Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Avian influenza virus and mutation
Köksal öksüz

2
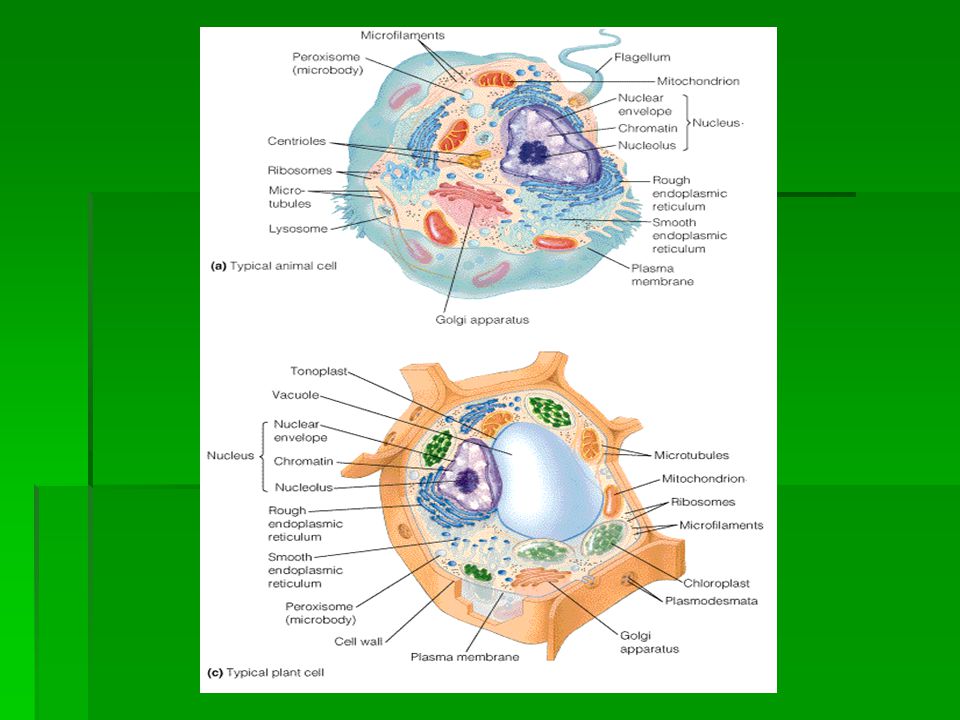
Bacterial cell Animal cell Plant cell

4
Gram-negative cell wall
Gram-positive cell wall Animal cell wall Plant cell wall

6
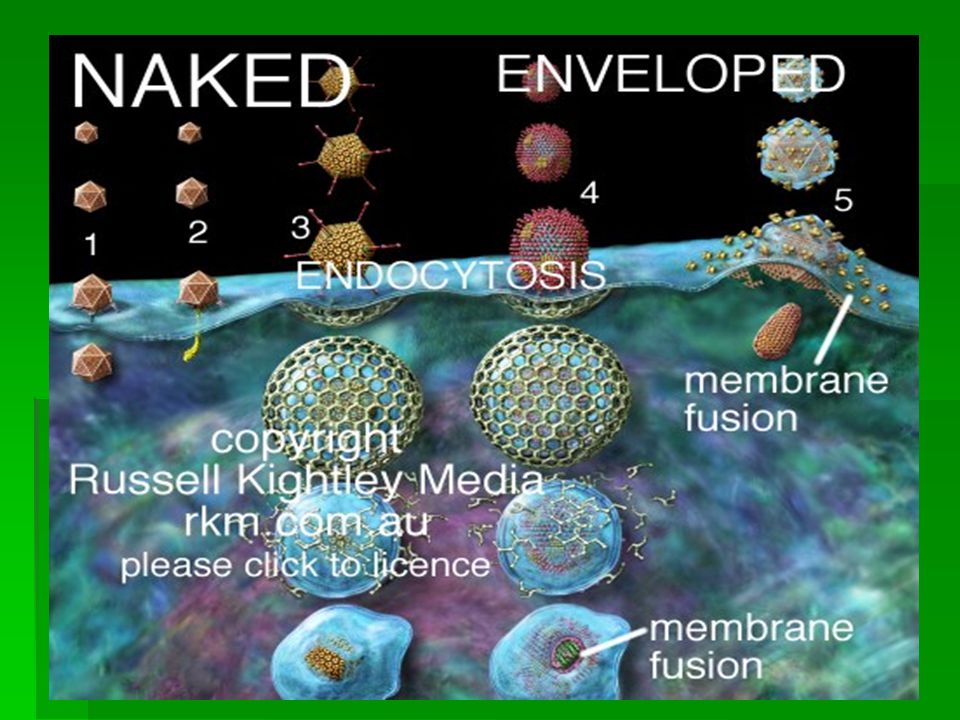
Graphic of Viral Entry into Animal Cells: virus attachment to cell surface (adsorption) and virus entry into cell. Picture shows translocation, pore formation, receptor mediated endocytosis using clathrin coated vesicles and membrane fusion. Some viruses can use more than one strategy. Other means are also employed. 1. NAKED VIRUS - TRANSLOCATION: particle crosses cell membrane intact (cf Principles of Molecular Virolgy, 3rd Edition, Alan J. Cann, Academic Press p 117)
")
7
2. NAKED VIRUS - GENOME INJECTION: virüs hücre yüzeyine tutunur ve genomunu plazma membranına açılmış bir por aracılığı ile bırakır (Bacteriophages, which attack bacterial cells, also inject their genomes and may use molecular "syringes" to do so, T4 phages injecting ).
..")
8
3. NAKED VIRUS - ENDOCYTOSIS: virüs hücre yüzey moleküllerine tutunur ve clathrin kaplı bir çukur oluşur ve onun içine düşer. Çukur tamamen virüsü kaplar ve sonunda clathrin kaplı bir kafes gibi kapanır. Bu molekül kafesi kısa bir süre sonra pervaneye benzeyen komponentlere ayrılır. Serbest kalan kese içerdiği viron’u endosoma taşır. Son aşamadai se viral komponentler açığa çıkar. Adenovirus

9
4. ENVELOPED VIRUS - ENDOCYTOSIS & MEMBRANE FUSION: virus enters cell by receptor mediated endocytosis. The cell membrane merges (fuses) with the endosome membrane and so the virus components are released. The virus shown here is an influenza virus, Influenza virus life cycle illustration
 with the endosome membrane and so the virus components are released. The virus shown here is an influenza virus, Influenza virus life cycle illustration.")
10
5. ENVELOPED VIRUS - MEMBRANE FUSION: virus enters the cell when its outer membrane fuses with the plasma membrane at the cell surface. The viral contents are then spilled into the cytoplasm of the cell. This example is HIV, which is unusual in having a conical core (most viral cores tend to be more spherical). HIV illustrations.
. HIV illustrations..")
11
next stage

12
Bacteriophages

13
T4 phages injecting

14
T4 phages injecting

15
adenovirus

16
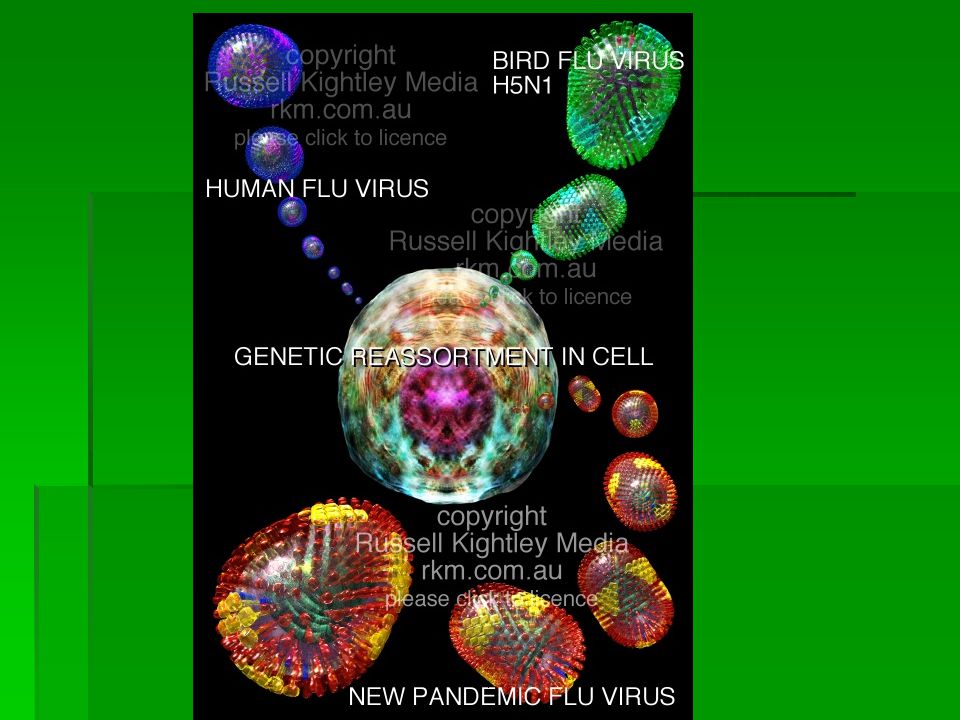
Genetic modification between human flu virus and avian influenza (bird flu) H5N1
 H5N1")
18
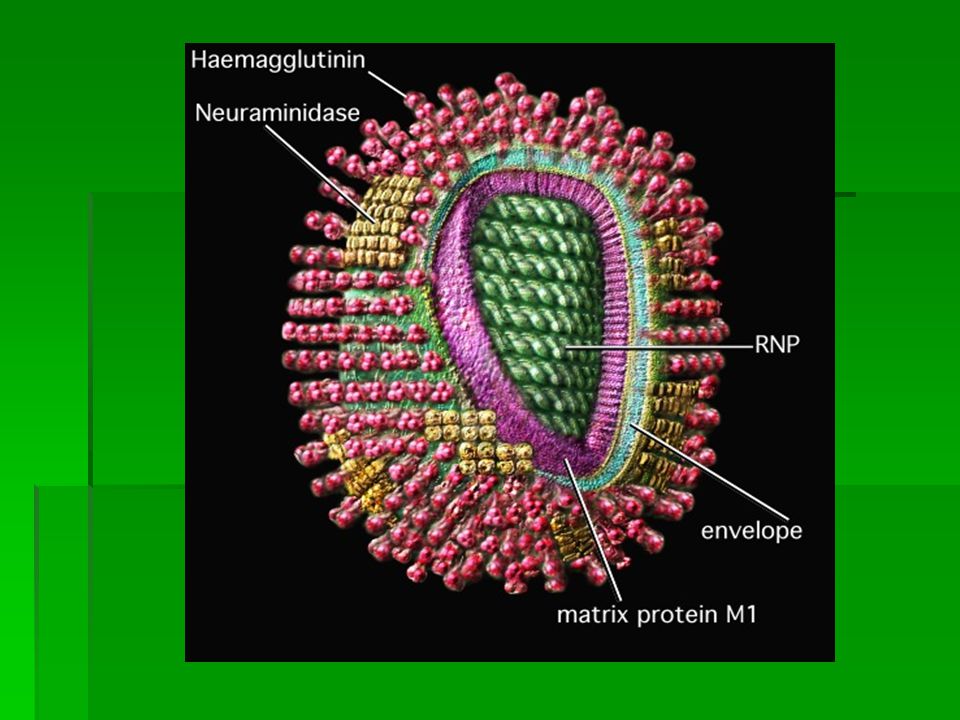
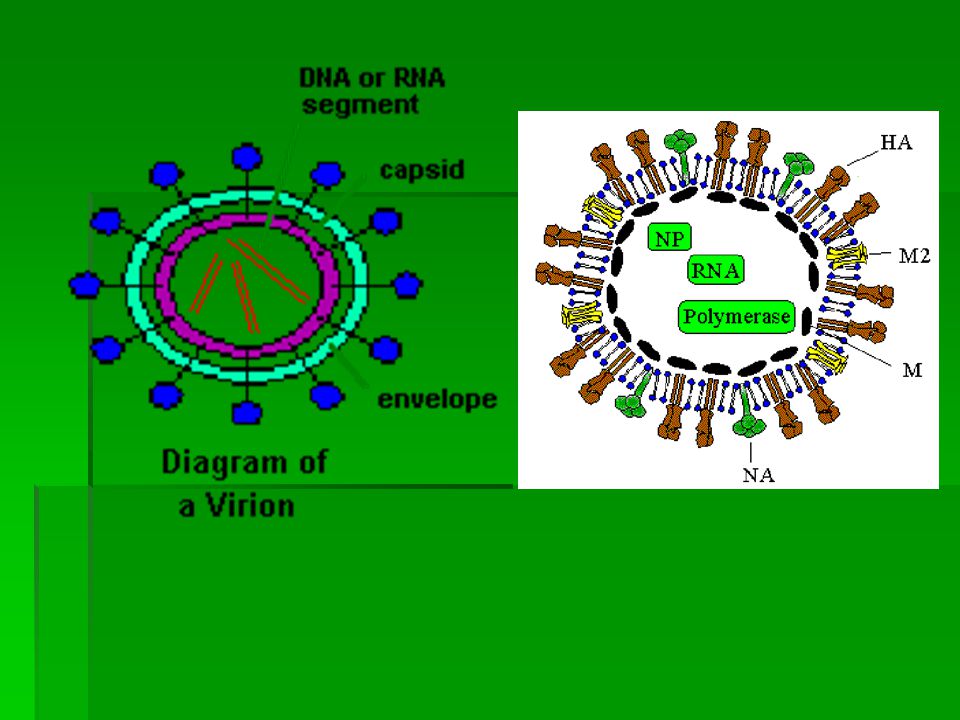
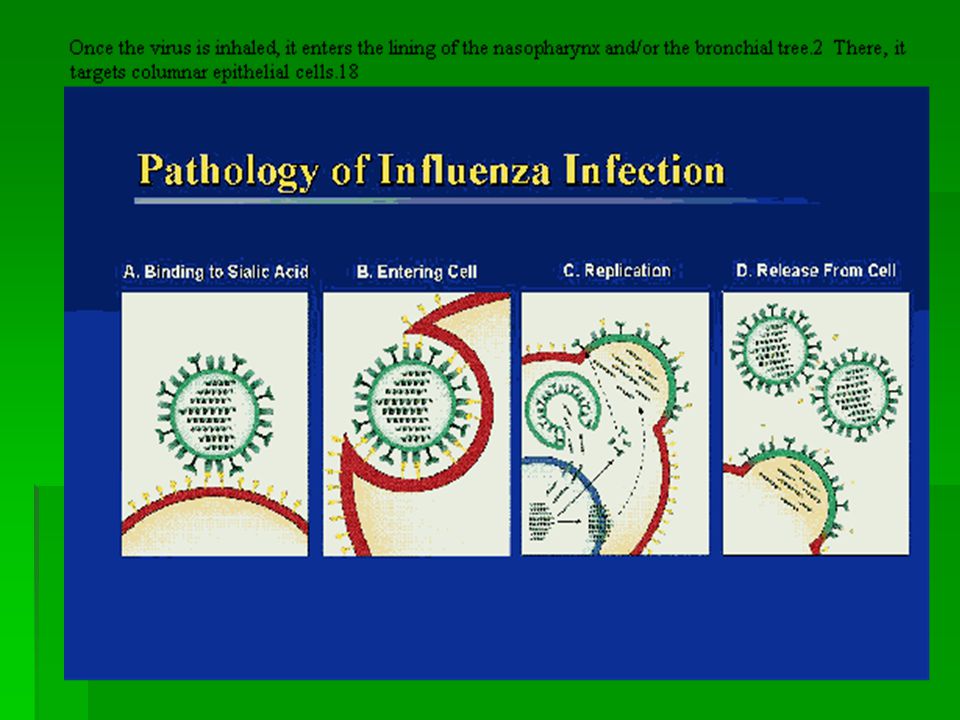
EXPLANATION OF PICTURE The green core ( RNP - ribonucleoprotein ) contains the genetic information of the virus wrapped up in protein. This combination of gentic material and protein is called the nucleocapsid. In influenza, the nucleocapsid is helical. The genetic information is stored as single stranded -ve sense RNA. The full complement of genetic information is called the genome and in influenza the genome is divided into eight segments. These segments are assumed to link together ( possibly in an ordered fashion ) to form a helix when the virus assembles at the cell surface. Overlying the nucleocapsid is a layer of matrix protein, M1, shown in purple. Overlying the matrix is the viral envelope (blue-green and edged in yellow) derived from the host cell membrane ( the nucleocapsid and the matrix proteins become wrapped in cell membrane as they bud from the infected cell ). The characteristic "spikes" of the influenza virus are haemagglutinin. They radiate all over the surface and are interspersed (in some types) by clusters of neuraminidase. These (HA and NA) molecules are thought to pass through the envelope and interact with the underlying matrix protein, M1
 to form a helix when the virus assembles at the cell surface. Overlying the nucleocapsid is a layer of matrix protein, M1, shown in purple. Overlying the matrix is the viral envelope (blue-green and edged in yellow) derived from the host cell membrane ( the nucleocapsid and the matrix proteins become wrapped in cell membrane as they bud from the infected cell ). The characteristic spikes of the influenza virus are haemagglutinin. They radiate all over the surface and are interspersed (in some types) by clusters of neuraminidase. These (HA and NA) molecules are thought to pass through the envelope and interact with the underlying matrix protein, M1.")
19
INFLUENZA VIRUS-TYPES
A- Birds, mammals (including humans, pigs, horses, sea mammals etc.) B- Humas C- Humans. Pigs
 B- Humas. C- Humans. Pigs.")
20
İnfluenza A ve B virüsleri iki yüzey glikoproteini taşır, Hemagglutinin (HA) ve Neuraminidase (NA). Her iki protein; aynı alıcı hücre molekülü olan sialik asidi tanımlar ve seçici olarak ona bağlanır. HA, virüs infeksiyonunu başlatmak için hedef hücrenin üzerindeki sialik asit içeren reseptöre bağlanır, oysa NA; hücresel reseptörlerden sialik asiti koparır ve hücre dışı geri kazandırıcıdır, yeni oluşan virüslerin serbest bırakılmasını kolaylaştırır ve komşu hücrelere enfeksiyonun sıçramasını teşvik eder. Bir çok araştırma glikoproteinlerin arasındaki reseptör bağlayıcı ve reseptör yıkıcı aktivitenin optimum etkileşiminin olmasının virus replikasyonu için gerekli olduğunu açıklamıştır.

21
Type A Influenza Surface Antigens Surface Antigen Subtype
1

23
H5N1 İnfluenza A H5N1, ilk kez güney afrikada 1961’de balıkçıllardan izole edilmiş olmakla birlikte, patojenitesi yüksek kuş gribi olarak daha önceden, ilk kez 1878’de İtalya’da tanımlanmıştır. Kuş gribi virüsünün doğal rezervuarı, yeşilbaş ördeklerdir. Ve enfeksiyona en en dayanıklı olan kuşlarda bunlardır. Virüsleri çok uzağa taşımalarına karşılık, yalnızca hafif hastalıklar geçirirler. Mutasyon sonucu oluşan H1N yıllarında milyon insanı öldürdü.

25
İnfluenza virüsü, solunum epitelia hücrelerine baştan başa bulaşmak ve geçişi kolaylaştırmak için birkaç mekanizmaya sahiptir A- virüsün yüzeyindeki HA proteinleri hücrenin yüzeyindeki tabakalaşmış sialik asite bağlanır B- bu komplexin oluşumu hücrenin tamamen virüsü kaplamasını tetikler C- viral RNA hücre nukleusuna girer ve viral replikasyon başlar D- yeni oluşmuş hücrenin yüzeyindeki NA; hücrenin yüzeyindeki sialik asit moleküllerini koparır, virüsün ortaya çıkmasına ve solunum organı’nın mukoz astarı boyunca yayılmasına olanak tanır. Viral replikasyon hücre ölüm prosesini başlatır, bu; virüsün ortaya çıkmasından ( replikasyondan sonra) birkaç saat sonra başlar.
 birkaç saat sonra başlar.")
29
BIRD FLU REASSORTMENT GRAPHIC: illustration above shows reassortment of viral RNA segments in a cell infected by two strains of influenza virus (human and bird flu) leading to a new and potentially dangerous strain that could spread easily from human to human and so trigger a deadly worldwide epidemic. Such genetic mixing might occur in pigs, since a pig might be infected by both strains and then pass the new virus on to humans. Alternatively, a person might become infected with bird flu and human flu and start an epidemic of the novel virus

30
END

32
Influenza virus life cycle illustration

33
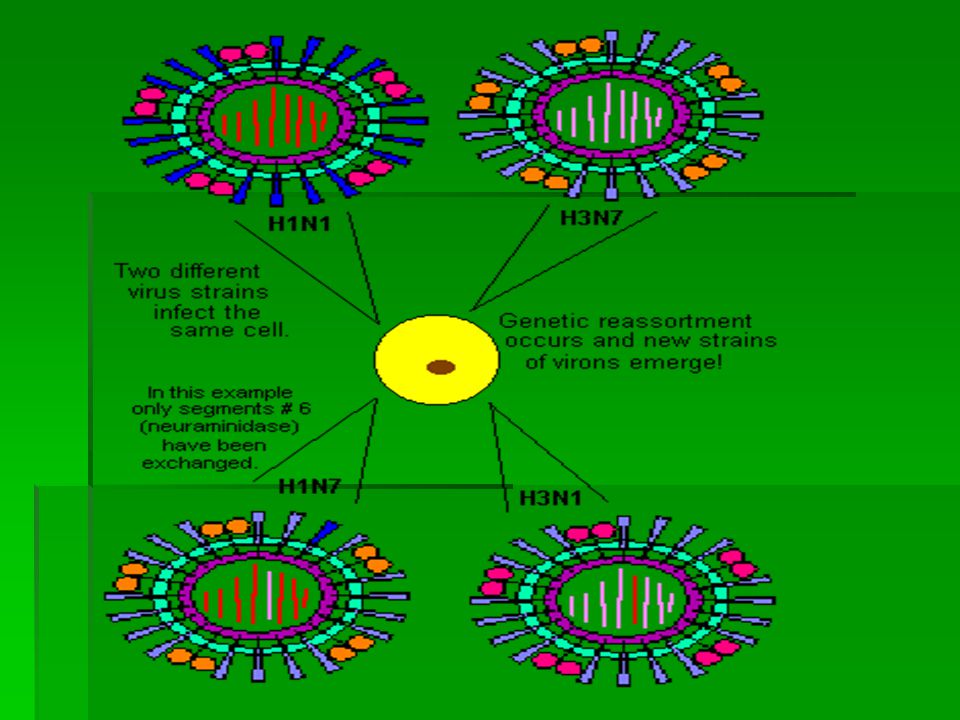
Influenza A virus has its RNA genome (genetic material) split into 8 segments. If two different viral types infect the same cell, then segments from both types can get jumbled together (they reassort) as the new virus particles are assembled. Consequently, new viral strains can emerge that contain a mixture of the parental genes. Image shows two different viral strains (BLUE genome at upper right and ORANGE genome at upper middle) infecting the same cell (at lower right). During replication, new viral particles may emerge that contain segments sourced form both the BLUE and the ORANGE strains. The new strain (BLUE & ORANGE STRIPED genome, shown at left) has the potential to spread rapidly.
 as the new virus particles are assembled. Consequently, new viral strains can emerge that contain a mixture of the parental genes. Image shows two different viral strains (BLUE genome at upper right and ORANGE genome at upper middle) infecting the same cell (at lower right). During replication, new viral particles may emerge that contain segments sourced form both the BLUE and the ORANGE strains. The new strain (BLUE & ORANGE STRIPED genome, shown at left) has the potential to spread rapidly..")
34
CELL ENTRY: At upper right, a BLUE influenza virus particle (representing an avian flu virus) is shown landing on the cell surface. The virus docks with cell membrane when the red spikes (haemagglutinin, shown in red) link to molecules on the cell surface. The cell surface folds inwards causing the virus particle to sink into the cell. The virus sinks deeper into the cell until it is completely wrapped up in cell membrane. The resulting membranous "bubble" (or vesicle) breaks free from the surface of the cell and transports its contained virus into the cell. The netlike structure beneath the docking virus and the cage-like structure around the resultant vesicle represent clathrin, a protein that forms an external scaffold that causes the cell membrane to invaginate and finally form the vesicle (this entry mechanism is called receptor mediated endocytosis please see our VIRAL ENTRY graphic). Further to the left an ORANGE influenza virus particle (representing a human flu virus) is shown landing on the cell surface.
 link to molecules on the cell surface. The cell surface folds inwards causing the virus particle to sink into the cell. The virus sinks deeper into the cell until it is completely wrapped up in cell membrane. The resulting membranous bubble (or vesicle) breaks free from the surface of the cell and transports its contained virus into the cell. The netlike structure beneath the docking virus and the cage-like structure around the resultant vesicle represent clathrin, a protein that forms an external scaffold that causes the cell membrane to invaginate and finally form the vesicle (this entry mechanism is called receptor mediated endocytosis please see our VIRAL ENTRY graphic). Further to the left an ORANGE influenza virus particle (representing a human flu virus) is shown landing on the cell surface..")
35
UNCOATING OF VIRUS AND RELEASE OF GENOME INTO CELL: The clathrin coat is then lost and the virus in its naked vesicle can be seen half out of frame at the right of the image. The engulfed virus then appears in an endosome (the large irregular yellow vesicle). It is more acidic in the endosome and this modifies the haemagglutinin spikes. The altered haemagglutinin draws the membranes of the virus and endosome together and they merge, creating a hole through which the viral contents are poured into the cytoplasm. These contents include the viral matrix protein (purple) and the nucleocapsid (BLUE segments). Some matrix protein is shown travelling to the nucleus. The nucleocapsid segments, which contain the viral genetic information, migrate to the nucleus. They move into the nucleus via nuclear pores (the flower like structures on the curved surface of the nucleus) and so deliver the viral genome to the nucleus (which contains the cell's own genetic material).
. It is more acidic in the endosome and this modifies the haemagglutinin spikes. The altered haemagglutinin draws the membranes of the virus and endosome together and they merge, creating a hole through which the viral contents are poured into the cytoplasm. These contents include the viral matrix protein (purple) and the nucleocapsid (BLUE segments). Some matrix protein is shown travelling to the nucleus. The nucleocapsid segments, which contain the viral genetic information, migrate to the nucleus. They move into the nucleus via nuclear pores (the flower like structures on the curved surface of the nucleus) and so deliver the viral genome to the nucleus (which contains the cell s own genetic material)..")
36
INSIDE THE NUCLEUS: In the nucleus, the viral genetic material (-ve sense RNA) produces viral messenger RNAs of various kinds (vmRNA) which travel out through the nuclear pores. (Messenger RNA, or mRNA, carries the genetic information that is used to direct protein maunfacture.) Some vmRNA directs the synthesis of nucleoprotein (green dots) that travel back into the nucleus. Other vmRNA directs the production of matrix protein (purple dots) shown emerging from a viral polyribosome (several ribosomes strung together along a length of viral mRNA) in the middle of the picture. Some matrix protein travels to the nucleus and some collects beneath the cell membrane. Other vmRNAs direct the production of external (transmembrane) viral proteins. The manufacture of such "external" proteins follows a different route. Production starts in the rough endoplasmic reticulum and progresses through the Golgi apparatus. The haemagglutinin (red) is shown progressing through the Golgi at lower left, finally being discharged onto the cell surface from a vesicle (the sphere containing red dots that is delivering its contents onto the cell surface through a hole). The neuraminidase (yellow) is shown (for clarity) going through the Golgi in parallel but above the haemagglutinin.
 Some vmRNA directs the synthesis of nucleoprotein (green dots) that travel back into the nucleus. Other vmRNA directs the production of matrix protein (purple dots) shown emerging from a viral polyribosome (several ribosomes strung together along a length of viral mRNA) in the middle of the picture. Some matrix protein travels to the nucleus and some collects beneath the cell membrane. Other vmRNAs direct the production of external (transmembrane) viral proteins. The manufacture of such external proteins follows a different route. Production starts in the rough endoplasmic reticulum and progresses through the Golgi apparatus. The haemagglutinin (red) is shown progressing through the Golgi at lower left, finally being discharged onto the cell surface from a vesicle (the sphere containing red dots that is delivering its contents onto the cell surface through a hole). The neuraminidase (yellow) is shown (for clarity) going through the Golgi in parallel but above the haemagglutinin..")
37
NEW VIRAL RNA SEGMENTS: In the nucleus, the viral -ve sense genome also produces +ve sense copies of itself. These are then used to create further copies of the viral genome. These new -ve sense viral genomic RNAs become associated with nucleoproteins and some matrix proteins that have migrated into the nucleus. Such newly formed nucleocapsids and their associated M proteins exit the nucleus via nuclear pores (BLUE and ORANGE segments can be seen streaming across the cell).
..")
38
VIRAL ASSEMBLY AT CELL SURFACE: Just beneath the cell surface, these individual BLUE and ORANGE ribonucleoprotein segments are shown associating together to form the helical nucleocapsid (the BLUE and ORANGE barrel-like structure). Around the new nucleocapsid, the matrix proteins are shown collected beneath the cell membrane (the haze of purple particles marked), while above the cell membrane, haemagglutinin and neuraminidase have coated the surface. With all these viral elements now in place, the newly forming virus particle (which contains segments derived from both the BLUE and ORANGE strains) can begin to take shape and to bud from the cell surface. The cell membrane that envelopes the emerging nucleocapsid and matrix protein becomes the viral envelope (complete with projecting spikes) and the virus particle is released. The new virus particle is now ready to infect another cell. Because it contains a new mix of genes, this reassortant can pose seroius dangers. This dramatic change in the genotype is called antigenic shift to distinguish it from the more minor changes that occur due to mutation or poor fidelity RNA copying, which are called antigenic drift.
, while above the cell membrane, haemagglutinin and neuraminidase have coated the surface. With all these viral elements now in place, the newly forming virus particle (which contains segments derived from both the BLUE and ORANGE strains) can begin to take shape and to bud from the cell surface. The cell membrane that envelopes the emerging nucleocapsid and matrix protein becomes the viral envelope (complete with projecting spikes) and the virus particle is released. The new virus particle is now ready to infect another cell. Because it contains a new mix of genes, this reassortant can pose seroius dangers. This dramatic change in the genotype is called antigenic shift to distinguish it from the more minor changes that occur due to mutation or poor fidelity RNA copying, which are called antigenic drift..")
39
Human Immunodeficiency Virus (HIV) particle (upper right) attaches to cell surface. The viral envelope merges with the cell plasma membrane (middle) releasing the matrix shell and core (purple bullet-shaped structure containing the viral genome) into the cell. The viral RNA (yellow string-like structure in core) is converted into viral DNA (red string-like structure in core) by the enzyme reverse transcriptase (green sphere). The viral genome is delivered to the nucleus (brownish sphere at bottom) and enters through a nuclear pore (flower-like opening). Once inside the nucleus, the viral DNA joins (integrates) with the host cell DNA. The viral DNA then directs the production of viral RNA which exits the nucleus through nuclear pores. Some viral RNA goes to form a new viral genome (yellow lump attached to underside of cell membrane at left) while other viral RNA directs the production of new viral proteins via ribosomes (3 brown spherical objects in middle of cell). The new viral components (genome + proteins) assemble at the cell surface (left) and a new virus particle buds from the cell (upper left).
 releasing the matrix shell and core (purple bullet-shaped structure containing the viral genome) into the cell. The viral RNA (yellow string-like structure in core) is converted into viral DNA (red string-like structure in core) by the enzyme reverse transcriptase (green sphere). The viral genome is delivered to the nucleus (brownish sphere at bottom) and enters through a nuclear pore (flower-like opening). Once inside the nucleus, the viral DNA joins (integrates) with the host cell DNA. The viral DNA then directs the production of viral RNA which exits the nucleus through nuclear pores. Some viral RNA goes to form a new viral genome (yellow lump attached to underside of cell membrane at left) while other viral RNA directs the production of new viral proteins via ribosomes (3 brown spherical objects in middle of cell). The new viral components (genome + proteins) assemble at the cell surface (left) and a new virus particle buds from the cell (upper left)..")
40
Human Immunodeficiency Virus (HIV or AIDS virus): HIV attacks a macrophage (top middle) and a Helper T Cell (lower left). New virus particles then bud from the macrophage. A B-lymphocyte (bottom right) gives rise to Plasma Cells (reddish cells on right) that produce antibodies (Y shaped molecules in red) that bind to HIV. A killer cell (bottom middle) will attack virus infected cells. The interaction of HIV and the immune system is very complex and varies over time.
 gives rise to Plasma Cells (reddish cells on right) that produce antibodies (Y shaped molecules in red) that bind to HIV. A killer cell (bottom middle) will attack virus infected cells. The interaction of HIV and the immune system is very complex and varies over time..")
41
Human Immunodeficiency Virus (HIV or AIDS virus): Human Immunodeficiency Virus: this illustration shows the external appearance of the virion. The virus is coated (enveloped) in host cell membrane, which is drawn as a bluish green semi transparent layer in which various membrane proteins can be seen floating. The viral knobs (golden projections at the viral surface) insert into the matrix. These knobs allow the virus to attach to cells.
 in host cell membrane, which is drawn as a bluish green semi transparent layer in which various membrane proteins can be seen floating. The viral knobs (golden projections at the viral surface) insert into the matrix. These knobs allow the virus to attach to cells..")
42
HIV cell entry and replication animation

43
back1

Benzer bir sunumlar















 TURKISH COMMERCIAL.>")





