Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Serbest Radikaller Ve Kanser ilişkisi

2
Serbest Radikal Nedir? Ortaklanmamış elektron taşıyan ve diğer biyolojik materyallerle reaksiyona girme eğilimi taşıyan atom veya moleküllere serbest radikal adı verilmektedir. İnsan vucudunda bütün hücrelere hiçbir zorlukla karşılaşmadan giren ve en çok kullanılma özelliğine sahip olan moleküler oksijen (O2) yapısı itibariyle radikal olmaya çok uygun olduğundan serbest radikal denilince aslında serbest oksijen radikalleri, daha genel bir tabirle reaktif oksijen türleri (ROS) akla gelmektedir.
 yapısı itibariyle radikal olmaya çok uygun olduğundan serbest radikal denilince aslında serbest oksijen radikalleri, daha genel bir tabirle reaktif oksijen türleri (ROS) akla gelmektedir.")
3
Serbest Radikal Kaynakları
Radyasyon Atom bombası denemeleri Nükleer atıklar Nükleer sızıntılar ( kazalar ) Nükleer tıp ( radyoloji, radyoterapi, radyoaktif araştırmalar ) Ultraviyole Işınları Güneş Işığı (UV ) Evlerde veya sanayide kullanılan elektronik cihazlar Hava Kirliliği Isınma veya ısıtma amaçlı kullanılan yakıtlar ( kalorifer dumanı... vs ) Eksoz borularının atıkları Sigara dumanı Enerji santralleri
 Nükleer tıp ( radyoloji, radyoterapi, radyoaktif araştırmalar ) Ultraviyole Işınları. Güneş Işığı (UV ) Evlerde veya sanayide kullanılan elektronik cihazlar. Hava Kirliliği. Isınma veya ısıtma amaçlı kullanılan yakıtlar ( kalorifer dumanı... vs ) Eksoz borularının atıkları. Sigara dumanı. Enerji santralleri.")
4
Serbest Radikallerin Meydana Geliş Şekilleri
1-) Kovalent bağlı normal bir molekülün, her bir parçasında ortak elektronlardan birinin kalarak homolitik bölünmesi. X : Y → X . + Y . 2-) Normal bir molekülden tek bir elektronun kaybı. X : Y → X - : Y + 3-) Normal bir moleküle tek bir elektronun eklenmesi. A + e- → A- .
 Kovalent bağlı normal bir molekülün, her bir parçasında ortak elektronlardan birinin kalarak homolitik bölünmesi. X : Y → X . + Y . 2-) Normal bir molekülden tek bir elektronun kaybı. X : Y → X - : Y + 3-) Normal bir moleküle tek bir elektronun eklenmesi. A + e- → A- .")
5
Serbest Radikal Reaksiyonları ve Serbest Radikal Türleri
Süperoksid anyon radikali ( O2.- ) Hidroperoksil radikali (HO2. ) Hidrojen peroksid ( H2O2 ) Hidroksil radikali (•OH ) Alkoksil radikali (RO•) Singlet oksijen ( O12 ) Nitrik oksit ( NO ) Peroksinitrit (ONOO-) Organik hidroperoksi (ROOH) Serbest radikaller genellikle birçok komşu molekülle reaksiyona girerler. Biyolojik sistemde devamlı olarak üretilirler. Serbest radikaller organik (Kinonlar) veya inorganik moleküller (süperoksit Anyon Radikali (O2)) halinde buluınurlar. Anyonik , Katyonik ve Nötral karakterde olabilirler. Ortaklanmamış elektronları sebebi ile yüksek reaksiyon kapasitesine sahiptirler. Bundan dolayı Diğer moleküllerle hızla kimyasal reaksiyona girmeye ve onların yapısını değiştirmeye eğilimlidirler. Aktif oksijen bileşiklerinin çoğu serbest radikaldir ve bunlara genellikle serbest oksijen radikalleri denilmektedir.
 Hidroperoksil radikali (HO2. ) Hidrojen peroksid ( H2O2 ) Hidroksil radikali (•OH ) Alkoksil radikali (RO•) Singlet oksijen ( O12 ) Nitrik oksit ( NO ) Peroksinitrit (ONOO-) Organik hidroperoksi (ROOH) Serbest radikaller genellikle birçok komşu molekülle reaksiyona girerler. Biyolojik sistemde devamlı olarak üretilirler. Serbest radikaller organik (Kinonlar) veya inorganik moleküller (süperoksit Anyon Radikali (O2)) halinde buluınurlar. Anyonik , Katyonik ve Nötral karakterde olabilirler. Ortaklanmamış elektronları sebebi ile yüksek reaksiyon kapasitesine sahiptirler. Bundan dolayı Diğer moleküllerle hızla kimyasal reaksiyona girmeye ve onların yapısını değiştirmeye eğilimlidirler. Aktif oksijen bileşiklerinin çoğu serbest radikaldir ve bunlara genellikle serbest oksijen radikalleri denilmektedir.")
6
Moleküler Oksijen (diradikal)
Moleküler oksiejen en dış yörüngesinde iki elektron bulundurur. Bu iki tek elektron birbirine paralel döner bu nedenle moleküler oksijen diradikaldir. Bu iki elektronun paralel sipinli olması nedeniyle moleküler oksijen oldukça stabildir. (diradikal)
")
7
Reaktif Oksijen Türleri (ROS)
Oksijen tek elektron alarak süperoksit radikaline dönüşüyor. Süperoksit radikali 2 proton alarak Hidrojen peroksit ‘e dönüşür. Hidrojen peroksit kendisi radikal değildir. Fentom ve Haber Weis tipi reaksiyonlarla demir ve bakır katalizi ile Hidroksil radikaline çevrilir.

8
1. Süperoksit Anyon Radikali (O2•-)
Tüm aerobik hücrelerde oksijen molekülünün bir elektron alarak indirgenmesi sonucu meydana gelir O2 + e- → O2.- O2.- + e- + 2H H2O2 Fe veya Cu iyonları H2O2 + O •OH + -OH +O2

9
2. Perhidroksi Radikali (HO.2)
3. Hidrojen Peroksit ( H2O2 ) O2 + 2e- + 2H H2O2 +O2 O2.- + e-+ 2H H2O2 +O2 2O H H2O2 +O2
 O2 + 2e- + 2H+ H2O2 +O2. O2.- + e-+ 2H+ H2O2 +O2. 2O H+ H2O2 +O2.")
10
4. Hidroksil Radikali ( •OH )
Hidroksil radikali (•OH), canlılarda başlıca şu mekanizmalarla oluşabilir: 1) Fenton reaksiyonu: Hidrojen peroksit Fe+2 ve diğer geçiş metalleri varlığında indirgenerek (•OH) radikali meydana gelir. O• Fe O2 + Fe+2 H2O2+ Fe •OH + -OH + Fe+3
, canlılarda başlıca şu mekanizmalarla oluşabilir: 1) Fenton reaksiyonu: Hidrojen peroksit Fe+2 ve diğer geçiş metalleri varlığında indirgenerek (•OH) radikali meydana gelir. O• -2 + Fe+3 O2 + Fe+2 H2O2+ Fe+2 •OH + -OH + Fe+3.")
11
2. Haber-Weiss Reaksiyonu: Hidrojen peroksit, süperoksit anyon radikali ile reaksiyona girerek hidroksil radikalini oluşturur. H2O2+ O• •OH+ -OH + O2 3. Suyun yüksek enerjili iyonize edici radyasyona maruz kalması sonucunda hidroksil radikali oluşur. χ veya γ ışını H-O-H • H + • OH

12
5. Singlet Oksijen ( 1O2 ) Oksijenin elektronlarından birinin dışarıdan enerji alması sonucunda kendi dönüş yönünün ters yönünde olan farklı bir orbitale yer değiştirmesi sonucu olur. O2- e O2

13
7. Peroksil Radikali (ROO • )
6. Nitrik Oksit (NO•) NO2• + O2• ONOO• ONOO • + H • OH + NO2 7. Peroksil Radikali (ROO • )
 NO2• + O2• ONOO• ONOO • + H+ • OH + NO2. 7. Peroksil Radikali (ROO • )")
14
Serbest Oksijen Radikallerinin Etkileri

15
Oksidatif DNA Hasarı ve Karsinogenesis

16
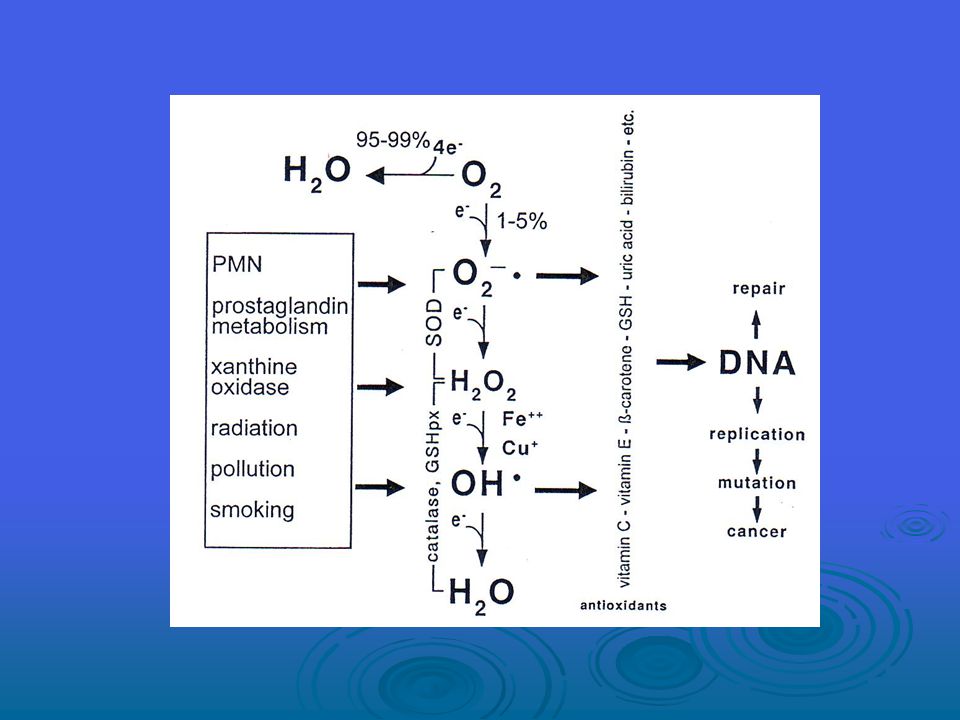
Reaktif oksijen türleri, çeşitli olaylar ve yolaklar aracılığıyla oluşur.
Bir insan hücresinin bir günde hidroksil radikaller ve diğer benzer reaktif türler tarafından yaklaşık 1.5x105 oksidatif darbeye maruz kaldığı tahmin edilmektedir. Hidroksil radikalin DNA molekülünün tüm bileşenleriyle reaksiyona girerek hem pürin hem de pirimidin bazlara ve aynı zamanda deoksiriboz omurgaya hasar verdiği bilinmektedir. Bu “oksidatif hasar” olaylarından doğan genetik materyalin kalıcı modifikasyonu, mutajenesis, karsinojenesis ve yaşlanmanın ilk adımını temsil eder.

17
Çeşitli kanser dokularında serbest radikallerin aracılık ettiği DNA hasarı oluşmuştur.
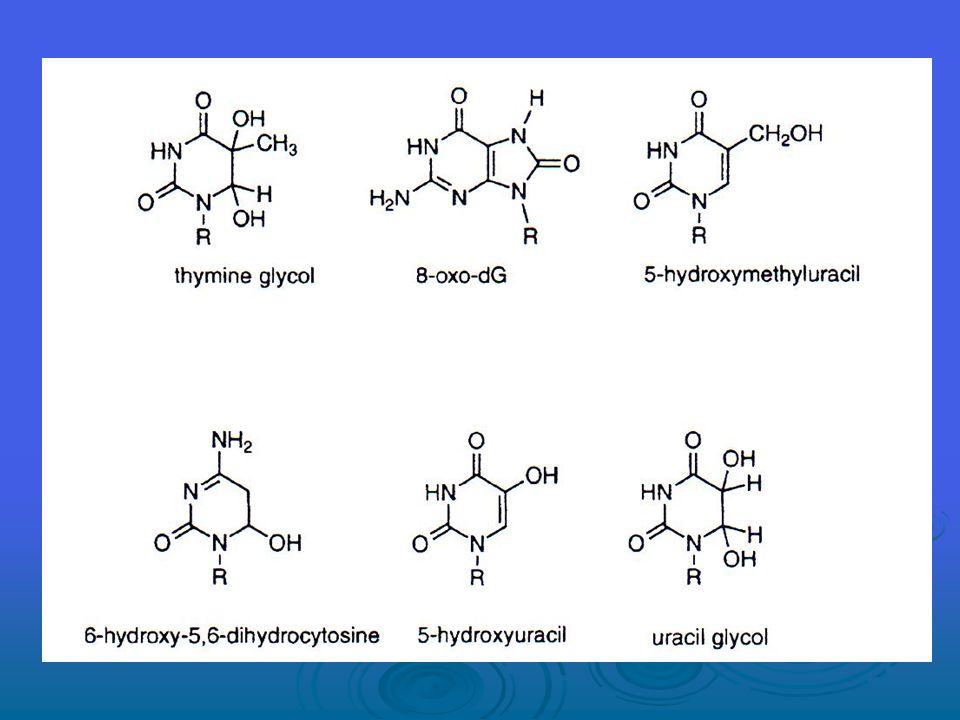
Bugüne kadar DNA oksidasyonundan doğan yüzden fazla ürün belirlenmiştir. ROS’un sebep olduğu DNA hasarı tek ya da çift zincirli DNA kırılmalarını, pürin, primidin veya deoksiriboz modifikasyonlarını ve DNA çapraz bağları içermektedir. DNA hasarı kesilme ya da transkripsiyon endüksiyonu, sinyal transdüksiyon yolaklarının endüksiyonu, kopyalama hataları ve genomik instabilite ile sonuçlanabilir ve bunların hepsi karsinojenesis ile ilişkilidir.

21
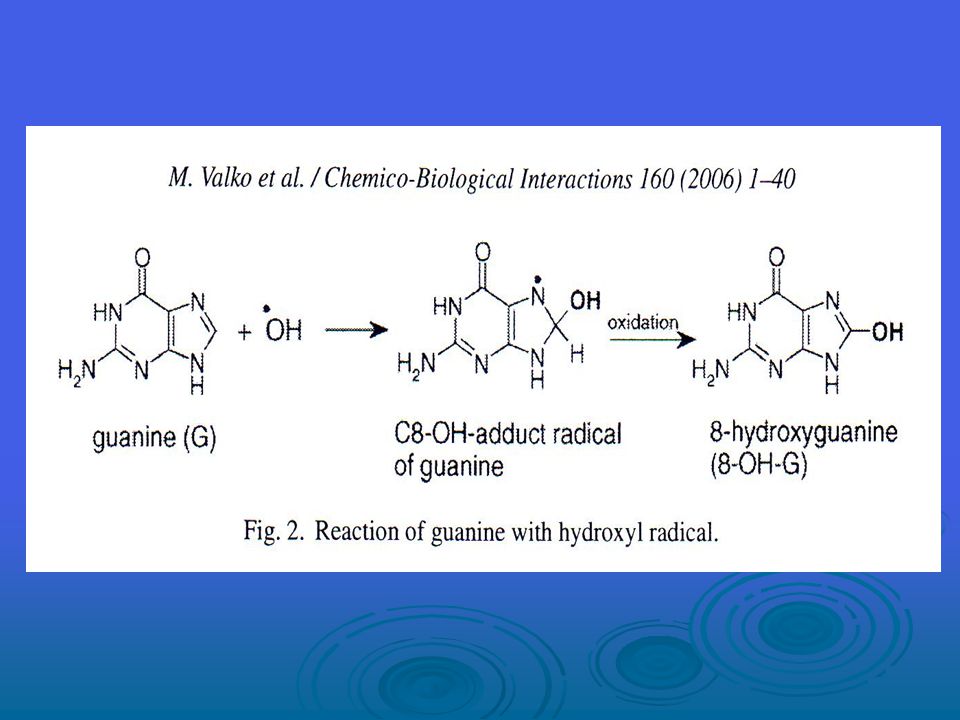
8-hidroksiguanin (8-OH-G)oluşumunun mekanizmalarını gösteren bir örnek verilmiştir.
İnsan idrarında 8-OH-G varlığı ilk olarak Ames ve meslektaşları tarafından rapor edilmiştir. Bu oksidize DNA prodüktü önemlidir çünkü hem oldukça kolay şekilde oluşur hem de mutajen ve karsinojendir. 8-hidroksiguaninin keto-enol tautomerizme uğradığını ve bu nedenle 8-OH-G’nin çoğu zaman 8-oxoguanin veya 8-oxo-G olarak adlandırıldığını görüyoruz; ancak 8-oxo-G ile 8-OH-G eşittir.

23
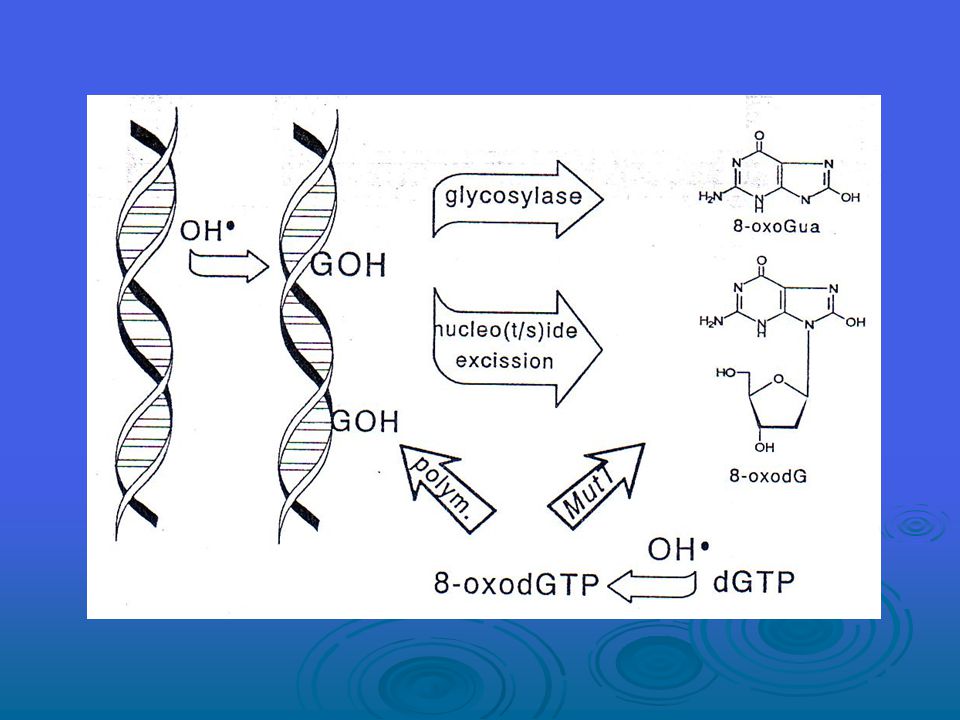
Hidroksil radikali, (3-10)x109M-1s-1lik bir alan içerisinde ikinci dizi oran sabitinde çift bağlı DNA bazları ekleyebilir ve timinin metil grubundan ve 2’ deoksiribozun beş karbon atomunun her birinden yaklaşık 2x109M-1s-1 lık bir oran sabitinde H-atomu abstract eder. DNA bazlarının OH radikalleri bir ekleme reaksiyonu yoluyla üretilirken, timinden türeyen radikal ve karbon merkezli şeker radikalleri abstraksiyon reaksiyonlarından doğar. Baz ve şeker radikallerinin bundan başka reaksiyonları; çeşitli modifiye bazlar ve şekerler, bazsız bölgeler, bağ kırılmaları ve DNA protein çapraz bağlarına yol açabilir.

25
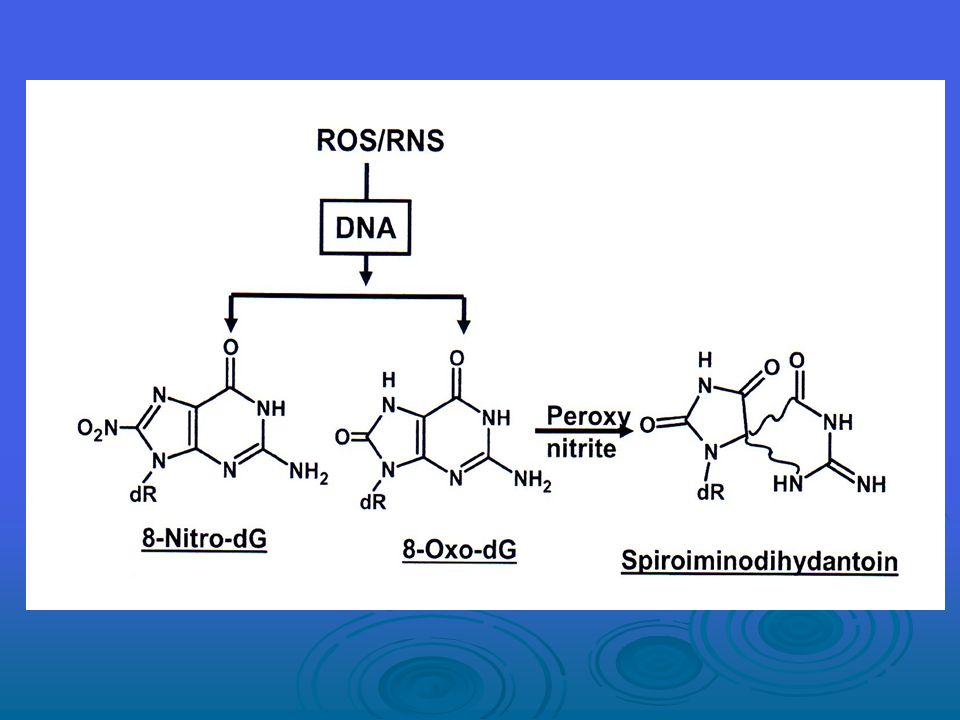
ROS’a ek olarak, peroksinitritler ve nitrojen oksitler gibi reaktif nitrojen türleri (RNS) de DNA hasarında belirtilmiştir. Peroksinitritlerin, guanin ile reaksiyona girdikten sonra 8-nitroguanin oluşturduğu ortaya konmuştur. Yapısı nedeniyle bu addüktün G:C T:A transversiyonlarına neden olma potansiyeli vardır. Bu lezyonun DNA’daki stabilitesi düşük olmasına karşın, bu nitrojen addüktü RNA’da stabildir. 8-nitroguanin ile karsinojenesis süreci arasındaki potansiyel bağlantı bilinmemektedir.

26
Neoplazide oksidatif nükleer DNA hasarının rolüne yönelik kapsamlı çalışmalara ek olarak kersinojenesis sürecinde mitokondrial oksidatif DNA hasarının ilgisi hakkında da kanıt bulunmaktadır. Mitokondriyal DNA’nın oksidasyona nükleer DNA’dan daha açık olduğu gerçeğini açıklamaktadır: (i) fizyolojik koşullar altında mitokondriya, tüketilen oksijenin ~%5’ini süperoksit anyona ve daha sonra hidrojen perokside dönüştürür; (ii) mitokondriyal DNA onarım kapasitesi sınırlıdır çünkü nükleotid eksizyon onarımından tamamen yoksundur; (iii) mitokondriyal DNA histonlar tarafından korunmaz.
 fizyolojik koşullar altında mitokondriya, tüketilen oksijenin ~%5’ini süperoksit anyona ve daha sonra hidrojen perokside dönüştürür; (ii) mitokondriyal DNA onarım kapasitesi sınırlıdır çünkü nükleotid eksizyon onarımından tamamen yoksundur; (iii) mitokondriyal DNA histonlar tarafından korunmaz.")
27
Hidrojen peroksit ve diğer reaktif oksijen türlerinin mitokondriyal biyojenesis, transkripsiyon ve mitokondriyal genomun replikasyonunda bulunan nükleer genlerin aktivasyonunda belirtilmiştir. Her ne kadar mutasyona uğramış mitokondriyal DNA’ya sahip tümör hücreleri bölgesi ve mitokondriyal DNA değişikliklerinin kanser sürecine ne derecede katkıda bulunduğu yeterli bir biçimde ortaya konmamış olsa da mitokondriyanın karsenojenesis ile bağlantısını destekleyen önemli miktarda bilgi bulunmaktadır. Bu bağlantı, mitokondriyal DNA parçacıklarının nükleer DNA’nın içine girmemiş olduğu gerçeğini destekler ve bu da onkojenlerin aktivasyonu için olası bir mekanizma öne sürer.

28
Sonuç olarak oksidatif genetik DNA modifikasyonunda gözlemlendiği gibi, oksidatif hasar ve mitokondriyal DNA’daki mutasyonun endüksiyonu, mitokondriyadan türemiş ROS, mitokondriyal genlerde mutasyonların endüksiyonu ve muhtemelen mitokondriyal genlerin nükleer DNA’ya girişi gibi karsinojenesis sürecinin birçok evresine dahil olabilir .

29
Yukarıda tanımlandığı gibi oksijen radikalleri mutajenesise yol açabilecek bazı DNA baz değişiklikleri başlatabilir. Ancak, DNA baz modifikasyonlarını onarabilen spesifik ve genel onarım mekanizmaları bulunmaktadır. Asıl ilgi çekici gerçek ise, onarım mekanizmalarının etkinliği, reaktif oksijen türlerine maruz kalmalarını takiben artabilir çünkü pek çok DNA onarım enzimlerinin ekspresyonu oksidatif stresten sonra upregulate olur.

30
Nükleer DNA’da oksidize olmuş bazların ~%90’ı tek nükleotid onarım mekanizmaları tarafından ve kalan %10 da uzun yama baz eksizyon onarımı ile onarıldığından tek nükleotid baz eksizyon onarımı, 8-OH-G’nin onarımı için birincil yoldur. 8-OH-G lezyonu için bir tamir mekanizmasının ilk kanıtı, röntgen ışınına tutulmuş bir farenin karaciğerinde gözlemlenmiştir; bu lezyonun seviyelerinin zamanla azaldığı görülmüştür. Bir onarım enzimi kısmi olarak E. coli’den arındırılmış ve daha sonra, klonlanan DNA onarım enzimi ile, yani daha önce E. coli’den izole edilmiş formamidopirimidin -DNA glikozilaz FPG proteini- ile aynı olduğu görülmüştür.

31
Nükleer DNA’daki DNA onarım mekanizmalarına ilişkin önemli bilgiler bulunmasına karşın, mitokondriyadaki onarım sistemlerine dair çok daha az şey bilinmektedir. Ancak nükleer DNA onarım mekanizmaları ile karşılaştırıldığında mitokondriyondaki DNA onarım kapasitesinin oldukça düşük olduğu görülmektedir. Hasar görmüş onarım kapasitesi mitokondriyal fonksiyon bozukluğuna ve dejeneratif hastalıkların baş göstermesine yol açabilir.

33
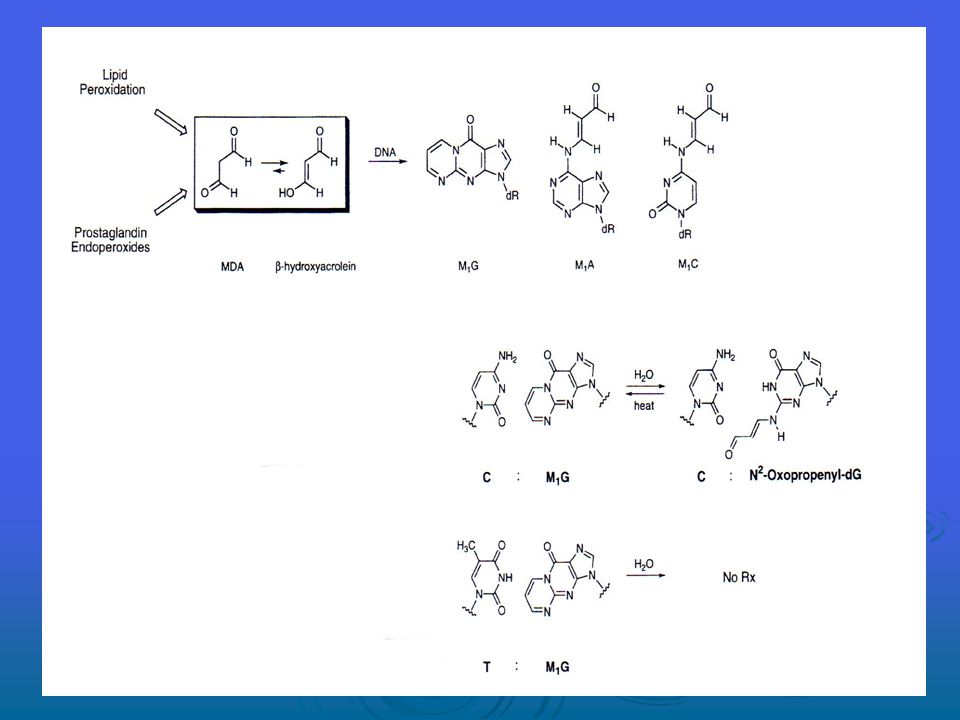
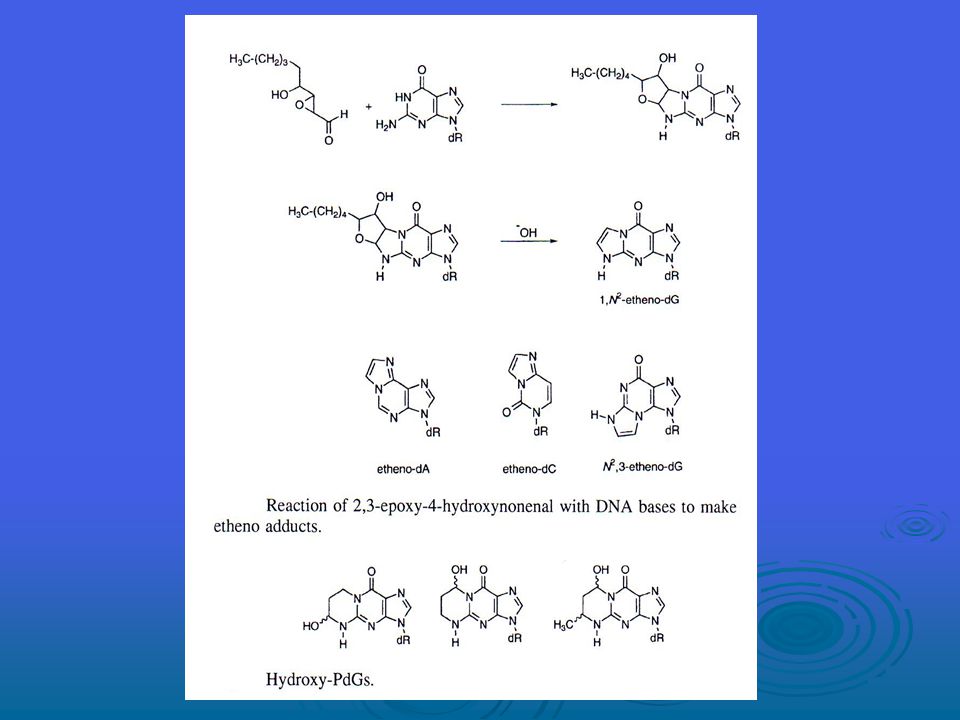
Lipit peroksidasyonun tüm süreci, üç evreden oluşur; başlangıç, yayılma ve sona erme -bu süreç Şekil ‘de sunulmaktadır. Oluştuktan sonra peroksil radikalleri (ROO), endoperoksitlere (malondialdehidin prekürsörü (öncüsü?)) bir siklizasyon reaksiyonu yoluyla yenden düzenlenebilir ve peroksidasyon işleminin son ürünü malondialdehid (MDA) olur. Lipid peroksidasyonun malondialdehid haricinde ana aldehid ürünü 4-hidroksi-2-nonenal’dir (HNE) MDA, bakteriyel hücre ve memeli hücrelerinde mutajen, farelerde ise karsinojendir. Hidroksinonenal, zayıf biçimde mutajendir ancak onun, lipid peroksidasyonun ana toksik ürünü olduğu görülmektedir. Buna ek olarak, HNE’nin sinyal transdüksiyon yolakları üzerinde güçlü etkileri vardır, bunun da hücrelerin fenotipik özellikleri üzerinde önemli bir etkisi vardır.
, endoperoksitlere (malondialdehidin prekürsörü (öncüsü )) bir siklizasyon reaksiyonu yoluyla yenden düzenlenebilir ve peroksidasyon işleminin son ürünü malondialdehid (MDA) olur. Lipid peroksidasyonun malondialdehid haricinde ana aldehid ürünü 4-hidroksi-2-nonenal’dir (HNE) MDA, bakteriyel hücre ve memeli hücrelerinde mutajen, farelerde ise karsinojendir. Hidroksinonenal, zayıf biçimde mutajendir ancak onun, lipid peroksidasyonun ana toksik ürünü olduğu görülmektedir. Buna ek olarak, HNE’nin sinyal transdüksiyon yolakları üzerinde güçlü etkileri vardır, bunun da hücrelerin fenotipik özellikleri üzerinde önemli bir etkisi vardır.")
35
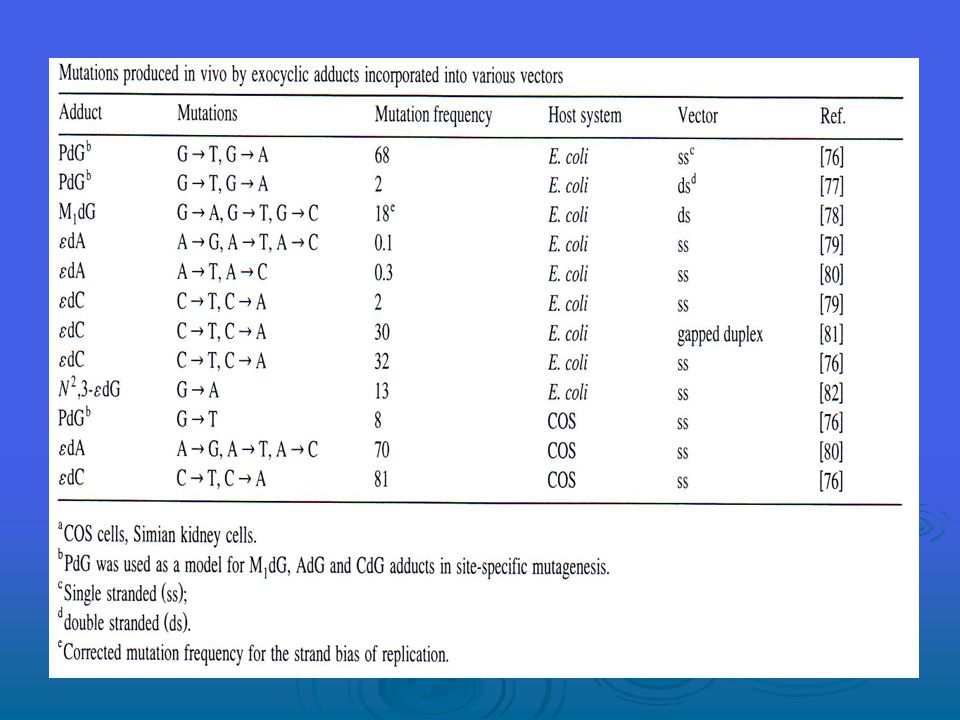
MDA, DNA bazları G, A ve C ile sırasıyla M1G, M1A ve M1C addüktlerini oluşturacak şekilde reaksiyona girebilir. Doku bakımından M1G addüktlerinin tespit sınırının altından 106 nükleosit başına 1.2 addükt gibi bir yüksekliğe (bu da yaklaşık hücre başına 6000 addükte tekabül eder) kadar çeşitlilik gösteren büyüklüklerde olduğu görülmüştür. M1G’nin E.coli’de mutajen olduğunu, T’ye transversiyon, A’ya da transisyon başlattığını onaylamıştır. Mutasyon sıklıkları, benzer sistemlerde 8-oxo-G için rapor edilenlerle benzerlik göstermektedir. M1G, hem bakteriyel hem de memeli nükleotid eksizyon onarım yolakları tarafından ve aynı zamanda E.coli’de yanlış eşleşme yoluyla onarılır. M1G’nin reaktif işlevselliği majör olukta mevcutken, N2-oxo-propenil-dG’nin reaktif işlevselliği DNA’nın minör oluğunda mevcuttur. DNA içerisinde M1G ve N2-oxo-propenal-dG’nin birbirine dönüşmesi DNA-DNA zincir arası çapraz bağlara veya DNA-protein çapraz bağlarına yol açabilir. Pek çok yazar tarafından hidroksil radikalinin proteinlerin oksidasyonundan sorumlu ana türleri temsil ettiği öne sürüldüyse de, kesin deneysel kanıt yoktur. Welch et al. tarafından gerçekleştirilen yeni bir çalışma, ferritin’in, sistein, tirosin ve diğer bazı rezidülerin oksidasyonunu içeren demir tarafından alana özgü modifikasyonunu ispatlamaktadır.
 kadar çeşitlilik gösteren büyüklüklerde olduğu görülmüştür. M1G’nin E.coli’de mutajen olduğunu, T’ye transversiyon, A’ya da transisyon başlattığını onaylamıştır. Mutasyon sıklıkları, benzer sistemlerde 8-oxo-G için rapor edilenlerle benzerlik göstermektedir. M1G, hem bakteriyel hem de memeli nükleotid eksizyon onarım yolakları tarafından ve aynı zamanda E.coli’de yanlış eşleşme yoluyla onarılır. M1G’nin reaktif işlevselliği majör olukta mevcutken, N2-oxo-propenil-dG’nin reaktif işlevselliği DNA’nın minör oluğunda mevcuttur. DNA içerisinde M1G ve N2-oxo-propenal-dG’nin birbirine dönüşmesi DNA-DNA zincir arası çapraz bağlara veya DNA-protein çapraz bağlarına yol açabilir. Pek çok yazar tarafından hidroksil radikalinin proteinlerin oksidasyonundan sorumlu ana türleri temsil ettiği öne sürüldüyse de, kesin deneysel kanıt yoktur. Welch et al. tarafından gerçekleştirilen yeni bir çalışma, ferritin’in, sistein, tirosin ve diğer bazı rezidülerin oksidasyonunu içeren demir tarafından alana özgü modifikasyonunu ispatlamaktadır.")
Benzer bir sunumlar















>")

 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")

>")

>")