Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
KARBONHİDRAT METABOLİZMASI
HİDROLİZ,SİNDİRİM SİSTEMİ,EMİLİM LAKTOZ İNTOLERANSI GLİKOLİZ VE DÜZENLENMESİ ANTİOKSİDAN MEKANİZMALAR GLUTATYONUN ROLÜ GLİOKSİLAZ PATHWAY’İ Dr. V.Kenan ÇELİK

2
Ösofagus:SİNDİRİM YOK MİDE: Asidik ortam amilazı inhibe eder.
SİNDİRİM SİSTEMİ AĞIZ: Tükrük amilazı ile nişastanın sindirimi Ösofagus:SİNDİRİM YOK MİDE: Asidik ortam amilazı inhibe eder. İNCE BARSAK: Pankreatik amilaz, tarafından nişastanın hidrolizi, İnce barsak mukoza hücreleri fırçamsı yüzeyi (Brush border): oligosakkaridazlar ve disakkaridazlar tarafından hidroliz. NİŞASTA LAKTOZ SUKROZ TREHALOZ FRUKTOZ K.Hidratların Erken sindirimi
: oligosakkaridazlar ve disakkaridazlar tarafından hidroliz. NİŞASTA. LAKTOZ. SUKROZ. TREHALOZ. FRUKTOZ. K.Hidratların. Erken. sindirimi.")
3
Tükrük veya Pankreatik
Tükrük bezlerinden salgılanan α-AMİLAZ, α-1-4 glikozidik bağlara etki eder. Tükrük veya Pankreatik AMİLAZ

4
K.HİDRATLARIN LÜMİNAL SİNDİRİMİ MALTOZ,LİMİT DEKTRİN + MONOSAKKARİTLER ,DİSAKKARİTLER

5
GLUKOAMİLAZ KOMPLEKSİ LAKTAZ, β-GALAKTOZİDAZ
OPTİMAL pH = 6 MALTOZ İZOMALTOZ SUKROZ GLUKOZ SUKRAZ-İZOMALTAZ KOMPLEKSİ GLUKOZ FRUKTOZ GLUKOAMİLAZ KOMPLEKSİ GLUKOZ OLİGOSAKKARİTLER GLUKOZ GALAKTOZ LAKTAZ, β-GALAKTOZİDAZ KOMPLEKSİ LAKTOZ TREHALAZ KOMPLEKSİ TREHALOZ GLUKOZ LİF

6
Laktoz glukoz + galaktoz
LAKTOZ İNTOLERANSI Laktoz glukoz + galaktoz laktaz gerekli İnce barsağın mikrovilli hücresinde oluşur Laktaz yetersizliği Gastro intestinal (GUT) bölgede laktoz birikimi su ile aşırı şişme; kramp oluşumu; diarrhea Intestinal bakteri: gaz oluşumu Genetic: %30 %80 Amerikan yerlilerinde %75 Afrikalı /Asyalılar 50% Latin 20% Kafkas GI hasar: inbar microvilli Hastalığı Aşırı ilaç Kötü beslenme
 bölgede laktoz birikimi. su ile aşırı şişme; kramp oluşumu; diarrhea. Intestinal bakteri: gaz oluşumu. Genetic: %30. %80 Amerikan yerlilerinde. %75 Afrikalı /Asyalılar. 50% Latin. 20% Kafkas. GI hasar: inbar microvilli. Hastalığı. Aşırı ilaç. Kötü beslenme.")
7
LAKTOZ İNTOLERANS TEDAVİSİ
Fermente edilmiş yiyecekler Peynir, yoğurt Önceden işlenmiş yiyecekler:Lactaid Süt ve süt ürünlerinden kaçınmak

8
İnce barsak İnce barsak 3 bölümden oluşur:: Duodenum, (Sindirim)
Jejunum, (besinlerin emilimi %95) Absorpsiyon milyonlarca hücrenin (villis) yer aldığı bölgede olur. İleum. (NaCl / sıvı emilimi, dehidratasyon)
 Absorpsiyon milyonlarca hücrenin (villis) yer aldığı bölgede olur. İleum. (NaCl / sıvı emilimi, dehidratasyon)")
9
İnce barsağın yapısı 1 4 5 2 3

10
Yüzey alanının emilimi artırıcı etkisi
Yapı Artan bağıl yüzeyalanı Yüzey alanı (m2) İncebarsağın Silindirik özel 4cm 3 1 280cm Kıvrımsal katlanma Villi Microvilli
 İncebarsağın. Silindirik özel. 4cm cm. Kıvrımsal. katlanma. Villi. Microvilli.")
11
Intestinal Absorption ve Secresyon
Villus hücreler Absorplama: NaCl Monosakkharidler Amino asitler Peptidler Yağlar Vitaminler Mineraller Su Crypt hücreler Cl & Su salınımı Villi Crypts

12
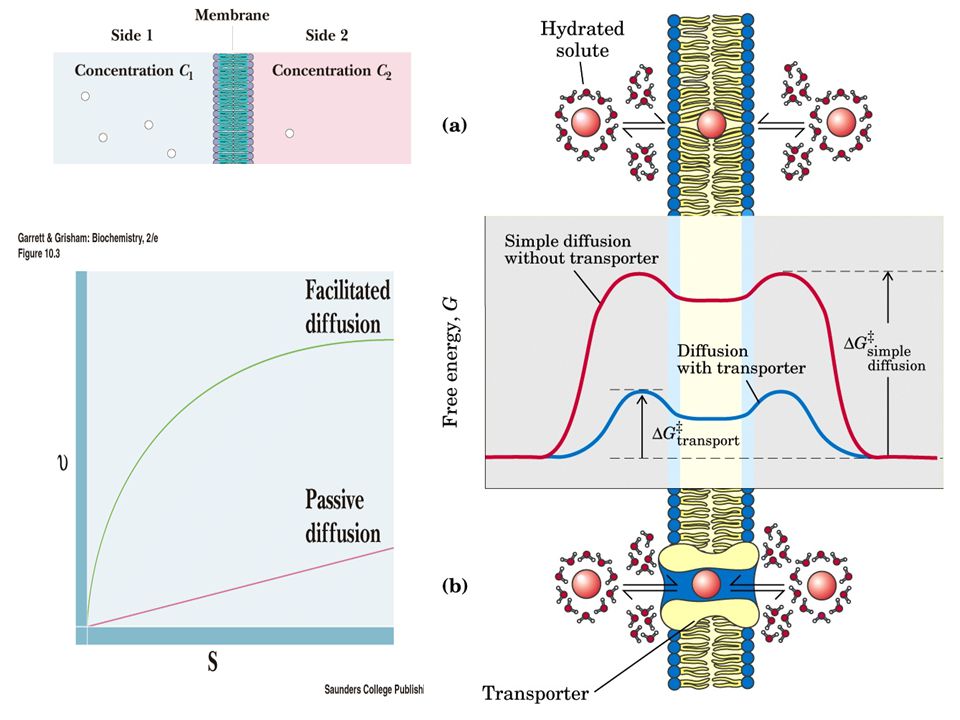
CHO Absorpsiyonu Glukoz ve Galaktoz Aktif emilim Enerji harcanır
Düşük konsantrasyondan yüksek konsantrasyona karşı taşınır Fruktoz Bir taşıyıcı kullanarak kolaylaşmış diffüzyonla Enerji harcanmaz.

14
KOLAYLAŞMIŞ DİFFÜZYON permeaz YADA transporter proteinler ARACILIĞI İLE OLUR.

15
Karbohidrat sindirimi ve emilimi
lumen de Nişaşta maltoz + maltrioz α-limit dextrinler Amylase brush-border da Maltoz/maltrioz glukoz Glukoamilaz (maltaz) Sukraz-izomaltaz α-limit dextrins glukoz Sukroz glukoz + fruktoz Laktoz glukoz + galaktoz Laktaz SGLT-1 Na+ coupled (Na/K ATPase for gradient) D-hexoses w/ pyranose ring GLUT-5 Fructose absorption Jejunum kolaylaşmış diffüzyon
 Sukraz-izomaltaz. α-limit dextrins glukoz. Sukroz glukoz + fruktoz. Laktoz glukoz + galaktoz. Laktaz. SGLT-1. Na+ coupled (Na/K ATPase for gradient) D-hexoses w/ pyranose ring. GLUT-5. Fructose absorption. Jejunum. kolaylaşmış diffüzyon.")
16
Na GLUKOZ LUMEN SGLT1 H2O Na GLUKOZ Na Na GLUT-2 GLUKOZ ~ kan K

17
FRUKTOZ LUMEN GLUT-5 FRUKTOZ GLUT-2 FRUKTOZ Kan

18
Kontraluminal membran
Fruktoz: glukoz, Glukoz Na Galaktoz LÜMEN İnce barsak Epitel hücresi Fırçamsı yüzey GLUT 5 SGLUT 1 Glukoz galaktoz Fruktoz glukoz ATP 3Na+ ADP + Pi Kontraluminal membran GLUT 2 3Na+ 2K+ Kapiler damar

19
AKTİF TRANSPORT: Primer transport Na/K ATPaz, H/K ATPaz, Ca ATPaz Sekonder transport: Glukoz, amino asit

20
Portal Ven Emilen monosakkaritlerin taşınımı Karaciğere teslim
K.ciğerde metabolizmanın gereksinimine göre yönlenme. (Glikoliz, depolama, sentez) Kan glukoz düzeyini idame ettirme.
 Kan glukoz düzeyini idame ettirme.")
21
Glukoz’un Hücrelere alınması
1.) Glukoz gradienti genellikle ESM>ISM mg/dl Normal < 45 mg/dl Hipoglisemi > mg/dl K.hidratca zengin bir öğün sonrası > mg/dl Kontrol edilemeyen diyabet 2.) Glukoz polar bir yapıdadır, bu nedenle HİDROFİLİK özellik gösterir. Özel glukoz taşıyıcıları (GLUT) aracılığı ile kolaylaşmış diffuzyonla taşınır. Bu derişim gradientine göre olur. (GLUT 1,2,3 ve 5) 3.) Düzenlenen glukoz alımı: İnsülin etkisi ile düzenlenen GLUT 4 kapıları aracılığıyla glukoz alınır.
 Glukoz gradienti genellikle ESM>ISM mg/dl Normal. < 45 mg/dl Hipoglisemi. > mg/dl K.hidratca zengin bir öğün sonrası. > mg/dl Kontrol edilemeyen diyabet. 2.) Glukoz polar bir yapıdadır, bu nedenle HİDROFİLİK özellik gösterir. Özel glukoz taşıyıcıları (GLUT) aracılığı ile kolaylaşmış diffuzyonla taşınır. Bu derişim gradientine göre olur. (GLUT 1,2,3 ve 5) 3.) Düzenlenen glukoz alımı: İnsülin etkisi ile düzenlenen GLUT 4 kapıları aracılığıyla glukoz alınır.")
22
GLUT 1,2,3,4, 5 TİP DOKU ÖZELLİKLER Nöron,eritrosit,beyin,
plasenta, böbrek. İnce barsak Km ~ 1 mM.Tetramer yapıda. 492 aa 664 aa, tetramer. Na ile glukoz taşınımı. GLUT 1 SGLUT 1 GLUT 2 Kc,Pankreasın β hücreleri, İn.bar.Böbrek,eritrosit , Km = mM. Hücre içi ve dışı glukoz derişimini dengeler. Glukoz taşıma potansiyeli yüksek, Afinitesi düşük. GLUT 3 Beyin,Böbrek, Plesanta GLUT 2 ye göre düşük Km (<1 mM), Hipoglisemi durumunda glukoz alımına öncelik oluşturur. GLUT 4 Kalp ve İskelet kası, Adipoz. Km (2.5-5 mM).İnsülin aracılığı ile glukoz alımı sağlanır.% kan glukoz değerini azaltır. GLUT 5 İn.barsak Km (~ 6 mM). Fruktoz alımını sağlar..
, Hipoglisemi durumunda glukoz alımına öncelik oluşturur. GLUT 4. Kalp ve İskelet kası, Adipoz. Km (2.5-5 mM).İnsülin aracılığı ile glukoz alımı sağlanır.% kan glukoz değerini azaltır. GLUT 5. İn.barsak. Km (~ 6 mM). Fruktoz alımını sağlar..")
23
GLUT 1,2,3 ve 5 1 2 3 4 5 6 7 8 9 10 11 12

24
Pankreas’ın β hücreleri tarafından salgılanan bir hormon.
İNSÜLİN Pankreas’ın β hücreleri tarafından salgılanan bir hormon. Başlıca rolü glukozun kan dolaşımından alınıp adipoz ve kas dokusu hücrelerine girişini sağlamak. Bir öğün sonrası artan kan glukozu beta hücrelerini stimule ederek kana İNSÜLİN salınımını sağlar. İnsülin hücre reseptörleri ile etkileşerek glukozun hücrelere girişine izin verir.

25
Sevgili arkadaşlar çalışın

26
GLUKOZ Stimulus–secretion coupling in the pancreatic b-cell
Translation INSULIN PDX1 NeuroD Pax6 E2A Transcription Splicing Processing Granule form. Secretion Glut-2 Glucose Glu-6P Glycolysis DAG + 1,4,5-Ins-P3 Depolarization K+ ATP Ca2+ sensitization and release from intracellular stores Exocytosis Glucokinase Ca2+ Voltage sens. Ca2+ Channel K+/ATP-Channel PLC PKC + Ca2+ Stimulus–secretion coupling in the pancreatic b-cell J. Jensen GLUKOZ

29
İnsülin Insülin Reseptör A zinciri 21AA B zinciri 30AA A Subunits
Ful aktif Tirozine Kinaz hücre içi enzimleri fosforile eder. (aktive/deaktive) A Subunits S S B Subunits Tirozin Kinaz (Otofosforilasyon)
 A Subunits. S. S. B Subunits. Tirozin Kinaz (Otofosforilasyon)")
30
GLUT 4 glu GLUT 4 GLUT4 GLUT4

31
Asetil CoA CO2 f GLUKOZ Piruvat Pentoz Fosfat a2 b c d g KARACİĞER Glikojen Glukoz-6-P Laktat h i j k e ' YAĞ

32
Asetil CoA CO2 f GLUKOZ Pirüvat Pentoz a4 b c d g KAS HÜCRESİ Glikojen G-6-P Laktat e h i ' LAKTAT KAN

33
BEYİN a1 GLUKOZ Pentoz b Glucose-6-P c d f Pirüvat Asetil CoA CO2 g

34
Asetil CoA f GLUKOZ Piruvat Pentoz a4 b c d ADİPOZ G-6-P YAĞ ASITLERİ j

35
a1 GLUKOZ PENTOZ b Glukoz-6P c d Pirüvat Laktat LAKTAT e
ERİTROSİT HÜCRESİ GLUT 2

36
KIRMIZI KAN HÜCRELERİNDE GLUKOZ METABOLİZMASI
70 kg olan bir kişi de 5 L kan vardır,~ 2 kg eritrosit taşır. Bu hücreler yegane enerji kaynağı olarak 20 g (0.1 mol) / gün glukoz tüketir.Toplam glukoz metabolizmasının % 10 . Kırmızı kan hücreleri diğer dokulara göre glukoz kullanım oranı oldukca yüksektir. (10g/kg doku/gün) Bu glukozun % 90 nı ANAEROBİK GLİKOLİZ yoluyla metabolize edilir. Günlük 0.2 mol LAKTAT (18 g/gün) oluşturulur. Glukoz kullanım oranının yüksek olmasına rağmen ATP sentezi diğer hücrelere göre daha düşük oranda (0.2 mol ATP/ gün)
 / gün glukoz tüketir.Toplam glukoz metabolizmasının % 10 . Kırmızı kan hücreleri diğer dokulara göre glukoz kullanım oranı oldukca yüksektir. (10g/kg doku/gün) Bu glukozun % 90 nı ANAEROBİK GLİKOLİZ yoluyla metabolize edilir. Günlük 0.2 mol LAKTAT (18 g/gün) oluşturulur. Glukoz kullanım oranının yüksek olmasına rağmen ATP sentezi diğer hücrelere göre daha düşük oranda (0.2 mol ATP/ gün)")
37
GLİKOLİZ - Biyokimyanın başlangıcında ilk keşfedilen metabolik yoldur. Karbohidrat metabolizmasında merkezi bir role sahiptir.Glikoliz yunanca gliko (tatlı) ve lizis (ayrışma) kelimelerinden oluşmuştur.6 C’lu hekzosların 3 ve 2 karbonlu (piruvat,etanol) bileşiklere ayrışmasıdır.Glikoliz AEROBİK ve ANAEROBİK koşullara göre farklılık göstermektedir. Glukozun aerobik koşullarda ki tam oksidasyonu (aerobik organizmalar) Glukozun anaerobik koşullarda ki metabolizması: Glukoz pirüvat CO2 + H2O ADP ATP Glikoliz TCA / oxidative fosforilasyon Glukoz pirüvat Laktat (. Memeli kas hücreleri, eritrosit) ADP ATP Glikoliz Etanol (maya)
 ve lizis (ayrışma) kelimelerinden oluşmuştur.6 C’lu hekzosların 3 ve 2 karbonlu (piruvat,etanol) bileşiklere ayrışmasıdır.Glikoliz AEROBİK ve ANAEROBİK koşullara göre farklılık göstermektedir. Glukozun aerobik koşullarda ki tam oksidasyonu (aerobik organizmalar) Glukozun anaerobik koşullarda ki metabolizması: Glukoz. pirüvat. CO2 + H2O. ADP. ATP. Glikoliz. TCA / oxidative. fosforilasyon. Glukoz. pirüvat. Laktat (. Memeli kas hücreleri, eritrosit) ADP. ATP. Glikoliz. Etanol (maya)")
38
Glikoliz hücre SİTOZOL’ün gerçekleşir.
1 Mol 6 6 2 Mol 2 Egzersiz yapan kaslarda,eritrositlerde

39
Evre 3: 3 C’lu bileşiklerin PİRUVATA oksitlenmesi ile ATP üretimi.
Glikoliz evreleri 1.Evre Evre 1: ATP gereksinimi vardır. Glukoz’ un fosforlanıp Fruktoz 1,6 bisfosfata (F-1,6 BP) dönüştürülmesi. Evre 2: F-1,6 BP’ dan 2 tane 3C’lu (GAL-3-P ve DHAP) bileşiklerin ayrılması. Evre 3: 3 C’lu bileşiklerin PİRUVATA oksitlenmesi ile ATP üretimi. 2.Evre 3.Evre
 dönüştürülmesi. Evre 2: F-1,6 BP’ dan 2 tane 3C’lu (GAL-3-P ve DHAP) bileşiklerin ayrılması. Evre 3: 3 C’lu bileşiklerin PİRUVATA oksitlenmesi ile ATP üretimi. 2.Evre. 3.Evre.")
40
Fosforilasyon KİNAZLAR arcılığı ile gerçekleşir
Fosforilasyon KİNAZLAR arcılığı ile gerçekleşir.Fosforlanan Glukoz’un kararlılığı azalır. Hekzokinaz Glukokinaz ΔG°’= kcal mol-1

41
Glukokinaz: Karaciğerde bulunur. Hekzokinaz:
hemen hemen tüm hücrelerde bulunur. Diğer mono sakkaritleride fosforile eder, fakat bunlara karşı afinitesi düşüktür.Glukoz için Km = 0.1 mM. Glukoz-6-fosfat tarafından inhibe edilir. Glukokinaz: Karaciğerde bulunur. Glukoza spesifiktir. glukoz-6-fosfat tarafından inhibe edilmez. Beslenme ile kanda yükselen glukoz tarafından indüklenir. Glikojen sentezi için G-6-P üretir.

42
Fosfogukoz izomeraz G°´ = 1.7 kJ/mole ΔG°’= .40 kcal mol-1 Aldo-keto dönüşümü Tersinir reaksiyon

43
Fosfofruktokinaz-1 İkinci ATP gereksinimi,Glikolitik yoldaki ikinci tersinmez reaksiyon ve oldukca ekzergoniktir. Glikoliz süresince enerji ve C gereksinimine yanıt oluşturmak üzere çok yüksek derecede FosfoFruktoKinaz (PFK) tarafından allosterik olarak Düzenlenir.Metabolizmanın entegrasyonun da önemli rol oynar. G°´ = kJ/mol ΔG°’= -3.4 kcal mol-1
 tarafından allosterik olarak Düzenlenir.Metabolizmanın entegrasyonun da önemli rol oynar. G°´ = kJ/mol. ΔG°’= -3.4 kcal mol-1.")
44
Aldolaz G°´= kJ/mol ΔG°’= 5.7 kcal mol-1

45
Trioz fosfat İzomeraz - Yalnız ca GLİSERALDHİT-3-FOSFAT ile glikoliz devam ettirilir. - aldoz-ketoz izomerizasyonu G-6-P izomerizasyonuna benzer. - Reaksiyonun Tersinir olması dihidroksiasetonun metabolize edilmesine olanak sağlar.Bu GLUKONEOGENEZ de oldukca önemlidir. G°´= +7.5 kJ/mol Bu tersinir reaksiyon ASİT-BAZ katelizi ile sağlanır.(Enz-Histidin-95 ve Glutamat-165)
")
46
************************************************
1. Evrenin sonunda: - 2 gliseraldehit oluşturuldu 2 ATP kullanıldı. Enerji üretimi bundan sonraki basamaklarda olacak.

47
-Yüksek enerjili (karboksilik-fosforikasit anhidrat) ilk bileşik.
2.EVRE GAL-3-P Dehidrogenaz -Yüksek enerjili (karboksilik-fosforikasit anhidrat) ilk bileşik. Arsenik GAPDH enzim inhibitörüdür.
 ilk bileşik. Arsenik GAPDH enzim inhibitörüdür.")
48
Gliseraldehit-3-P Dehidrogenaz.GALPDH

49
SUBSTRAT DÜZEYİ FOSFORİLASYONLA ilk ATP üretimi.
Fosfogliserat kinaz SUBSTRAT DÜZEYİ FOSFORİLASYONLA ilk ATP üretimi. Buraya kadar harcanan ATP üretilmiş olur (2 mol ATP)
")
50
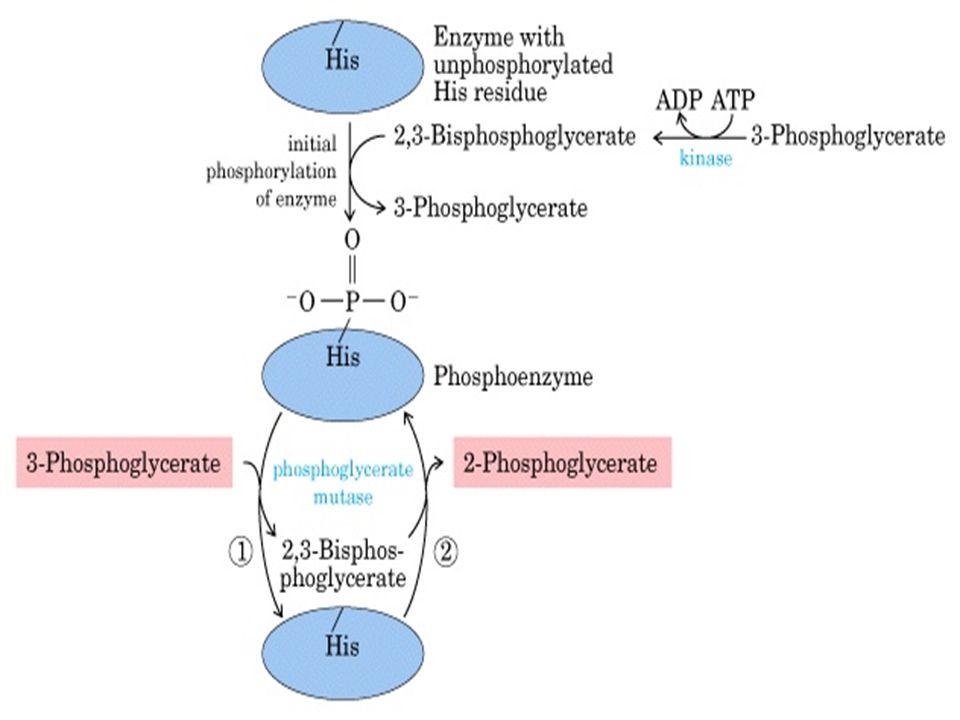
Fosfogliserat MUTAZ

52
Fosfogliserat mutaz ‘ın etki mekanizması:

53
*****Florid iyonu enolazın kompetetif inhibitörüdür.

54
ENOLAZ’ın etki mekanizması:

55
Pirüvat KİNAZ Glikoliz de ATP oluşturmak için gerçekleşen 2. SUBSTRAT DÜZEYİ FOSFORİLASYON

57
Glikolizde enerji blançosu:
Harcanan enerji = 2 ATP 1. glukoz + ATP glucose-6-P 2. fruktoz-6-P + ATP fruktoz1,6 bisfosfat Üretilen enerji = 4 ATP + 2 NADH 2 gliseraldehit-3-P + 2 Pi + 2 NAD+ 2 (1,3 bisfosfogliserat) + 2 NADH 2. 2 (1,3 bisfosfogliserat) + 2 ADP2 (3-P-gliserat) ATP 3. 2 PEP + 2 ADP 2 pirüvat + 2 ATP Net = 2 ATP and 2 NADH
 + 2 NADH (1,3 bisfosfogliserat) + 2 ADP2 (3-P-gliserat) + 2 ATP PEP + 2 ADP 2 pirüvat + 2 ATP. Net = 2 ATP and 2 NADH.")
58
Glukoz + 2NAD+ = 2Pirüvat + 2NADH + 2H+ G°´ = -146 kJ/mol
2ADP + 2Pi = 2ATP + 2H2O G°´ = 2X(30.5 kJ/mol) = 61 kJ/mol G°´ (toplam) = = -85 kJ/mol Glikolizin standart koşullar altında ekzergonik bir reaksiyon olmasından mekanizma TERSİNMEZ dir. Glukoz + 6O2 = 6CO2 + 6H2O G°´ = kJ/mol Glukoz + 2NAD+ = 2Pirüvat + 2NADH + 2H G°´ = -146 kJ/mol Glikolizde piruvat oluşumu ile salınan serbest enerji toplam serbest enerjinin %5.2 sini oluşturur.
 = 61 kJ/mol G°´ (toplam) = = -85 kJ/mol. Glikolizin standart koşullar altında ekzergonik bir reaksiyon olmasından mekanizma TERSİNMEZ dir. Glukoz + 6O2 = 6CO2 + 6H2O G°´ = kJ/mol Glukoz + 2NAD+ = 2Pirüvat + 2NADH + 2H+ G°´ = -146 kJ/mol Glikolizde piruvat oluşumu ile salınan serbest enerji toplam serbest enerjinin %5.2 sini oluşturur.")
59
Egzersiz yapan kaslar,eritrosit Bazı mikroorganizmalar.
Glukoz Glikoliz (10 ardıl tepkime)) Anaerobik koşullarda Anaerobik koşullarda 2 Pirüvat Aerobik koşullarda 2 Etanol + 2 CO2 2 Laktat Mayalarda etanol fermantasyonu LAKTAT FERMANTASYONU Egzersiz yapan kaslar,eritrosit Bazı mikroorganizmalar. 2 Asetil-CoA TCA döngüsü 4 CO2 + 6 H2O Aerobik koşullar altında ki Hayvan,bitki ve mikrobial hücreler.
) Anaerobik. koşullarda. Anaerobik. koşullarda. 2 Pirüvat. Aerobik. koşullarda. 2 Etanol + 2 CO2. 2 Laktat. Mayalarda etanol. fermantasyonu. LAKTAT FERMANTASYONU. Egzersiz yapan kaslar,eritrosit. Bazı mikroorganizmalar. 2 Asetil-CoA. TCA. döngüsü. 4 CO2 + 6 H2O. Aerobik koşullar altında ki. Hayvan,bitki ve mikrobial hücreler.")
60
Homolaktik Fermentatasyon:
-Anaerobik koşullar altında yada çok ani ATP ihtiyacı gerektiğinde , ATP üretiminde ana kaynak GLİKOLİZDİR. -NAD+ ise GAPDH reaksiyonu için çok önemli bir KOFAKTÖR dür.NADH dan NAD+ oluşturulması kas hücrelerinde bu reaksiyon ile olmaktadır. -LDH, pirüvatı laktata indirgeyerek NADH’ ı oksidasyona uğratır ve NAD+ oluşturur .Her bir laktat oluşumu için 1 NADH harcanır.(2 laktat 2 NADH).Net enerji 2 ATP dir. laktat fermentasyonu aşırı egzersizde iskelet kasında (her nekadar kas yorg.), eritrosit ve retinada önemlidir.Aynı zamanda O2 ‘siz koşullarda bitki ve mikropların üreyip büyümeleri için de gereklidir. LAKTAT Dehidrogenaz
.Net enerji 2 ATP dir. laktat fermentasyonu aşırı egzersizde iskelet kasında (her nekadar kas yorg.), eritrosit ve retinada önemlidir.Aynı zamanda O2 ‘siz koşullarda bitki ve mikropların üreyip büyümeleri için de gereklidir. LAKTAT. Dehidrogenaz.")
61
Alkolik fermentasyon:
Mikroorganizmalar ve mayalar glikoliz için (6.tep.GAPDH) gerekli NAD+’ı pirüvatı alkole ve karbondioksite dönüştürerek sağlarlar. Bu 2 basamaklı bir prosestir: Piruvat dekarboksilaz (PDK) reaksiyonu: Bu enzim Mg++ bağımlıdır ve ayrıca enzime-bağlı kofaktöre gereksinir.TİAMİN PİROFOSFAT (TPP). Bu reaksiyonda 1 mol CO2 salınır ve asetaldehit oluşturulur. Alkol dehidrogenaz reaksiyonu: Asetaldehit NADH aracılığı ile etanole indirgenirken glikoliz için de NAD+ sağlanır . Pirüvat Dekarboksilaz Alkol Dehidrogenaz
 gerekli NAD+’ı pirüvatı alkole ve karbondioksite dönüştürerek sağlarlar. Bu 2 basamaklı bir prosestir: Piruvat dekarboksilaz (PDK) reaksiyonu: Bu enzim Mg++ bağımlıdır ve ayrıca enzime-bağlı kofaktöre gereksinir.TİAMİN PİROFOSFAT (TPP). Bu reaksiyonda 1 mol CO2 salınır ve asetaldehit oluşturulur. Alkol dehidrogenaz reaksiyonu: Asetaldehit NADH aracılığı ile etanole indirgenirken glikoliz için de NAD+ sağlanır . Pirüvat. Dekarboksilaz. Alkol. Dehidrogenaz.")
62
Tiamin pirofosfat (TPP) Pirüvat dekarboksilaz etki mekanizması
Nükleofilik atak Tiamin pirofosfat (TPP) TPP nin eklenmesi İle CO2 ‘in ayrılması Karbanyonun protonlanması Pirüvat dekarboksilaz etki mekanizması
 TPP nin eklenmesi. İle CO2 ‘in. ayrılması. Karbanyonun. protonlanması. Pirüvat dekarboksilaz etki mekanizması.")
63
Böylece redoks tepkimesinde net bir değişim olmamakta ve stokiyometrik
Olarak eşitlik sağlanmaktadır.

64
1 2 3 6 4 7 5 8 9 10
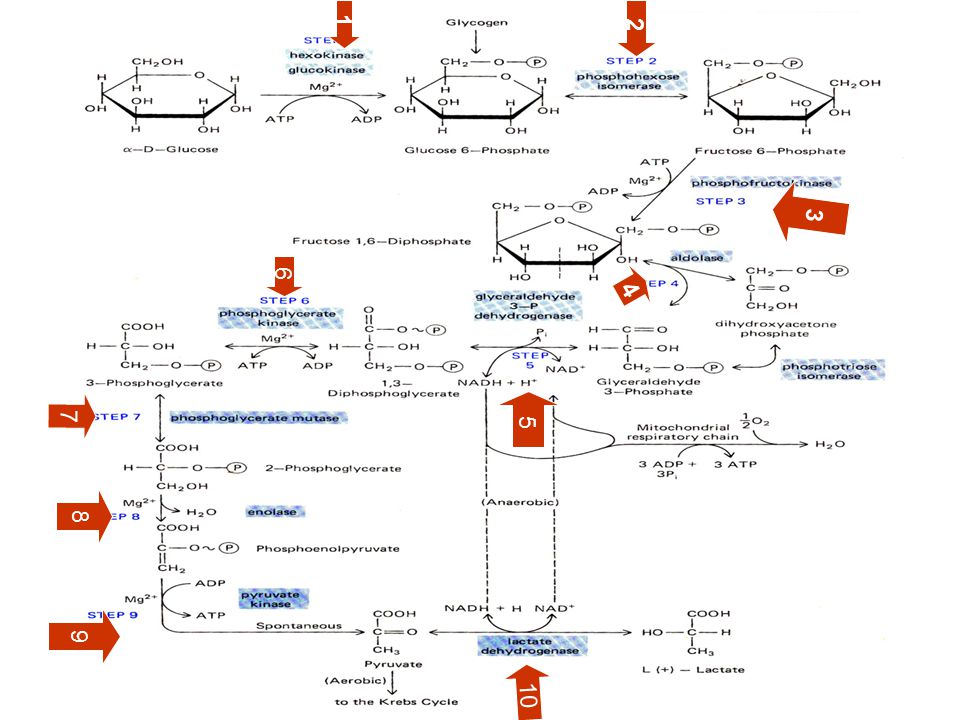
65
GLİKOLİZ’İN DÜZENLENMESİ:
METABOLİK REAKSİYONLAR İÇİN İKİ TİP KONTROL VARDIR: a) Substratla sınırlanan : Hücre içerisinde reaktan ve ürünlerin konsantrasyonları denge durumuna yaklaşmışsa, bu durumda reaksiyon yönü S varlığına bağlıdır. b) Enzimle-sınırlanan: Substrat ve ürün konsantrasyonu denge derişimine ulaşmasından çok fazla ise , reaksiyon hızı enzim aktivitesi ile belirlenir.Böylesi reaksiyonlar tüm metabolik reaksiyonların akışını kontrol eden tek reaksiyonlardır. Glikoliz’de glikolizin akışını düzenleyen 3 enzimatik basamak vardır. Hekzokinaz (HK) Fosfofrukto kinaz(PFK)********(pastör etkisi) Pirüvat kinaz (PK)
 Substratla sınırlanan : Hücre içerisinde reaktan ve ürünlerin konsantrasyonları denge durumuna yaklaşmışsa, bu durumda reaksiyon yönü S varlığına bağlıdır. b) Enzimle-sınırlanan: Substrat ve ürün konsantrasyonu denge derişimine ulaşmasından çok fazla ise , reaksiyon hızı enzim aktivitesi ile belirlenir.Böylesi reaksiyonlar tüm metabolik reaksiyonların akışını kontrol eden tek reaksiyonlardır. Glikoliz’de glikolizin akışını düzenleyen 3 enzimatik basamak vardır. Hekzokinaz (HK) Fosfofrukto kinaz(PFK)********(pastör etkisi) Pirüvat kinaz (PK)")
66
HEKZOKINAZ Glikolizde glukozun fosforlandığı ilk basamak
HEKZOKINAZ Glikolizde glukozun fosforlandığı ilk basamak. Kendi ürünü olan glukoz-6-P tarafından inhibe edilir. Glikoliz yavaşlar. 2. FOSFOFRUKTO KİNAZ (PFK) Glikoliz de hız sınırlayıcıdır. Allosterik yapısı olan multimerik düzenleyici bir enzimdir. Enerji düzeylerinin yeterliliği/yetersizliği ile sınırlanır. İnhibitörleri: ATP ve Sitrat 2ADP ATP +AMP yüksek enerji Aktivatörleri: ADP, AMP, ve fruktoz 2,6 bisfosfat düşük enerji
 Glikoliz de hız sınırlayıcıdır. Allosterik yapısı olan multimerik düzenleyici bir enzimdir. Enerji düzeylerinin yeterliliği/yetersizliği ile sınırlanır. İnhibitörleri: ATP ve Sitrat. 2ADP ATP +AMP yüksek enerji. Aktivatörleri: ADP, AMP, ve. fruktoz 2,6 bisfosfat. düşük enerji")
67
Boş Döngü (futile Cycle)
ATP + Fruktoz-6-P → ADP + Fruktoz-1,6-bisP + H+ Fruktoz-1,6-bisP + H2O → Fruktoz-6-P + Pi TOP: ATP + H2O → ADP + Pi + H+ + ISI Hiçbir metabolik iş yapılmadan ATP harcanmıştır.Bu nedenle bu döngüye FUTİLE DÖNGÜ denir.

68
ATP kullanıldığında ADP miktarı artar, adenilat siklaz aracılığı ile 2ADP ATP +AMP oluşturlur.Bu da glikolizin aktivasyonunu artırır. Eritrositlerde [ATP]= 1.5 mmol/L,[AMP]= 0.03 , ATP deki küçük bir değişim AMP miktarında çok büyük değişimlere neden olur.PFK-1 AMP deki değişimlere karşı oldukça duyarlıdır. K.c ve eritrositlerde F-1,6-BP ile düzenleme feed-forward regülasyonu olarak bilinir.Eritrosit sitozolin de sınırlı sayıda biriken triozP araürünlerinin artmasından dolayı önemli olabilir.(glioksilatlar). Eritrositlerde glikoliz ürünü olan 2,3-BPG (5 mmol/L), ATP ve Pi derişimlerinin yaklaşık 5 katıdır.Bu ara ürün Hb nin (-)allosterik efektörüdür ve O2 ye olan ilgisini azaltır.Denizden çok yüksek yerlerde (adaptasyon) ve anemik koşullarda periferik dokulara O2 salınımını artırır. Ayrıca fetal Hb (HbF) üzerinde etkisi az olduğundan anne eritrositlerin de artan 2,3-BPG HbA dan O2 nin HbF ye aktarılması sağlanır.
![ATP kullanıldığında ADP miktarı artar, adenilat siklaz aracılığı ile 2ADP ATP +AMP oluşturlur.Bu da glikolizin aktivasyonunu artırır. Eritrositlerde [ATP]= 1.5 mmol/L,[AMP]= 0.03 , ATP deki küçük bir değişim AMP miktarında çok büyük değişimlere neden olur.PFK-1 AMP deki değişimlere karşı oldukça duyarlıdır.](http://slideplayer.biz.tr/slide/3046389/11/images/68/ATP+kullan%C4%B1ld%C4%B1%C4%9F%C4%B1nda+ADP+miktar%C4%B1+artar%2C+adenilat+siklaz+arac%C4%B1l%C4%B1%C4%9F%C4%B1+ile+2ADP+ATP+%2BAMP+olu%C5%9Fturlur.Bu+da+glikolizin+aktivasyonunu+art%C4%B1r%C4%B1r.+Eritrositlerde+%5BATP%5D%3D+1.5+mmol%2FL%2C%5BAMP%5D%3D+0.03+%2C+ATP+deki+k%C3%BC%C3%A7%C3%BCk+bir+de%C4%9Fi%C5%9Fim+AMP+miktar%C4%B1nda+%C3%A7ok+b%C3%BCy%C3%BCk+de%C4%9Fi%C5%9Fimlere+neden+olur.PFK-1+AMP+deki+de%C4%9Fi%C5%9Fimlere+kar%C5%9F%C4%B1+olduk%C3%A7a+duyarl%C4%B1d%C4%B1r..jpg "K.c ve eritrositlerde F-1,6-BP ile düzenleme feed-forward regülasyonu olarak bilinir.Eritrosit sitozolin de sınırlı sayıda biriken triozP araürünlerinin artmasından dolayı önemli olabilir.(glioksilatlar). Eritrositlerde glikoliz ürünü olan 2,3-BPG (5 mmol/L), ATP ve Pi derişimlerinin yaklaşık 5 katıdır.Bu ara ürün Hb nin (-)allosterik efektörüdür ve O2 ye olan ilgisini azaltır.Denizden çok yüksek yerlerde (adaptasyon) ve anemik koşullarda periferik dokulara O2 salınımını artırır. Ayrıca fetal Hb (HbF) üzerinde etkisi az olduğundan anne eritrositlerin de artan 2,3-BPG HbA dan O2 nin HbF ye aktarılması sağlanır.")
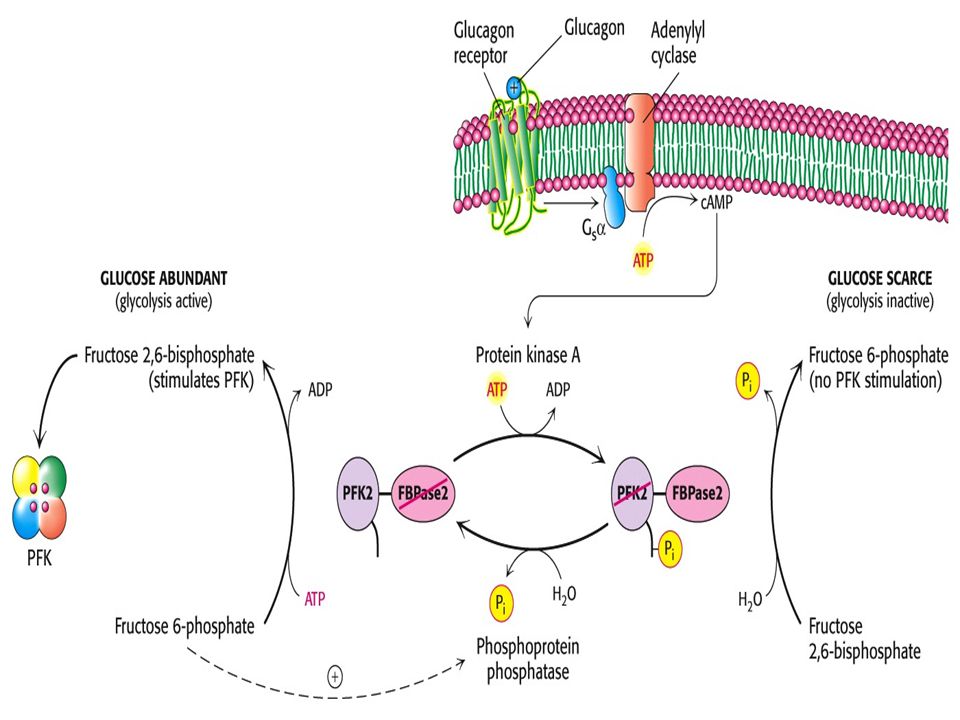
69
FBPaz 2 PFK2 BİFONKSİYONEL enzim, düzenleyici bölge fosforlandığında PFK-2 inaktif , FBPaz aktif. Defosforile olduğunda ise PFK-2 aktif,FBPaz ise inaktif olmaktadır. Bu modifikasyon glukagon hormonu aracılığı ile olmaktadır.GLİKOLİZ ve GLİKONEOGENEZ metabolik yolak düzenlenir. AMP, ADP ve Fruktoz 2, 6 bisfosfat bu enzimin allosterik aktivatötüdür.

70
Fosfofruktokinaz 2 (PFK-2) F-6-P dan F-2,6-BP oluşumunu katalizler.
F-2,6-BP hayvan ve bitki hücrelerinde bulunur. PFK nın aktivitesini arttıran allosterik bir aktivatördür. PFK nın affinitesini F-6-P a karşı arttırarak glikoliz stimüle edilir. Fosfofruktokinaz 2 (PFK-2) F-6-P dan F-2,6-BP oluşumunu katalizler. Fruktoz bisfosfataz 2 (FBPaz-2), F-2,3-BP tı tekrar F-6-P a dönüştürür.Glikoliz inhibe olur.
 F-6-P dan F-2,6-BP oluşumunu katalizler. Fruktoz bisfosfataz 2 (FBPaz-2), F-2,3-BP tı tekrar F-6-P a dönüştürür.Glikoliz inhibe olur.")
72
Pirüvat kinaz ın glikolizdeki düzenleyici rolü:
Pirüvat Kinaz allosterik olarak düzenlenen,tetramer yapıda, L (liver) ve M (muscle and brain) formuna sahip, ve kendi ürünü olan ATP tarafından inhibe edilir. Aynı zamanda uzun zincirli yağ asitleri ve asetilCoA tarfından da inhibisyona uğrar. Fruktoz 1,6-bisfosfat (feed-forward) enzimin her iki formunu aktive eder. Transaminasyon tepkimelerinde alanin aa’i pirüvata tersinir olarak dönüşebilmektedir.Yüksek alanin derişimi PK ı inhibe edebilir. Kan glukoz düzeyinin durumuna göre PK nın regüle edilmesi yine glukagon hormonu aracılığı ile sağlanır.
 ve M (muscle and brain) formuna sahip, ve kendi ürünü olan ATP tarafından inhibe edilir. Aynı zamanda uzun zincirli yağ asitleri ve asetilCoA tarfından da inhibisyona uğrar. Fruktoz 1,6-bisfosfat (feed-forward) enzimin her iki formunu aktive eder. Transaminasyon tepkimelerinde alanin aa’i pirüvata tersinir olarak dönüşebilmektedir.Yüksek alanin derişimi PK ı inhibe edebilir. Kan glukoz düzeyinin durumuna göre PK nın regüle edilmesi yine glukagon hormonu aracılığı ile sağlanır.")
73
Bu kovalent modifikasyon kas ve beyin glukoz gereksinimini karşılamak üzere gerçekleşir.

74
Anaerobic glycolysis
computing Presentation copyright © 2002 David A Bender and some images copyright © 2002 Taylor & Francis Ltd Gliloliz: Glukozun anaerobik metabolizması; CORİ DÖNGÜSÜ Anaerobic glycolysis

75
GLİKOJEN PENTOZ FOSFAT METABOLİZMASI GLUKOZ GLUKOZ-6-FOSFAT GLİKOLİZ

76
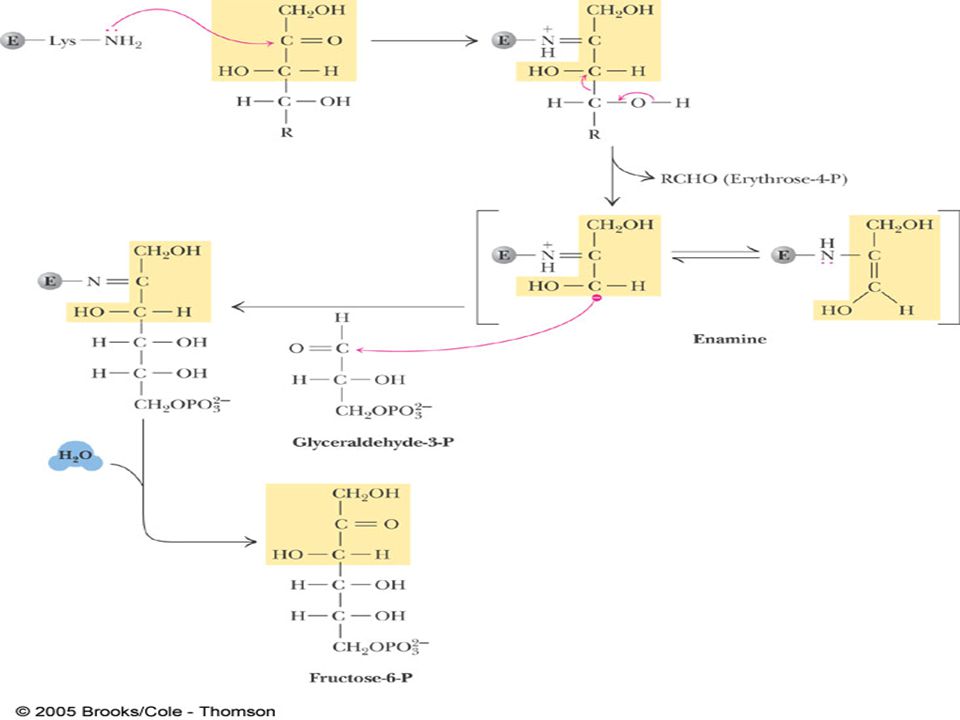
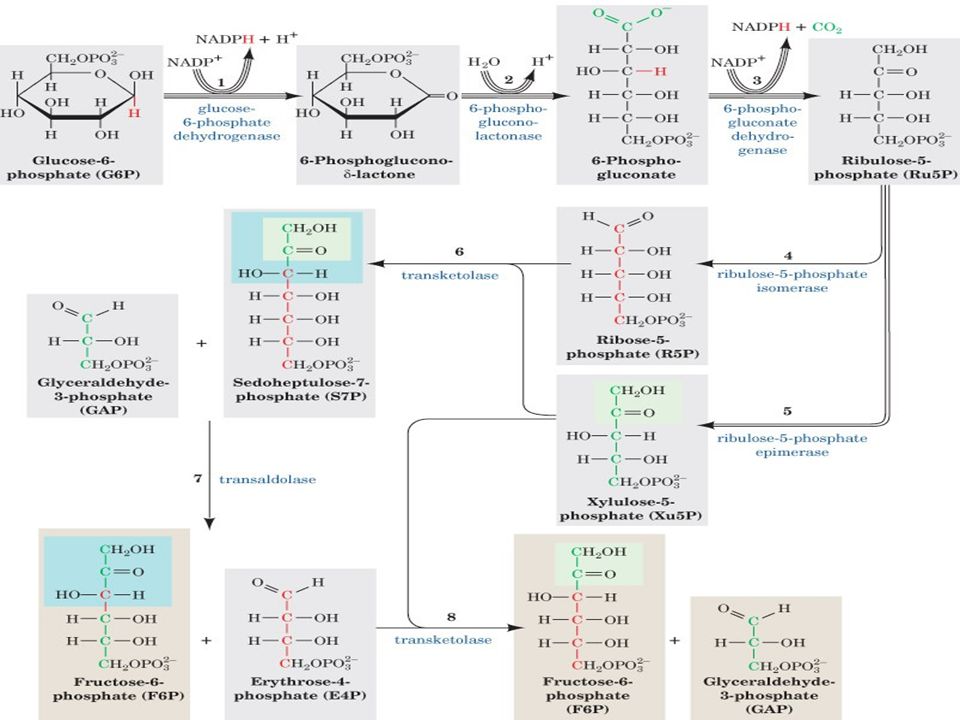
PENTOZ FOSFAT YOLU:( Entner-Doudoroff pathways)
Hekzos mono fosfat yolu (SHUNT) olarakta bilinir. G-6-P dan başlar, metabolizma için gerekli NADPH ve RİBOZ üretilir. Gal-3-P ve F-6-P ara ürünleri ile glikolizi aktive eder. Bu özelliğinden dolayı SHUNT olarak ifade edilir.
 olarakta bilinir. G-6-P dan. başlar, metabolizma için gerekli NADPH ve RİBOZ üretilir. Gal-3-P ve F-6-P ara ürünleri ile glikolizi aktive eder. Bu özelliğinden dolayı SHUNT olarak ifade edilir.")
77
NADPH gereksinen yollar. SENTEZ: - Yağ asit biosentezi
-Kolesterol biosentezi - Nörotransmitter biosentezi - Nükleotid biosentezi DETOKSİFİKASYON: -Okside Glutatyonun indirgenmesi, -Sitokrom P450 monooksigenaz Glikoliz için Ara (3 ve 6 C) Metabolit üretimi CoA, FAD, SAM, NAD+/NADP+
 Metabolit. üretimi. CoA, FAD, SAM, NAD+/NADP+")
78
PENTOZ FOSFAT SHUNT’ ININ AKTİF OLDUĞU DOKULAR
DOKU FONKSİYONU Adrenal gland Steroid sentezi Karaciğer Yağ asiti ve kolesterol sentezi Testis ve Ovaryum Steroid sentezi Adipoz doku Yağ asit sentezi Meme bezleri Yağ asit sentezi Eritrosit İndirgenmiş GLUTATYONUN idamesi Hekzos Mono Fosfat (HMF) yolu hücre SİTOZOLÜN de yer alan amfibolik (aerobik veya anaerobik) bir pathway dir.
 yolu hücre SİTOZOLÜN de yer alan amfibolik (aerobik veya anaerobik) bir pathway dir.")
79
Çok hızlı bölünen hücreler NADPH dan çok RİBOZ-5-P a ihtiyaçları vardır.

80
G-6-P Dehidrogenaz düzenleyici bir enzimdir. NADP+
G-6-P Dehidrogenaz düzenleyici bir enzimdir. NADP+.ye karşı oldukça spesiktir.(Km değeri NAD+ ya göre1000 kez küçüktür). Laktonaz eksikliğinde halkasal yapının açılma hızı çok yavaşlar bu nedenle 6-Fosfoglukonolakton birikir, toksik etki gösterir. Laktonaz
. Laktonaz eksikliğinde halkasal. yapının açılma hızı çok yavaşlar. bu nedenle 6-Fosfoglukonolakton. birikir, toksik etki gösterir. Laktonaz.")
81
6-fosfoglukonat dehidrogenaz

82
NADPH,, anabolik (sentetik) yolaklarda redüktant olarak fonksiyon görür, ör., yağ asit sentezi.
NAD+, katabolik yolaklarda é akseptörü olarak görev alır.

83
Oksidasyon basamağı tersinmezdir.

84
EPİMERAZ KSİLULOZ-5-P RİBULOZ-5-P İZOMERAZ RİBOZ-5-P

85
TRANSKETOLAZ

86
Transketolaz enzimi prostetik grup olarak TİAMİN PİROFOSFAT (TPP)(Vit B1) grubuna gereksinimi vardır. Tiazolyum halkası Aminoprimidin halkası

88
TRANSALDOLAZ

91
Hücrede hem NADPH hemde RİBOZ-5-P’ a ihtiyaç varsa ilk 5 reaksiyon baskındır.

92
Pirimer olarak Riboz-5-P gereksinimi varsa Pentoz Fosfat Yolunun Oksidasyon basamağı bypass olabilir.

93
Çok miktarda NADPH üretilebilir, döngünün yeniden oluşabilmesi için G-6-P oluşturulur.

94
Hem ATP hemde NADPH gereksinimi olduğunda HMF yolu GLİKOLİZ’e yönlenebilir.
Bu bölünmeleri için DNA ve RNA ya ihtiyacı olmayan hücrelerde oldukca önemlidir.(Eritrosit)
")
95
PENTOZ FOSFAT YOLUNUN DÜZENLENMESİ:
Metabolik yolun kullanımı temelde enerji gereksinimine bağlıdır.(sentez’ e karşı glikoliz, yada NADH üretmek için HMF yolu). Metabolik yolun kararlılığı G-6-P ‘ı kullanan Fosfoglukozizomeraz ve Fosfofrukto kinaz (Glikoliz) aktivitelerine karşı G-6-P dehidrogenaz (HMF) aktivitesi belirleyecektir. Bundan dolayı, PFK aktivitesi ATP/AMP düzeyine bağlıdır. G-6-PDH aktiviteside NADPH ve yağ asitlerine bağlıdır.
. Metabolik yolun kararlılığı G-6-P ‘ı kullanan Fosfoglukozizomeraz ve Fosfofrukto kinaz (Glikoliz) aktivitelerine karşı G-6-P dehidrogenaz (HMF) aktivitesi belirleyecektir. Bundan dolayı, PFK aktivitesi ATP/AMP düzeyine bağlıdır. G-6-PDH aktiviteside NADPH ve yağ asitlerine bağlıdır.")
96
Glukoz- 6-fosfat DH düzenleyici bir enzimdir
Glukoz- 6-fosfat DH düzenleyici bir enzimdir.İnsülin tarafından indüklenebilir. NADPH enzimin etkili bir kompetetiv inhibitörüdür. Genellikle sitoplazmik NADPH/NADP+= 100 dür. Bu oranın üzerinde enzim inhibe edilir. G-6-PDH eksikliğide çok ciddi bir sorundur. Bu tür kişilerde yeterince NADPH üretilemediğinden eritrositlerde glutatyon GSH rejenere edilemez, reaktif oksijen radikallerinden korunamaz ve hemolitik anemi gelişir.

97
Glukoz 6-Fosfat Dehidrogenaz Yetmezliği
Homozigot mutation: yüksek hemolizis ve anemi Heterozigot mutation: Normall de semptom yok Radikal oksijen türlerini (ROS,ROT) oluşturan ilaçlar (Kinin ve türevleri) kullanılmadıkça, ve Bakla türü besinler tüketmedikce. Dünyada 300 milyondan fazla insan etkilenmekte. 400 bin çeşit G6PDH eksikliğine rastlanmış.(genetik kökenli enzimopati) X kromozomuna bağlı bir gen ürünü olması erkekleri etkilemektedir. Mutasyon (homozigot) yalnız bir baz değişikliğine bağlı olarak aa diziliminde etkili.Daha çok Akdeniz popülasyonu, Afrika, Asya Malarya hastalığı ile benzer özellikler göstermektedir.Enzim eksikliği bu hastalığa direnci de etkileyebilmektedir. Özellikle eritrosit de mitokondri olmaması bu enzm eksikliğinde yeterince NADPH üretilemediğinden ORAK HÜCRE ANEMİSİ oluşmakta.
 oluşturan ilaçlar (Kinin ve türevleri) kullanılmadıkça, ve Bakla türü besinler tüketmedikce. Dünyada 300 milyondan fazla insan etkilenmekte. 400 bin çeşit G6PDH eksikliğine rastlanmış.(genetik kökenli enzimopati) X kromozomuna bağlı bir gen ürünü olması erkekleri etkilemektedir. Mutasyon (homozigot) yalnız bir baz değişikliğine bağlı olarak aa diziliminde etkili.Daha çok Akdeniz popülasyonu, Afrika, Asya. Malarya hastalığı ile benzer özellikler göstermektedir.Enzim eksikliği bu hastalığa direnci de etkileyebilmektedir. Özellikle eritrosit de mitokondri olmaması bu enzm eksikliğinde yeterince NADPH üretilemediğinden ORAK HÜCRE ANEMİSİ oluşmakta.")
98
Oksidatif strese neden olan diğer ilaçlar.
Sulfonamidler (antibiotik) Aspirin , Nitrofurantoin,Quinadin ve kinin, Naftalin Bakla (visin & izouramil) Ayrıca diğer bakterial ve viral enfeksiyonlarda neden.
 Aspirin , Nitrofurantoin,Quinadin ve kinin, Naftalin. Bakla (visin & izouramil) Ayrıca diğer bakterial ve viral enfeksiyonlarda neden.")
99
Düşük RBC sayımı & düşük hemoglobin
malarya Siyah renkte idrar Hemoliz sonucu Hb nin atılımının artması. Düşük RBC sayımı & düşük hemoglobin Yük.hemoliz Artmış bilirübin Hem yıkımı FAVİZM: Akdeniz tipi G6PDH eksiliği olan kişiler baklaya karşı oldukça hassastır. Sindirim sonrası açığa çıkan oksitleyici ajanların (Polihidroksipirimidin -glukositler, divisin ve izouramil) hücre hasarı ortadan kaldırılamadığından bu hastalık oluşturmaktadır.
 hücre hasarı ortadan kaldırılamadığından bu hastalık oluşturmaktadır.")
100
ANTİOKSİDAN MEKANİZMALAR ve OKSİJEN RADİKALLERİ:
Monooksigenazlar: Mikrozomal P450 sistem flavoprotein kompleksleri, sitokrom P450, FeS & protein -Yabancı kimyasal bileşiklerin HİDROKSİLASYON tepkimelerini katalizlerler. Benzo[a]piren aren oksid Guanine kovalent olarak bağlanan bir karsinojen.

101
Sitokrom P450 Enzimleri Karaciğer düz endoplazmik reticulm da
Gut mukoza Çok küçük miktarlarda akciğer mukoza,böbrek,deri ve beyin Monooksigenaz reaksiyonları: NADPH + H+ + O2 + SH NADP+ + H2O + SOH Monooksigenasyon denmesi tek bir atom’u oksidasyona uğrattığı için.

102
NADPH bir é ‘u Flavoprotein reductaz’ a verir.
Sitokrom P450 Döngüsü NADPH bir é ‘u Flavoprotein reductaz’ a verir. Flavoprotein reductase oksitlenmiş cyt P450-ilaç kompleksini indirger. Molecular O2 é alır ve aktive olur, “activated O2”-cytoch-kompleki oksitlenir. O2*- H2O2 2H+ R.OH

103
O2 sensör & taşınım 1) O2 taşıyıcıları : Hb & Mb
2) O2 sensör : Heme protein I. NADPH oxidase (cytochrome b) Mitokondrial Elektron Taşınımı : cytochrome c oxidase O2 nin% 85–90 kullanır. ( O2’ ‘e olan afinitesi çok yüksek) Oxidase & oxygenase (cytosolic) : % of O2 ex) cytochrome p
 O2 sensör : Heme protein. I. NADPH oxidase (cytochrome b) Mitokondrial Elektron Taşınımı. : cytochrome c oxidase O2 nin% 85–90 kullanır. ( O2’ ‘e olan afinitesi çok yüksek) Oxidase & oxygenase (cytosolic) : 10-15% of O2. ex) cytochrome p")
104
O2 Radikal nasıl oluşur: Tek elektron kaybı X e- + X*+ 21% (159mmHg)
2) Tek elektron alımı X + e- X*- 3) Bir kovalent bağın Homolitik kırılması homolytic fission A:B A* + B* heterolytic fission A:B A- + B+ O2 21% (159mmHg) Dokularda : 40mmHg Intracellular : 1-10mmHg gradient mitokondri içinde : < 0.5mmHg
 Tek elektron alımı. X + e- X*- 3) Bir kovalent bağın Homolitik kırılması. homolytic fission. A:B A* + B* heterolytic fission. A:B A- + B+ O2. 21% (159mmHg) Dokularda : 40mmHg. Intracellular : 1-10mmHg. gradient. mitokondri içinde. : < 0.5mmHg.")
105
Moleküler oksijen paralel spinli iki çiftleşmemiş é taşıyan çift merkezli yapıdadır.
p* 2p 2p s* 2s 2s s* 1s s 1s Ground state O2 superoksit O2*- singlet O2 singlet O2 (93.6 kJ) (Activated, 157kJ) Generated by energy input
 (Activated, 157kJ) Generated by. energy input.")
106
Oksijen toksitesinde süperoksit teorisi: Neden superoksit sitotoksik?
( O2*- suya göre organik çözücülerde 8-10 kat daha fazla çözündüğünden önemli bir faktördür. ) O2*- H2O2 O2 SOD e- Neden superoksit sitotoksik? 1) Superoksit, diğer antioxidant defans enzimlerin aktivitelerini azaltabilir. Katalaz, Glutathione peroxidase 2) Direct etkilediği enzimler: 6-phosphogluconate dehydratase, aconitase, fumarase mitochondrial NADH dehydrogenase creatine kinase ribonucleotide reductase calcineurin 4) Diğer oksijen radikallerini oluşturması.
 O2*- H2O2. O2. SOD. e- Neden superoksit sitotoksik 1) Superoksit, diğer antioxidant defans enzimlerin aktivitelerini azaltabilir. Katalaz, Glutathione peroxidase. 2) Direct etkilediği enzimler: 6-phosphogluconate dehydratase, aconitase, fumarase. mitochondrial NADH dehydrogenase. creatine kinase. ribonucleotide reductase. calcineurin. 4) Diğer oksijen radikallerini oluşturması.")
107
ROS oluşumu H2O2 OH* O2*- HO2* ONOO- 2H+ e- e- Fe++ O2 SOD Fenton Rxn
(Hidroksill radikali) H+ HO2* (Hidroperoksil radikali) NO* ONOO- (Peroksinitrit)
 H+ HO2* (Hidroperoksil radikali) NO* ONOO- (Peroksinitrit)")
108
Reaktif oksijen türlerinin üretildiği ve detoksifiye edildiği yer PEROKSİZOM’lardır.İlk 1950’lerde hayvan hücrelerinde, 1960’ larda ise bitki hücrelerinde elektron mikroskobu ile gözlemlendi. 1965’de karaciğerden izole edildi.H2O2 yi hem oluşturup hemde yıktığı için peroksizom dendi. Glukoz sınırlı Daha fazla Glu. sınırlı Metanol sınırlı

109
Peroksizomların metabolik Fonksiyonları:
Maya Biosentez: lizin Katabolizma: aa, metanol,ya -oksidasyon, H2O2, glioksilat döngüsü. Mantar Biosentez: penicillin Katabolizzma:ya -oxidation H2O2, glioksilat döngüsü. Bitki Katabolizma: purinler, fotosentezin bazı reaksiyonları (Glikolat’ın , glisine ve serin’in ,gliserat’a dönüştürülmesi.),ya -oxidation, H2O2 ,glioksilat dön. Memeli Biosentez: eter fosfolipid (plazmalojen), kolesterol ve safra asiti, poliunsature ya. Katabolizma: aa, pürinler,prostaglandin,poliamin,ya,- -oksidas, H2O2
,ya -oxidation, H2O2 ,glioksilat dön. Memeli. Biosentez: eter fosfolipid (plazmalojen), kolesterol ve safra asiti, poliunsature ya. Katabolizma: aa, pürinler,prostaglandin,poliamin,ya,- -oksidas, H2O2.")
110
Peroksizomal Enzimler.

111
Reactive Oksijen Türleri (ROS)
Radikaller Radikal olmayanlar Superoksit, O2* Hidrogen peroksit, H2O2 Hidroksil, OH* Hipoklorik asit, HOCl Peroksil, RO2* Ozon, O3 Alkoksil, RO* Singlet oksijen, 1 g Hidroperoksil, HO2* Peroksinitrit, ONOO-

112
ROs ve ANTİOKSİDANLAR REAKTİF TÜRLER ANTİOKSİDANLAR Singlet oksijen 1O2 vitamin A, vitamin E Superoksit radikal O2- superoksid dismutaz, vitamin C Hidrojen peroksit H2O2 Katalaz, glutathione peroxidase Peroksil radical ROO vitamin C, vitamin E Lipid peroxyl radical LOO vitamin E Hydroxyl radical OH vitamin C

113
ROS Kaynak Endojen Eksojen prostaglandin sente. radiation, ultrasound
respiratory chain cigarette smoke autooxidation drugs SERBEST RADİKALLER phagocytes heat oxyhemoglobin pesticides oxidative enzymes infections accumul reduced.metab hyperoxia, exercise air pollution (NOx, O3)
")
114
ROS’un enzimatik kaynakları
Xanthine oxidase Hypoxanthine + 2O2 Xanthine + O2.- + H2O2 NADPH oxidase NADPH + O2 NADP+ + O2.- Amine oxidases R-CH2-NH2 + H2O + O2 R-CHO + NH3 + H2O2 Myeloperoxidase Hipohalojen asit oluşumu H2O2 + X- + H+ HOX + H2O NADH oxidase reaction Hb(Mb)-Fe3+ + ROOH Compound I + ROH Compound I + NADPH NADP· + Compound II Compound II + NADH NAD· + E-Fe3+ NAD· + O2 NAD+ + O2.- Aldehyde oxidase 2R-CHO + 2O2 2R-COOH + O2.- Dihydroorotate dehydrogenase Dihydroorotate + NAD· + O2 NADH + O2.- + Orotic acid
-Fe3+ + ROOH Compound I + ROH. Compound I + NADPH NADP· + Compound II. Compound II + NADH NAD· + E-Fe3+ NAD· + O2 NAD+ + O2.- Aldehyde oxidase. 2R-CHO + 2O2 2R-COOH + O2.- Dihydroorotate dehydrogenase. Dihydroorotate + NAD· + O2 NADH + O2.- + Orotic acid.")
115
Antioksidant defans enzimleri
i. Süperoksit Dismutaz : superoksit uzaklaştırır a. Cu-Zn SOD (eritrosit) b. Mn SOD ( mitokondri) ii. Katalaz : hidrojen peroksit iii. Glutathione-bağımlı Enzimler : peroksit a. Glutathione peroxidase (GPx) and glutathione reductase b. Glutathione-S transferase superfamily iv. Tioredoksin v. Diğer disülfit-bağımlı Enzimler a. Mixed disulfides b. Protein disulfide isomerase vi. Diğer Peroksidazlar
 b. Mn SOD ( mitokondri) ii. Katalaz : hidrojen peroksit. iii. Glutathione-bağımlı Enzimler : peroksit. a. Glutathione peroxidase (GPx) and glutathione reductase. b. Glutathione-S transferase superfamily. iv. Tioredoksin. v. Diğer disülfit-bağımlı Enzimler. a. Mixed disulfides. b. Protein disulfide isomerase. vi. Diğer Peroksidazlar.")
117
Fonksiyonları:İndirgeyici ajan, Konjugayon,aa transportu,Kofaktör, Proteinlerde disülfit bağlarının düzenlenmesi. SH grubu tamponlama gör. (Hb de Fe++)
.")
118
Glutathione Peroxidase :organik hidroperoksitleri katalize eder:
2 GSH + ROOH GSSG + ROH + H2O Glutatyon Peroksidaz yapısında sistein analoğu olan selenosistein vardır. SELENYUM eser elementine gereksinimi vardır.

119
FAGOSİTOZİS’ de ROS lerinin ROLÜ:
HMF O2 NADPH NADPH Oksidaz NADP+ O2.- Lizozom SOD H2O2 Lizozomal enz Cl- Myeloperpksidaz Bakteri HOCl

120
GLUTATYONUN glioksilaz pathway’indeki ROLÜ
Metabolizma süresince bir miktar triozfosfat spontan olarak METİLGLiOKSAL a dönüşür. Reaktif bir dikarbonil şekerdir, proteinlerdeki SH grupları ve guanidin,imidazol ve amino grupları ile reksiyona girer. Proteinlerde çapraz bağlara neden olarak enzimleri inaktive eder. Metilglioksal ayrıca GLİSİN ve ASETON metabolizması süresince de oluşur. GSH bağımlı bir sistem (GLİOKSALAZ PATHWAY) tüm hücrelerde mevcuttur. Metilglioksalı GSH kullanarak laktata dönüştürmektedir.
 tüm hücrelerde mevcuttur. Metilglioksalı GSH kullanarak laktata dönüştürmektedir.")
121
METİLGLİOKSAL’ ın DETOKSİFİKASYONU:
H O H O O O O- GSH H2O C C-SG C-SG C C=O C=O H-C-OH H-C-OH CH CH CH CH3 Glioksilaz 1 GSH Glioksilaz 2 LAKTAT Metilglioksal

Benzer bir sunumlar



















.>")

