Sunuyu indir
1
Prof.Dr.Akgün YAMAN Mikrobiyoloji Anabilim Dalı
KOMPLEMAN SİSTEMİ Prof.Dr.Akgün YAMAN Mikrobiyoloji Anabilim Dalı

2
Kompleman Sistemi C, tek bir protein olmayıp 30’u aşkın protein komponentinden oluşur. C proteinleri reaksiyondan ziyade keşfedilme sıralarına göre numaralandırılır. Plazma globulinlerinin %10 kadarı C proteinleridir.

3
Kompleman Sistemi Zararlı patojenleri vücuttan uzaklaştırır.
Humoral bağışıklıkta ve iltihap oluşumunda rolü vardır. Yani; İmmün ve enflamatuvar reaksiyonlara katılmak, bu reaksiyonları amplifiye etmek ve güçlendirmek. Doğuştan bağışıklık sisteminin bir parçasıdır.

4
Kompleman Sistemi Tarihçe
Kompleman sisteminin tanınması, ilk olarak 1888’de Nuttal ile başlamıştır. Memelilerin taze plazma ya da serumunun bazı bakteriler üzerinde eritici etkisi olduğunu bulmuştur. Bu etkinin 56ºC’ de 30 dakika ısıtmakla veya uzun süre bekletmekle kaybolduğunu gözlemiştir.

5
Kompleman Sistemi Pfeiffer 1894’de, önce kobayları kolera vibriyonuna karşı bağışıklamıştır. Bu bağışık kobayların peritonuna kolera vibriyonu kültürü enjekte etmiştir. Belirli aralıklarla aldığı periton sıvısı örneklerini incelemiştir. Kolera vibriyonlarının, deney tüpünde bağışık taze serumla karıştıklarında eridiklerini, ama bağışık serum beklemiş (bayat) olursa, erime olayının gerçekleşmediğini görmüştür.
 olursa, erime olayının gerçekleşmediğini görmüştür.")
6
Kompleman Sistemi Bayat veya ısıtılmış taze serum + Kolera vibriyonu + bağışık olmayan taze normal kobay serumu vibriyonlar erirler. Kolera vibriyonlarının erimesi için, kendilerine karşı oluşturulmuş spesifik antikorları taşıyan bağışık serum gereklidir, ama yeterli değildir. Bayatlamakla ya da 56º C’ de 30 dakika ısıtmakla etkisizleşen, antikor dışında ve bağışıklıkla ilgisi olmayan bir etkenin bulunduğu bu deneylerle ortaya çıkmıştır.

7
Kompleman Sistemi Enfeksiyona yanıtta ilk aktive olan sistemlerden birisidir. Hücre aktivasyonu Sitoliz Opsonizasyon

8
Kompleman Genleri C2, Faktör B ve C4 genleri, insanda ve diğer türlerde 6. kromozom üzerinde MHC sınıf II (HLA-DR) ile MHC sınıf I arasında bulunmaktadır. Bunlara MHC sınıf III genleri denir. γ- IFN, IL-1, IL-6 ve TNF gibi sitokinler kompleman proteinlerinin sentezini uyarırlar
 ile MHC sınıf I arasında bulunmaktadır. Bunlara MHC sınıf III genleri denir. γ- IFN, IL-1, IL-6 ve TNF gibi sitokinler kompleman proteinlerinin sentezini uyarırlar.")
9
Kompleman proteinleri Sentezlenme Yerleri
Hepatositler Monosit/Makrofaj hücreleri Hematopoetik hücreler Fibroblastlar Endotelial hücreler Adipositler Astrositler Nöronlar

10
Kompleman Sistemi Kompleman proteinleri
Normalde plazma da inaktif haldedir. Etkinliği için C fiksasyonu ve aktivasyonu gerekli Dolayısıyla aktivatör molekülün büyüklüğü önemli Ag-Ab. agregatları C.ı iyi fikse eder. Aktive olan komponent ve kompleks (C1, C4b2a)
")
11
Kompleman Proteinleri
Kompleman sistemindeki bu protein yapısındaki komponentler gerçekte proenzim gibi düşünülebilir. Kompleman sisteminin ilk komponenti aktive olduğunda bu proenzim aktivitesi kazanır. Yani proenzim 1 enzim 1 izleyen komponenti (proenzim 2) enzim 2, bu da proenzim 3’ü enzim 3
 enzim 2, bu da proenzim 3’ü enzim 3.")
12
Kompleman Proteinleri
Bir önceki komponent kendisinden sonraki komponentin daima daha çok sayıda molekülünü aktive ederek etkili olur Çığ gibi büyüyen çok güçlü bir aktivasyon olur.

13
İsimlendirme C” – 11 kompleman proteinini göstermektedir
( C1q, C1r, C1s, C4, C2, C3, C5, C6, C7, C8, C9) Faktör – Alternatif yolun proteinlerine ‘Faktör’ denir ve tek harfle gösterilir (Örneğin Faktör B veya FB) Üst çizgi – enzimatik olarak aktif bir protein veya kompleksi gösterir.
 Faktör – Alternatif yolun proteinlerine ‘Faktör’ denir ve tek harfle gösterilir (Örneğin Faktör B veya FB) Üst çizgi – enzimatik olarak aktif bir protein veya kompleksi gösterir.")
14
İsimlendirme Küçük harfler – proteolitik bir ürün fragmanını tanımlar
Küçük başlangıç bölünme fragmanı ‘a’ fragmanı, büyük parça ‘b’ fragmanı (Örneğin C3a, C3b) Ancak bu kurala uymayan C2’ dir. C2b küçük fragman, C2a büyük fragman olarak isimlendirilir.
 Ancak bu kurala uymayan C2’ dir. C2b küçük fragman, C2a büyük fragman olarak isimlendirilir.")
15
İsimlendirme • “R” – kompleman sistemindeki reseptörleri tanımlamaktadır (CR1 veya C5aR). Kompleman reseptörleri ya bağlarına (Ör; C5a reseptörü) ya da Cluster of Differentiation (CD) Sistemine göre isimlendirilir (Örneğin C3b reseptörü; CD35 olarak isimlendirilir. Regulatör proteinler genelde molekülün fonksiyonel aktivitesiyle ilişkili isimlerin kısaltmasıyla adlandırılır) (Örneğin; Decay Accelerating Factor, DAF)
 ya da Cluster of Differentiation (CD) Sistemine göre isimlendirilir (Örneğin C3b reseptörü; CD35 olarak isimlendirilir. Regulatör proteinler genelde molekülün fonksiyonel aktivitesiyle ilişkili isimlerin kısaltmasıyla adlandırılır) (Örneğin; Decay Accelerating Factor, DAF)")
16
Kompleman Sisteminin Başlıca Komponentleri
Klasik yolda etkin olanlar C1q, C1r, C1s, C4, C2 Alternatif yolda etkin olanlar D, C3, B Membran atak kompleksi C5, C6, C7, C8, C9 Kontrol proteinleri (plazma proteinleri) Pozitif regülasyon yapanlar Properdin Negatif regülasyon yapanlar C I inhibitörü (CI INH) C4bp Faktör H Faktör I Anaflatoksin inhibitörü (karboksipeptaz B) S proteini (vitronektin)
 Pozitif regülasyon yapanlar. Properdin. Negatif regülasyon yapanlar. C I inhibitörü (CI INH) C4bp. Faktör H. Faktör I. Anaflatoksin inhibitörü (karboksipeptaz B) S proteini (vitronektin)")
17
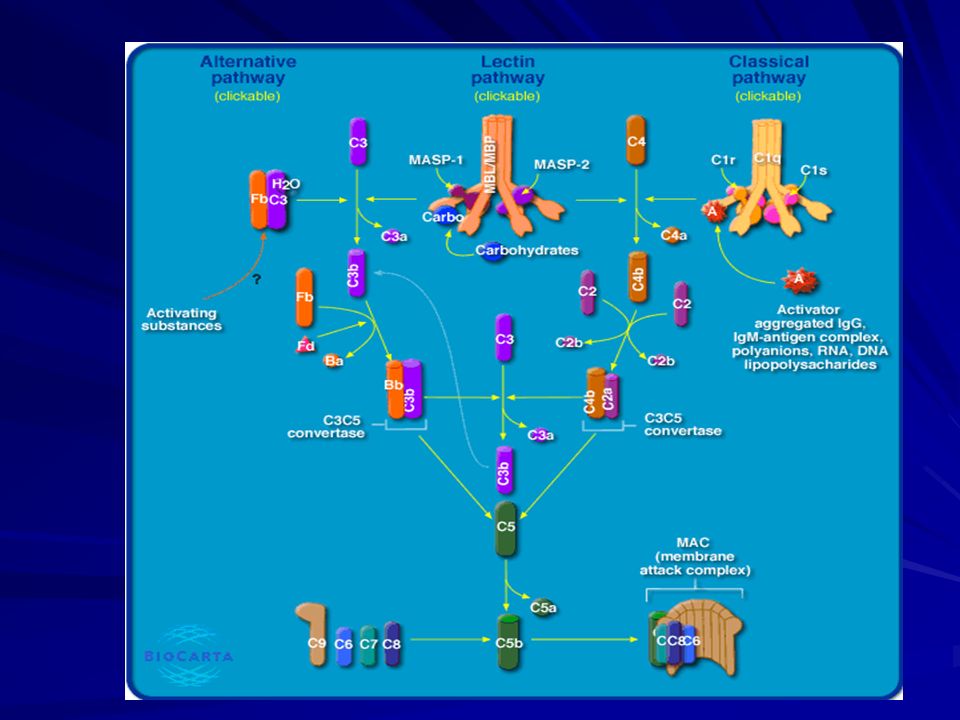
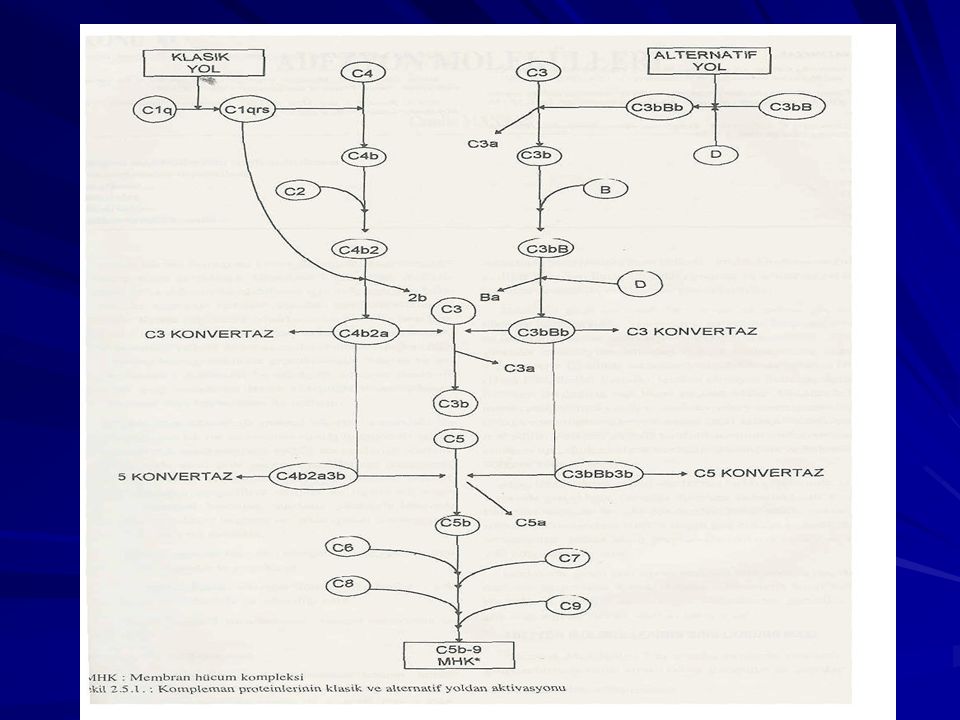
Kompleman Sistemi Klasik yol Ag-Ab kompleksleri
Alternatif yol - LPS Lektin yolu - Mannan-Bağlayan Protein Yolu -Mannoz, N-asetilglukozamin

18
Kompleman Sistemi Klasik yol C1, C2, C3, C4, C5, C6, C7, C8 ve C9
IgM ve Ig G ( IgG3, IgG1 ve çok az olarak IgG2 )
")
19
Klasik yol aktivasyonunun komponentleri
Ca++ C1r C1s C1q C4 C2 C3 Chelating agents dismantle the C1 complex and are anti-complementary. Heat destroys the C2 component. Sample for C measurement should be drawn in a green-top vial (no EDTA), must be kept cold and tested as soon as possible. C1 kompleks
, must be kept cold and tested as soon as possible. C1 kompleks.")
20
C1q ve C1’in Yapısı

21
Klasik yol C1 proteini, C1q, C1r, C1s
C1q parçası 6 polipeptid zincirinden oluşur. Karboksi uçları bir baş şeklinde globüler yapıdadır. Globüler baş kısımlarında 2 adet C1r ve 2 adet C1s molekülleri bulunur.

22
Klasik yol C1 ile bağlantı halinde olan C1r ve C1s C1r2s2 dimerik çiftlerini oluşturur ve her dimerik çift C1q’ yu bağlar C1qr2s2

23
Ag-Ab kompleksleri ile C1 aktivasyonu
C1q polipeptidleri globüler baş kısımlarıyla antikor molekülünün Fc kısmına bağlanır. IgG molekülünde CH2, IgM molekülünde CH3 domainleri üzerindedir. Bir antikor molekülünün Fc kısmında 1 adet C1q bağlanma yeri vardır. Antikor molekülü IgG veya IgM serumda serbest haldeyken C proteinleriyle bağlanmaz, antikor +antijen, kompleks oluşturduğunda, C bağlama yeri C1q için uygun hale gelir.

24
Klasik yol C1q molekülünün aktivasyon kazanabilmesi için, en az iki polipeptid başının antikorun Fc kısmına bağlanması gerekir. Her Ig molekülünde 1 adet C1q bağlanma yeri olduğu için, C1q molekülünün en az 2 Ig G molekülüne bağlanması gerekir. IgG molekülleri bakteri yüzeylerinde C1q’yu aktive edebilir. IgM molekülü , pentamer yapıdadır, bir tek IgM molekülü C1q proteinin aktivasyonu için yeterlidir

25
Klasik yol IgM molekülü IgG’ye oranla daha iyi kompleman bağlar.
IgG alt sınıflarından IgG3, IgG1 ve IgG2 IgG4, IgA, IgD, IgE komplemanı bağlamazlar.

26
C1qrs IgG’nin Fc Bölgesine Bağlanır

27
C1 parçasının en son C1s kısmının aktivasyonuyla, kompleman sisteminin ikinci molekülü C4 proteini etkilenir ve parçalanır. C C4a ve C4b C2 proteini serumda bulunan çözünür üçüncü parçadır. Hücre veya bakteri yüzeyinde bağlı halde bulunan C4b molekülüne, Mg varlığında bağlanır. C C2a ve C2b C4b2a klasik yol C3 konvertaz’ıdır. C4b2a C C3a ve C3b (metastable C3b) C4b2a + 3b C4b2a3b (klasik yol C5 konvertazı) C C5a ve C5b
 C4b2a + 3b C4b2a3b (klasik yol C5 konvertazı) C5 C5a ve C5b.")
28
Klasik yol

29
Klasik yol

30
Klasik Yol C3-konvertazın oluşumu
C4a b Ca++ C1r C1s C1q C4

31
Klasik Yol C3-konvertazın oluşumu
C2b a C4a Ca++ C1r C1s C1q C4b2a C3 konvertaz’ dır Mg++ C4b

32
Klasik Yol C5-konvertazın oluşumu
C2b C4a Ca++ C1r C1s C1q C3a b C4b2a3b C5 konvertaz; Membran Kompleksinin öncüsüdür Mg++ C4b C3 C2 a

33
Klasik yol

34
Alternatif Aktivasyon Yolu
C142 kompleksi bypas edilir. C3-C9 Klasik yolda rol oynamayan faktör B, faktör D ve properdin yer alır.

35
Alternatif Aktivasyon Yolu
Özgül antijen-antikor birleşmesi olmadan meydana gelir. Aktivasyon C3 proteinin parçalanmasıyla başlar. C3 proteinin parçalanması için C3 konvertaz molekülünün oluşması gerekir. C3 konvertaz molekülünün oluşması, iki molekülden başlar. C3b ve C3(H2o) veya C3i C3b molekülü klasik yolda oluşur C3 (H2o) , C3 molekülünde bulunan thioester bağının su molekülüyle hidrolize olmasıyla oluşur.
 veya C3i. C3b molekülü klasik yolda oluşur. C3 (H2o) , C3 molekülünde bulunan thioester bağının su molekülüyle hidrolize olmasıyla oluşur.")
36
Alternatif Aktivasyon Yolu
Faktör B C3b veya C3i ile birleşir (Faktör D proteazın etkisine duyarlı hale gelir). Faktör D Faktör B Ba ve Bb serbest C3bBb veya C3(H2o)Bb, C3iBb (C3 konvertaz) C3bBb kompleksi stabil değildir (yarı ömrü 2 dk) Properdin ile birleşerek daha dayanıklı hale gelir.
. Faktör D. Faktör B Ba ve Bb. serbest C3bBb veya C3(H2o)Bb, C3iBb. (C3 konvertaz) C3bBb kompleksi stabil değildir (yarı ömrü 2 dk) Properdin ile birleşerek daha dayanıklı hale gelir.")
37
Alternatif Yolda C3 Konvertaz Oluşumu

38
Alternatif Yolda C3 Konvertaz Oluşumu

39
Alternatif Yol Bileşenleri
D C3 B P

40
C3 - aktivasyonunun kendiliğinden oluşumu
C3 konvertaz oluşumu D b i H2O B C3a b C3 C3 C3iBb kompleksi çok kısa bir yaşam süresine sahiptir

41
C3-aktivasyonunun amplifikasyon kolu
Kendiliğinden oluşan C3b parçalanmaz b D B C3a b C3b C3

42
C3-aktivasyonunun amplifikasyon kolu
b D B C3a b C3b C3 Bb C3b C3a

43
C3-aktivasyonunun amplifikasyon kolu
b D Bb B b C3a C3b C3b C3 Bb C3b C3a C3a

44
C3-aktivasyonunun amplifikasyon kolu
Bb Bb C3b C3b C3b Bb C3b C3a C3a C3a

45
C3-aktivasyonunun amplifikasyon kolu
Bb Bb C3b C3b Bb C3b C3a C3a C3a

46
C3b stabilizasyonu ve C5 aktivasyonu
C3b bir aktivatör (koruyucu) membran bulur Bu alternatif yolun stabil C5 konvertazıdır D b P B C3b C3
 membran bulur. Bu alternatif yolun stabil C5 konvertazıdır. D. b. P. B. C3b. C3.")
47
C3’ün başlattığı Konvertaz

48
Alternatif Yol Aktivasyonu

49
Alternatif Yol Aktivasyonu

50
LEKTİN YOLU Lektinler karbonhidratları bağlayan proteinlerdir.
Mannoz bağlayan protein (MBP) bakterinin terminal grubundaki mannozu bağlayabilir. Klasik yol bu şekilde Antikor bağımsız şekilde de aktive olabilir. Lektine dayalı kompleman sistemi “mannoz-bağlayan protein” (MBP) ile başlar. MBP, MBP-associated serine protease (MASP) ile etkileşebilir. MASP C1q[r2s2], olan C3 konvertaz gibi etki yapar.
 bakterinin terminal grubundaki mannozu bağlayabilir. Klasik yol bu şekilde Antikor bağımsız şekilde de aktive olabilir. Lektine dayalı kompleman sistemi mannoz-bağlayan protein (MBP) ile başlar. MBP, MBP-associated serine protease (MASP) ile etkileşebilir. MASP C1q[r2s2], olan C3 konvertaz gibi etki yapar.")
51
Mannoz bağlayan lektin yolunun komponentleri
C4 MASP2 C2 MASP1 MBL

52
Mannoz bağlayan lektin yolu
C4b C4a C2b C2a C4b2a, C3 konvertaz; C5 konvertaz oluşumunu sağlar C4b C4 C2a C2 MASP2 MASP1 MBL

53
Lektin Yolu

54
Membran Atak Kompleksi
C4b2a3b ve C3bBb3b kompleksi C5 konvertaz etkisi gösterir. C C5a veC5b C6, C7,C8,C9 moleküllerinde proteolitik parçalanma olmadan bağlanma ve polimerizasyon olur. C5bC6 kompleksi hidrofiliktir. C7 eklenir, fosfolipid bağlayıcı etki ortaya çıkar. Serbest C5b67 kompleksinin ömrü 0.1 saniyedir. Hücre zarına bağlı olarak ömrü uzar.

55
Membran Atak Kompleksi
C8 ve C9 ile bağlantı yapabilir.C7 molekülü amfifiliktir. Aynı molekülde bulunmaları polimerizasyona yol açar C5b6789 polimerizasyonu sonucunda hücre zarını delip geçen borucuklar oluşur.Çapı ˚A kadardır. Na ve su alışverişinin bozulması içteki yüksek ozmotik basınç hücreyi lizise götürür. C9 molekülü hücrede erime oluşturmasında en etkili olandır.

56
İki yolun C5-konvertazı
(Klasik ve lektin yolunun) C5-konvertaz (Alternatif yolun) Bb C4b C2a C3b C3b C3b
 C5-konvertaz. (Alternatif yolun) Bb. C4b. C2a. C3b. C3b. C3b.")
57
Membran Atak Kompleksi Oluşumu

58
Klasik yol

59
Kompleman Sistemi: Terminal Yol

60
Litik Yol Bileşenleri C7 C6 C5 C8 C 9

61
Litik yol C5-aktivasyonu
b C5a C5 C3b C4b C2 a

62
C5 konvertaz oluşumu Litik yol aktivasyonuna öncülük eder

63
Litik yol Litik komplekslerin toplanması
C6 C5 b C7

64
Litik yol Litik kompleksin hücre membranına girişi
9 C 9 C 9 C 9 C 9 C 9 C 9 C 9 C 9

65
Terminal Kompleman Bileşenleri
Protein Aktif bileşen Fonksiyon C5 C5a İnflamasyonun küçük medyatörüdür. C5b MAK’ ni başlatır. C6 C5b’ yi bağlar, C7 için akseptördür. C7 C5b, C6 bağlar, amfifilik kompleks lipid bariyer içine girer. C8 C5b,6,7 bağlanır,C9 polimerizasyonu başlatır. C9 C9n C5b,6,7,8 polimerize olur, membran kanalı ve lizis oluşur.

66
Kompleman Aktivasyon Yolları

68
Kompleman Proteinlerinin Biyolojik Fonksiyonları
Hücre lizisi Opsonizasyon ve fagositoz Lökositlerin inflamasyon ve aktivasyonunda kemotaktik etkileşim İmmün komplekslerin solubilizasyonu ve fagositik temizlenmesi: Spesifik antikor cevabının indüklenmesi

69
Kompleman Konak Savunma Fonksiyonları
Kemotaksis, inflamasyon Opsonizasyon, nötralizasyon, B hücre aktivasyonu Litik kompleks formasyonu

70
Hücre lizisi Yabancı mikroorganizmalar öldürülür
Humoral immün yanıt sonucu oluşan özgül antikorlar vardır. Komplemanın C1-9’a kadar aktivasyonuyla olur. MAK ile yüzey zarda delik oluşur ve lizis gelişir.

71
Fagositoz için mikroorganizmaların opsonizasyonu
Hücre yüzeyine bağlı olarak C3b ve iC3b molekülleri oluşur. CR1 kompleman reseptörü C3b’ ye, CR2 kompleman reseptörü iC3b’ ye bağlanır.

72
Kompleman Sistemi bakterileri opsonize ederek fagositozunu kolaylaştırır

73
Lökositlerin inflamasyon ve aktivasyonunda kemotaktik etkileşim
C3a, C4a, C5a parçaları anaflotoksindir C3a ve C4a reseptörleri, mast hücresi, bazofil, düz kas hücreleri ve lenfositlerde bulunurlar. C5a reseptörü de mast hücresi, bazofil, nötrofil, monosit, makrofaj ve endotel hücrelerinde vardır.

74
Lökositlerin inflamasyon ve aktivasyonunda kemotaktik etkileşim
C3a/C5a’nın Biyolojik Fonksiyonları Anafilotoksik ve kemotaktik moleküllerdir. Mast hücrelerinin, bazofillerin ve eosinofillerin toksik degranülasyonuna sebep olurlar (histamin salınımı). Artan vasküler permeabiliteyi ve ödemi indükler. Sitokinlerin salınımını, adezyon molekülleri ve akut faz proteinlerinin ekspresyonlarını indükler. Respitatuvar yangıyı indükler/arttırır.
. Artan vasküler permeabiliteyi ve ödemi indükler. Sitokinlerin salınımını, adezyon molekülleri ve akut faz proteinlerinin ekspresyonlarını indükler. Respitatuvar yangıyı indükler/arttırır.")
75
C5a’ nın biyolojik etkileri

76
İmmün komplekslerin solubilizasyonu ve fagositik temizlenmesi
Eritrositlerde CR1 reseptörleri vardır. Bu reseptörlerin C3b’ye affiniteleri fazladır. Dolaşımdaki immün kompleksler komplemanı aktive ettiklerinde oluşan C3b antikor molekülüne kovalan bağla bağlanır. Böylece immün kompleksler eritrositlere bağlanır. Eritrositler antijen- antikor-kompleman kompleksini karaciğer ve dalağa fagositoz için toplar İmmün kompleksler dolaşımdan temizlenir.

77
Spesifik antikor cevabının indüklenmesi
B lenfositler ve antijen sunan hücrelerde antijen lokalizasyonunu arttırarak spesifik antikor cevabını indükler Kompleman sisteminin humoral bağışıklıktaki rolünün yanısıra, iltihap oluşumunda da önemli rolü vardır.

78
Kompleman sisteminin biyolojik fonksiyonları
Biyolojik etki Kompleman Oluş şekli Hücre lizisi C5b-C9 Membran atak kompleksiyle mikroorganizmaların ölümü Opsonizasyon fagositoz C3b, iC3b MO. ların yüzeyindeki C3b veya iC3b, nötrofil ve makrofajlardaki CR1(ve CR3, CR4) e bağlanır, fagositozu başlatır. İnflamasyon vasküler cevaplar C5a>C3a>C4a C5a, C3a, C4a mast hücrelerinden histamin salınmasına ve düz kas kasılmasına yol açarlar. Polimorfonüklear lökosit aktivasyonu C5a C5a nötrofiller için kemotatraktan olup, nötrofillerin oksidatif metabolizmasını aktive eder. İmmün komplekslerin uzaklaştırılması Klasik yol, C3b Ig molekülleri komplemanı aktive eder; immünkomplekse bağlı C3b, eritrositlerdeki CR1 ile bağlanır, eritrositler karaciğer ve dalaktan geçtikçe immünkompleksler dolaşımdan temizlenir.
 e bağlanır, fagositozu başlatır. İnflamasyon vasküler cevaplar. C5a>C3a>C4a. C5a, C3a, C4a mast hücrelerinden histamin salınmasına ve düz kas kasılmasına yol açarlar. Polimorfonüklear lökosit aktivasyonu. C5a. C5a nötrofiller için kemotatraktan olup, nötrofillerin oksidatif metabolizmasını aktive eder. İmmün komplekslerin uzaklaştırılması. Klasik yol, C3b. Ig molekülleri komplemanı aktive eder; immünkomplekse bağlı C3b, eritrositlerdeki CR1 ile bağlanır, eritrositler karaciğer ve dalaktan geçtikçe immünkompleksler dolaşımdan temizlenir.")
79
Kompleman Eksikliği Aktivasyon komponentleri – tekrarlayan bakteriyel enfeksiyonlar, SLE Terminal Komponentler – tekrarlayan bakteriyel enfeksiyonlar Reseptörler - SLE (CR1 ifadesinde eksiklik), hayatı tehtid eden bakteriyel enfeksiyonlar (CR3, CR4) Konjenital: C7, C8, C9’de eksiklik - “terminal compleman komponentleri”. Neisseria enfeksiyonlarında belirgin artan insidans. N. meningitidis: menenjit, bakteriemi N. gonorrhoeae: STD, bakteriemi Kazanılmış: kompleman sistemin aktivasyonu sırasında kompleman proteinlerinin aşırı kullanımı.
, hayatı tehtid eden bakteriyel enfeksiyonlar (CR3, CR4) Konjenital: C7, C8, C9’de eksiklik - terminal compleman komponentleri . Neisseria enfeksiyonlarında belirgin artan insidans. N. meningitidis: menenjit, bakteriemi. N. gonorrhoeae: STD, bakteriemi. Kazanılmış: kompleman sistemin aktivasyonu sırasında kompleman proteinlerinin aşırı kullanımı.")
80
Kompleman Sistemiyle İlgili Hastalıklar
Klasik yol C1q SLE, bakteriyel infeksiyonlar, hipopogamaglobülinemi,glomerülonefrit C1r Renal hastalık, SLE, romatoid hastalık,tekrarlayaninfeksiyonlar C1s SLE C4 SLE C2 Romatoid artrit, SLE, Glomerulonefrit, tekrarlayan infeksiyonlar C3 Piyojenik infeksiyonlar, Glomerulonefrit Alternatif Yol Properdin Piyojenik infeksiyonlar Faktör D Piyojenik infeksiyonlar Terminal komponentler C5 Dissemine Neisseria infeksiyonları C6 Dissemine Neisseria infeksiyonları C7 Dissemine Neisseria infeksiyonları, Glomerulonefrit, SLE C Dissemine Neisseria infeksiyonları, SLE C9 Dissemine Neisseria infeksiyonları

81
Kompleman Sistemiyle İlgili Hastalıklar
Kompleman Sisteminin düzenlenmesinde rol oynayan proteinlerin eksikliği C1INH Herediter anjionörotik ödem DAF Herediter anjionörotik ödem CD59 Paroksismal noktürnal hemoglobinüri Kompleman tip3 ve tip 4 reseptör lökosit adezyon eksikliği Herediter anjionörotik ödem

82
Kompleman Sistemi-Özet
Plazma proteinleri + lökositlerdeki kompleman reseptörleri + Düzenleyici proteinler Doğal immunitenin bir parçası Birçok eşleşmiş proteolitik reaksiyonla giderek artan etki elde edilir. Fonksiyonları İnflamatuar hücreleri uyarır (kemotaktik): C3a, C5a Patojenleri opsonize eder: C3b, C3bi Direkt mikrobisit: MAK
: C3a, C5a. Patojenleri opsonize eder: C3b, C3bi. Direkt mikrobisit: MAK.")







>")






>")





