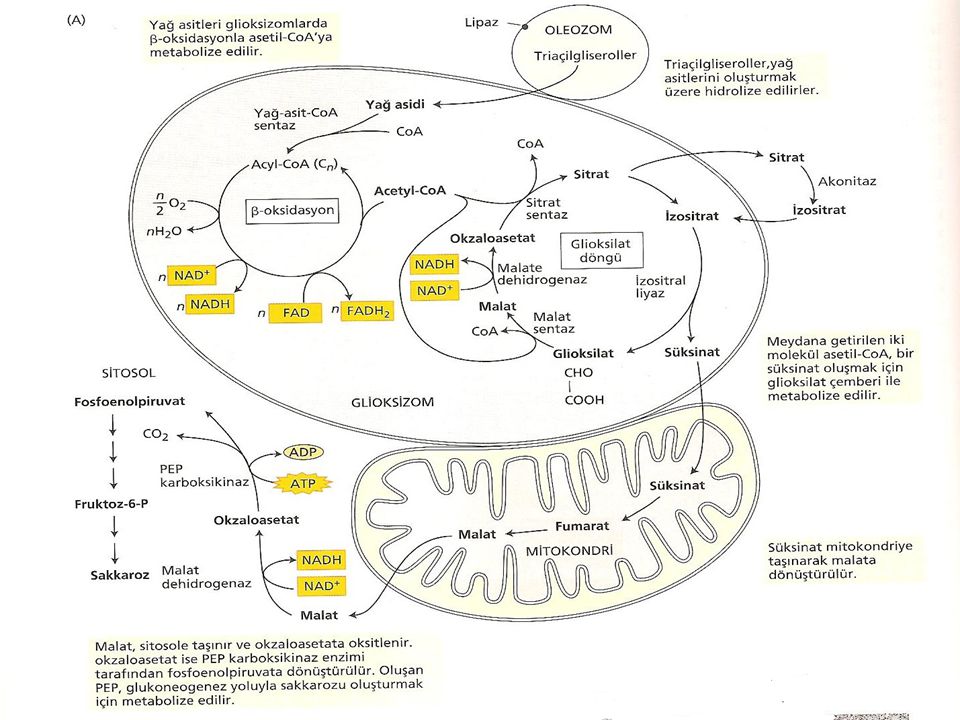
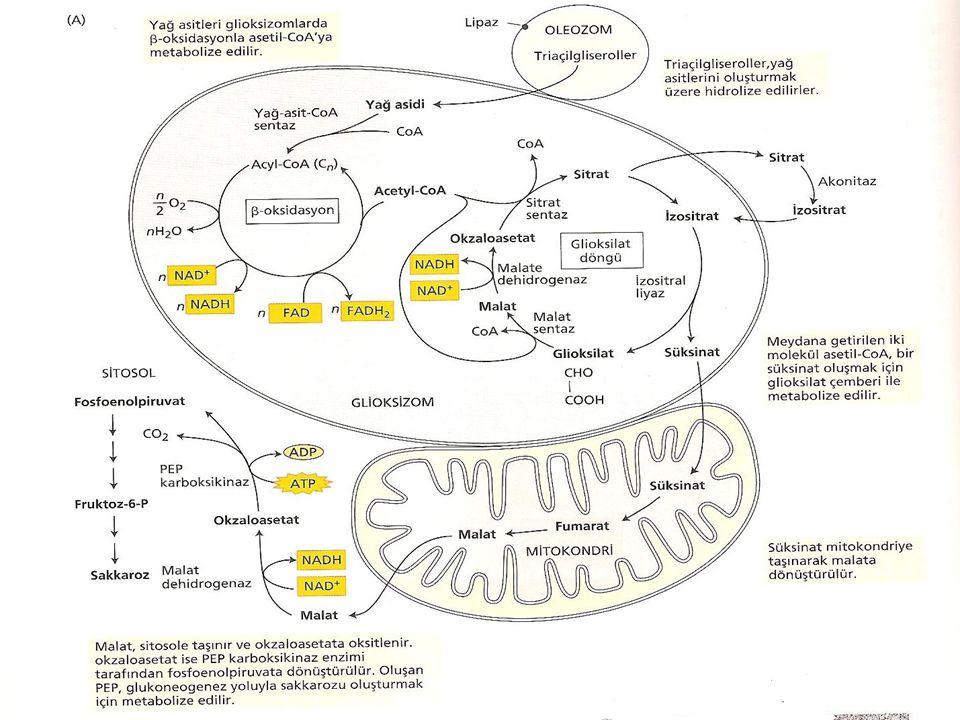
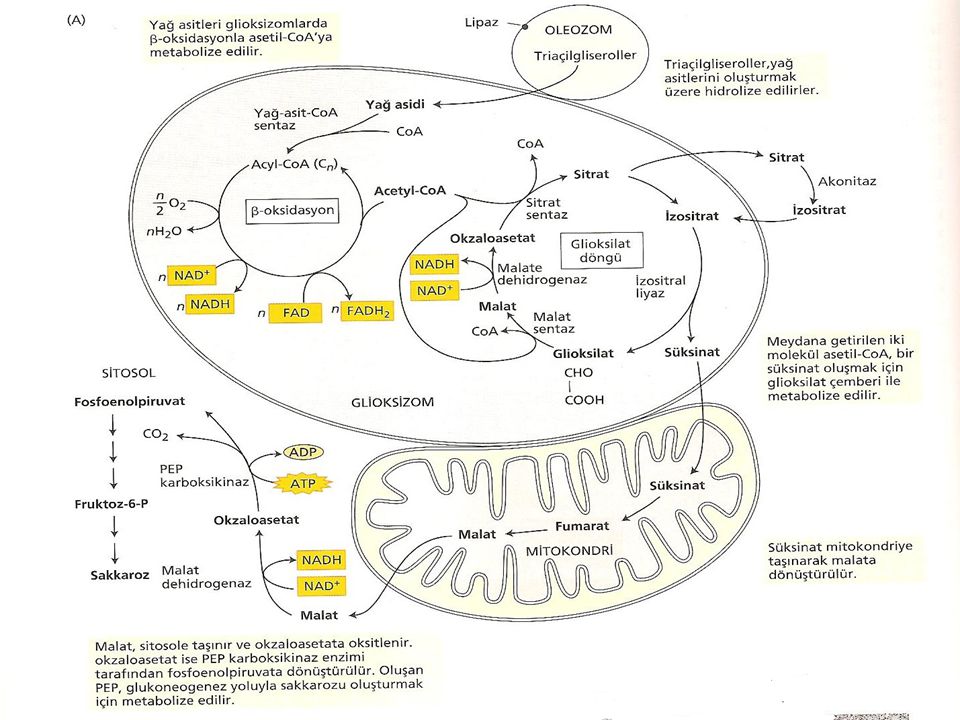
Yağ içeren tohumlarda lipidlerin sukroza dönüştürülmesi çimlenme ile başlar. Yağ cisimlerinde depolanan triaçilgliserollerin serbest yağ asidine hidrolizi ile başlayan işlemler zinciri bu yağ asitlerinin, asetil-CoA’yı oluşturmak üzere oksidasyona uğraması ile devam eder (Şekil). Yağ asitleri glioksizom olarak adlandırılan bir peroksizom çeşidinde okside olurlar. Glioksizom tohumların yağ bakımından zengin dokularında bulunan tek tabakalı bir zarla çevrilmiş organeldir.
. Yağ asitleri glioksizom olarak adlandırılan bir peroksizom çeşidinde okside olurlar. Glioksizom tohumların yağ bakımından zengin dokularında bulunan tek tabakalı bir zarla çevrilmiş organeldir.")
23
Lipidlerin Sukroza dönüştürülmesi
Acetil CoA, süksinatı oluşturmak için glioksizomlarda metabolize edilmektedir. Bu formda glioksizomlardan mitokondriye taşınmakta ve orada ilk olarak okzaloasetata ve daha sonra da malata dönüşmektedir. Bu işlemler glukogenesis yoluyla malatın glukoza dönüşmesiyle sitoplazmada son bulur ve daha sonra glukozdan sukroz oluşur.

25
Bir kabak fidesinin yağ depolayan kotiledonlarındaki bir hücrenin
Elektron mikroskobundaki görünümü

26
Lipaz Hidrolizi Lipidlerin karbonhidratlara dönüşmesinin başlangıç aşamasında, lipaz enzimi tarafından yağ cisimlerinde depolanan trigliseritler parçalanır. Lipaz enzimi triaçilgliserolleri gliserole ve 3 molekül yağ asidine hidroliz eder.

28
Yağ asitlerinin β oksidasyonu
Triaçilgliserollerin hidrolizinden sonra, meydana gelen yağ asitleri glioksizoma girer ve orada yağ –açil-CoA’ya dönüştürülerek aktifleştirilir. Yağ-açil-CoA, β oksidasyon reaksiyonları için bir başlangıç substratıdır.Burada Cn yağ asitleri (n sayıda karbondan oluşmuş yağ asitleri) bir dizi reaksiyonla n/2 sayıdaki asetil – CoA molekülüne yıkılırlar. Bu reaksiyon sonunda her bir asetilCoA için 1 NADH ve 1 FADH2 oluşur ve ½ O2’i H2O’ya indirgenir.
 bir dizi reaksiyonla n/2 sayıdaki asetil – CoA molekülüne yıkılırlar. Bu reaksiyon sonunda her bir asetilCoA için 1 NADH ve 1 FADH2 oluşur ve ½ O2’i H2O’ya indirgenir.")
30
Glioksilat Döngüsü Glioksilat döngüsünün işlevi, 2 molekül asetilCoA’yı süksinata dönüştürmektir. β oksidasyonla üretilen asetil CoA glioksilat devrini oluşturan bir dizi reaksiyonla öncelikle glioksizomlarda metabolize edilir. Başlangıç olarak asetil CoA, sitratı oluşturmak için okzaloasetatla reaksiyona girer. Daha sonra izositrata izomerize olmak için sitoplazmaya transfer edilir. İzositrat tekrar peroksizoma alınır ve glioksilat oluşum yoluna özgü 2 reaksyonla malata dönüştürülür.

31
Glioksilat Döngüsü İlk olarak izo sitrat (C6), süksinat (C4) ve glioksilatı (C2) oluşturmak için izositrat liyaz enzimi tarafından parçalanır. Oluşan süksinat mitokondriye geçer. Daha sonra malat sentaz, malatı oluşturmak için asetil CoA’nın ikinci bir molekülü ile glioksilatı birleştirir. Malat daha sonra malat dehidrogenaz yardımıyla döngüyü sürdürmek için diğer Asetil CoA ile birleşebilen okzaloasetata yükseltgenir. Meydana gelen glioksilat gliosizomlardaki çembere katılır. Süksinat ise daha sonraki işlevlerde kullanılmak üzere mitokondriye taşınır.
, süksinat (C4) ve glioksilatı (C2) oluşturmak için izositrat liyaz enzimi tarafından parçalanır. Oluşan süksinat mitokondriye geçer. Daha sonra malat sentaz, malatı oluşturmak için asetil CoA’nın ikinci bir molekülü ile glioksilatı birleştirir. Malat daha sonra malat dehidrogenaz yardımıyla döngüyü sürdürmek için diğer Asetil CoA ile birleşebilen okzaloasetata yükseltgenir. Meydana gelen glioksilat gliosizomlardaki çembere katılır. Süksinat ise daha sonraki işlevlerde kullanılmak üzere mitokondriye taşınır.")
34
Yağ asidi sentezi için gerekli asetil KoA, glukoz, bazı amino asitler ve yağ asitlerinden mitokondride oluşmaktadır. Yağ asidi sentezi ile ilgili enzimler sitozolde bulunur. Mitokondride oluşan asetil KoA’nın yağ asidi sentezine katılabilmesi için sitozole geçmesi gerekir.

35
Yağ asidi sentezi için gerekli olan NADPH’nin başlıca iki kaynağı vardır:
Birinci ve en önemli NADPH kaynağı, glukozun pentoz fosfat yolunda yıkılımıdır. İkinci NADPH kaynağı, sitoplazmada oksaloasetatın indirgenmesiyle oluşan malatın malik enzim etkisiyle pirüvata oksidatif dekarboksilasyonudur.

36
Yararlanılan Kaynak: Bitki Fizyolojisi (Taiz&Zeiger) Ed. Prof.Dr. İsmail Türkan
 Ed. Prof.Dr. İsmail Türkan")
Benzer bir sunumlar