Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Redoks Denge Potansiyeli
Serbest Enerji Redoks Denge Potansiyeli BİYOFİZİK A.B.D. Araştırma Gör. Ayhan ÜNLÜ

2
Biyoenerjetik, biyolojik sistemlerde enerji transferini ve kullanımını tanımlar. Termodinamik alanından birkaç temel kuralı, özellikle serbest enerji kavramını temel alır. Serbest enerjideki değişimler (G) kimyasal bir reaksiyonun enerji olarak olabilirliği hakkında bilgi sağlar ve bu şekilde reaksiyonun gerçekleşip gerçekleşemeyeceği hakkında tahminde bulunma olanağı verir.
 kimyasal bir reaksiyonun enerji olarak olabilirliği hakkında bilgi sağlar ve bu şekilde reaksiyonun gerçekleşip gerçekleşemeyeceği hakkında tahminde bulunma olanağı verir.")
3
Serbest enerji kavramı Adenozin trifosfatın (ATP) enerji-veren reaksiyonlardan, enerji-gereken reaksiyonlara enerji taşınmasındaki rolünü anlamak için gereklidir. Biyoenerjetik reaksiyon bileşenlerinin sadece başlangıç ve sonuç enerji durumları ile ilgilenir, mekanizması veya kimyasal değişimin oluşması için ne kadar zaman gerektiği ile ilgilenmez. Kısacası, biyoenerjetik işlemin mümkün olup olmadığını tahmin eder.

4
SERBEST ENERJİ Bir kimyasal reaksiyonun yönü ve ne kadar devam edeceği, reaksiyon sırasında iki faktörün ne derece değişeceğine bağlıdır. Bunlar, entalpi (H, reaksiyona girenlerin ve ürünlerin ısı içeriklerindeki değişimlerin ölçüsüdür) ve entropi (S, reaksiyona girenlerin ve ürünlerin gelişigüzelliği ve düzensizliğindeki değişimin ölçüsüdür. Bu termodinamik birimlerin hiçbirisi kendi başına bir kimyasal reaksiyonun yazıldığı yönde spuntan olarak ilerleyip ilerlemeyeceğini saptamak için yeterli değildir. Ancak, matematiksel olarak birleştirildiklerinde entalpi ve entropi üçüncü bir birimi, serbest reaksiyonunun spontan olarak ilerleyeceği yönü tahmin eden serbest enerjiyi tanımlamak için kullanılabilir.
 ve entropi (S, reaksiyona girenlerin ve ürünlerin gelişigüzelliği ve düzensizliğindeki değişimin ölçüsüdür. Bu termodinamik birimlerin hiçbirisi kendi başına bir kimyasal reaksiyonun yazıldığı yönde spuntan olarak ilerleyip ilerlemeyeceğini saptamak için yeterli değildir. Ancak, matematiksel olarak birleştirildiklerinde entalpi ve entropi üçüncü bir birimi, serbest reaksiyonunun spontan olarak ilerleyeceği yönü tahmin eden serbest enerjiyi tanımlamak için kullanılabilir.")
5
Serbest Enerji Değişimi
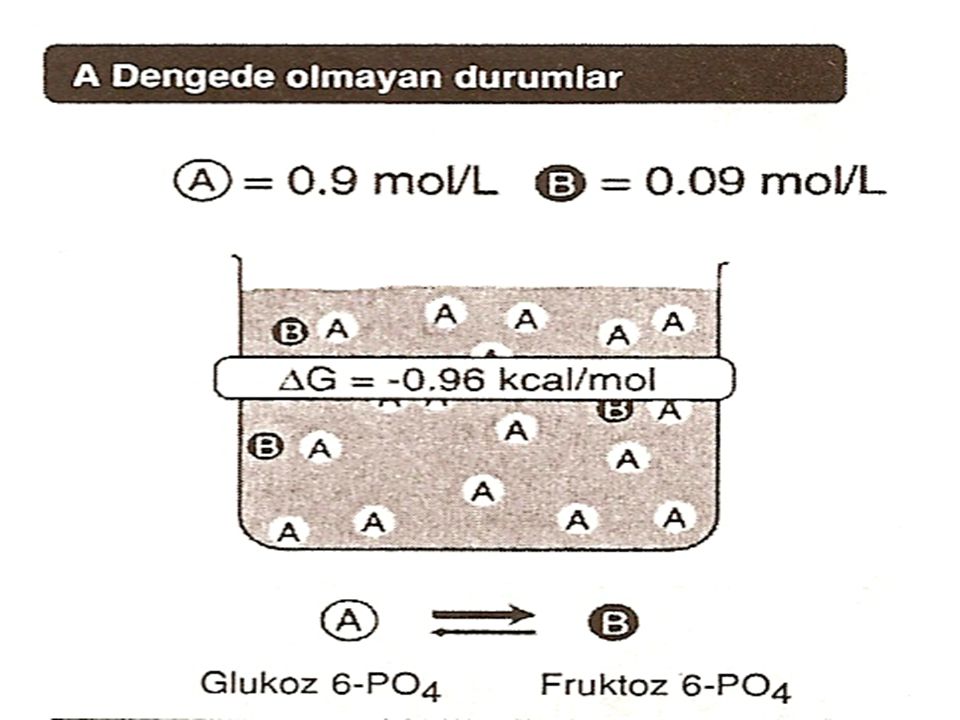
Serbest enerjideki değişim 2 şekilde olabilir. G (Serbest Enerji Değişimi) Daha geneldir. , çünkü serbest enerjideki değişimi ve böylece ürünlerin ve reaktanların belirli konsantrasyonlarında reaksiyonun yönünü tahmin eder. Gº (Standart Serbest Enerji Değişimi) Ürünler ve reaktanlar 1 mol/l iken olan enerji değişimidir. [Standart durumda reaktanlar veya ürünlerin konsantrasyonu 1 mol/l olmasına rağmen, protonların konsantrasyonunun 10-7 mol/l olduğu -ki bu pH= 7 dir- kabul edilir].
 Daha geneldir. , çünkü serbest enerjideki değişimi ve böylece ürünlerin ve reaktanların belirli konsantrasyonlarında reaksiyonun yönünü tahmin eder. Gº (Standart Serbest Enerji Değişimi) Ürünler ve reaktanlar 1 mol/l iken olan enerji değişimidir. [Standart durumda reaktanlar veya ürünlerin konsantrasyonu 1 mol/l olmasına rağmen, protonların konsantrasyonunun 10-7 mol/l olduğu -ki bu pH= 7 dir- kabul edilir].")
7
G işareti reaksiyonun yönünü tahmin eder
Serbes enerjideki değişim, G, sabit bir ısı ve basınçta reaksiyonun yönünün tahmini için kullanılabilir. AB

8
Eğer G negatif bir sayı ise, net bir enerji kaybı vardır ve reaksiyon spontan olarak yazıldığı yönde ilerler yani A, B'ye çevrilir. Reaksiyonun ekzergonik olduğu söylenir.

9
Eğer G pozitif bir sayı ise, net bir enerji kazancı vardır ve reaksiyon spontan olarak B'den A'ya ilerlemez . Reaksiyonun endergonik olduğu söylenir ve reaksiyonun B'den A'ya ilerleyebilmesi için sisteme mutlaka enerji eklemek gerekir. G= O ise, reaktanlar denge halindedir. Bir reaksiyon, spontan olarak ilerliyorsa -yani serbest enerji kaybediliyorsa reaksiyon, G sıfıra ulaşıncaya ve denge kuruluncaya kadar devam eder.

10
İleri ve geri reaksiyonlarda G
İleri doğru olan reaksiyonlarda (AB) serbest enerji, geri doğru olanlara (BA) göre büyüklük olarak eşit, fakat işaret olarak terstir. Örneğin ileri doğru olan reaksiyonlarda G cal/mol ise, geri reaksiyonda cal/mal dür.
 serbest enerji, geri doğru olanlara (BA) göre büyüklük olarak eşit, fakat işaret olarak terstir. Örneğin ileri doğru olan reaksiyonlarda G cal/mol ise, geri reaksiyonda cal/mal dür.")
11
G reaktan ve ürünlerinin konsantrasyonuna bağlıdır
AB reaksiyonunun G'si reaktan ve ürünlerinin konsantrasyonuna bağlıdır. Sabit ısı ve basınçta aşağıdaki eşitlik elde edilebilir.

12
Standart serbest enerji değişimi, Go
G0, standart serbest enerji değişimi olarak isimlendirilir, çünkü reaktan ve ürünlerin 1 mol/l konsantrasyonda ,tutulduğu standart koşullarda serbest enerji değişimine G eşittir. Bu koşullarda ürünlerin reaktanlara oranının doğal logaritması (ln) sıfırdır (In 1 = 0) ve bu nedenle
 sıfırdır (In 1 = 0) ve bu nedenle.")
13
G0 sadece standart koşullarda tahmin edici olur
Standart koşullarda G0 reaksiyonunun devam edeceği yönü tahmin için kullanılabilir, çünkü bu koşullarda G0, G'ye eşittir. Ancak, G0 fizyolojik koşullarda reaksiyonun yönünü tahmin edemez, çünkü sadece sabitelerden meydana gelmiştir (R, T ve Kden) ve bu nedenle ürün veya substrat konsantrasyonlarındaki değişimler ile değiştirilemez.
 ve bu nedenle ürün veya substrat konsantrasyonlarındaki değişimler ile değiştirilemez.")
14
G0 ile Kden arasındaki ilişki
AB reaksiyonunda, daha fazla net kimyasal değişimin meydana gelmediği zaman denge noktasına erişilir. Yani B'nin A'ya çevrildiği hızla A da B'ye çevrilmektedir. Bu durumda [B]'nin [A]'ya oranı, her iki komponentin gerçek konsantrasyonlarından bağımsız olarak, sabittir:

15
burada Kden denge sabitesini, [A]den ve [B]den ise A ve B'nin denge halinde konsantrasyonlarını göstermektedir. Eğer A B reaksiyonunun sabit ısı ve basınçta dengeye gelmesine izin verilirse, toplam serbest enerji değişimi (G) sıfırdır, Bu nedenle
![burada Kden denge sabitesini, [A]den ve [B]den ise A ve B nin denge halinde konsantrasyonlarını göstermektedir.](http://slideplayer.biz.tr/slide/2685860/10/images/15/burada+Kden+denge+sabitesini%2C+%5BA%5Dden+ve+%5BB%5Dden+ise+A+ve+B+nin+denge+halinde+konsantrasyonlar%C4%B1n%C4%B1+g%C3%B6stermektedir..jpg "Eğer A B reaksiyonunun sabit ısı ve basınçta dengeye gelmesine izin verilirse, toplam serbest enerji değişimi (G) sıfırdır, Bu nedenle.")
20
ENERJİ TAŞIYICI OLARAK ATP
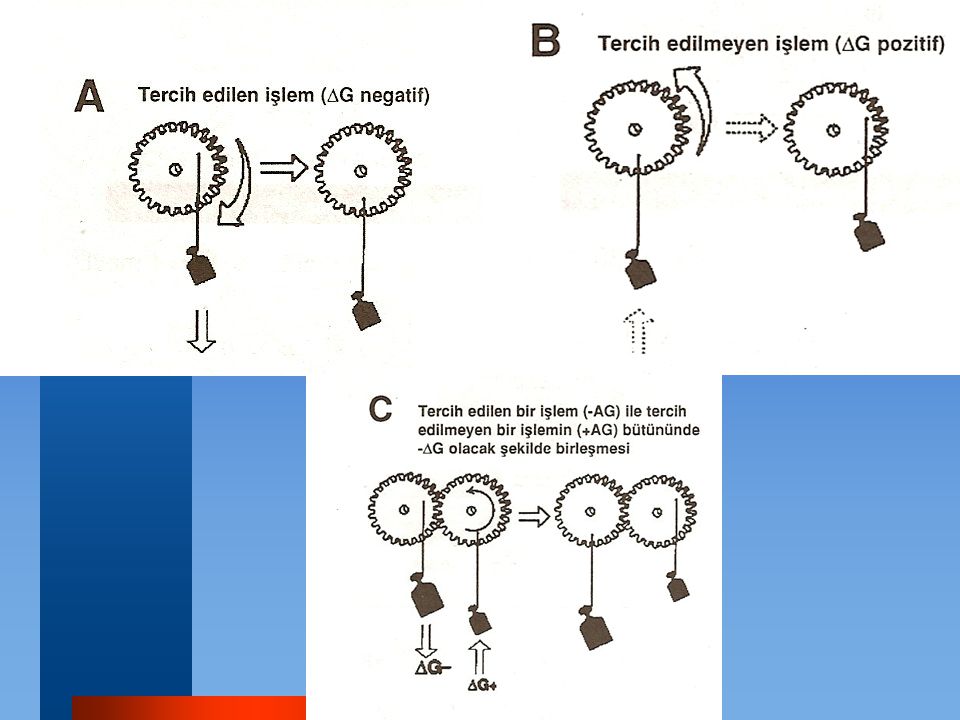
Hücre membranından konsantrasyon farkına karşı geçen iyonların hareketinde olduğu gibi, büyük pozitif G'ye sahip reaksiyon ve işlemler, iyonların endergonik hareketi adenozin trifosfatın (ATP) hidrolizi gibi büyük negatif G 'ye sahip ikinci spontan süreç ile birleştirilerek mümkün hale getirilir
 hidrolizi gibi büyük negatif G ye sahip ikinci spontan süreç ile birleştirilerek mümkün hale getirilir.")
21
ATP ile taşınan enerji ATP, üç fosfat grubunun bir adenozin molekülüne bağlanması ile oluşmuştur. Eğer bir fosfat ayrılırsa adenozin difosfat (ADP), eğer iki fosfat ayrılırsa adenozin monofosfat (AMP) oluşur.
, eğer iki fosfat ayrılırsa adenozin monofosfat (AMP) oluşur.")
22
ATP'nin G0'si ATP hidrolizinin standart serbest enerjisi, G0, terminal fosfat gruplarının herbiri için ortalama cal/mol'dür. Bu büyük, negatif G0 nedeniyle ATP yüksek enerjili fosfat bileşiği olarak adlandırılır.

23
Çok yüksek enerjili fosfat bileşikleri
ATP den daha yüksek enerjiye sahip fosfat içeren bileşikler vardır. Bu çok yüksek enerjili bileşikler fosfoenolpirüvat, 1,3-bifosfogliserat ve fosfokreatin‘i i içerir, ki bunlarda hidrolizin standart serbest enerjisi cal/mol'den büyüktür

24
ELEKTRON TRANSPORT ZİNCİRİ
Glukoz, yağ asidi gibi enerjiden-zengin moleküller sonunda CO2 ve su oluşan bir dizi oksidasyon reaksiyonu ile metabolize olurlar. Bu reaksiyonların metabolik ara ürünleri elekronlarını özelleşmiş koenzimler olan nikotinamid adenin dinükleotid (NAD+) ve flavin adenin dinükleotid (FAD)'e vererek enerjiden-zengin indirgenmiş koenzimler NADH ve FADH2 oluştururlar.
 ve flavin adenin dinükleotid (FAD) e vererek enerjiden-zengin indirgenmiş koenzimler NADH ve FADH2 oluştururlar.")
25
Bu indirgenmiş koenzimler, daha sonra elektron transport zinciri adı verilen bir grup özelleşmiş elektron taşıyıcılarına bir çift elektron verirler. Elektronlar, elektron transport zincirinde ilerledikçe, serbest enerjilerinin büyük bölümünü kaybederler. Bu enerjinin bir bölümü ADP ve Pi'tan ATP sentezlenerek tutulabilir. Bu işlem oksidatit fosforilasyon olarak tanımlanır. ATP olarak yakalanamayan serbest enerjinin kalan bölümü ise ısı olarak salınır.

26
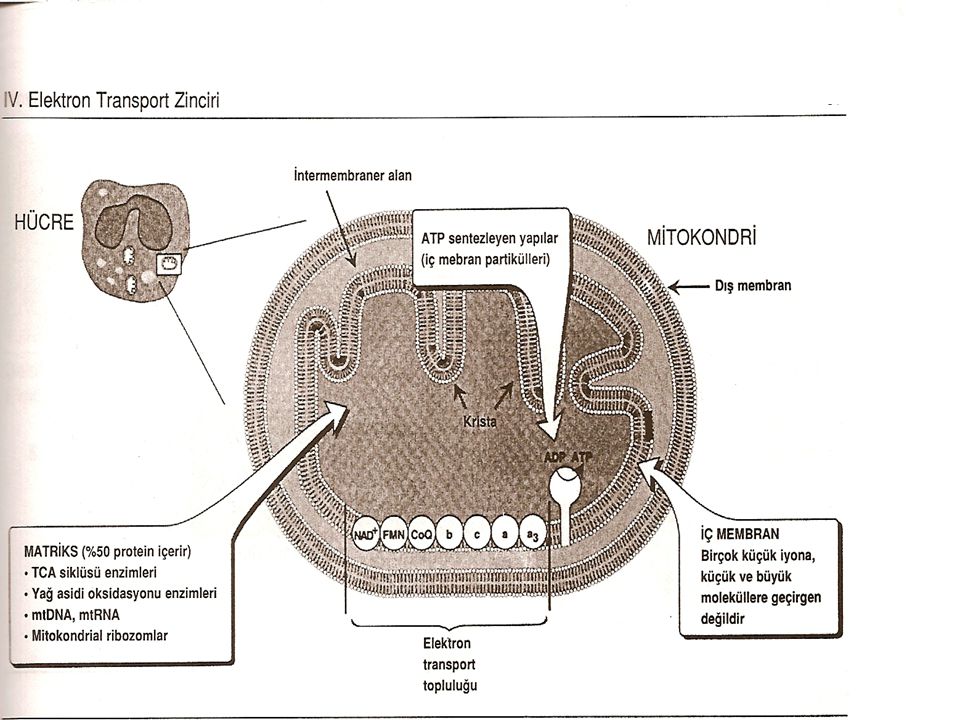
Mitokondri Elektron transport zinciri mitokondri iç zarındadır ve vücutta farklı yakıtlardan elde edilen elektronların oksijene iletilmelerindeki son ortak yoldur. Elektron trasportu ve oksidatif fosforilasyon ile ATP sentezi vücudun mitokondri taşıyan tüm hücrelerinde sürekli oluşur.

27
Mitokondrinin yapısı Elektron transport zincirinin komponentleri iç zarda yerleşmiştir. Dış zar birçok iyonun ve küçük molekülün serbestçe geçişini sağlayan porlar (gözenekler) taşımasına rağmen, mitokondri iç zarı H+, Na+, K+ da dahil olmak üzere küçük iyonların çoğuna, ATP, ADP, pirüvat gibi küçük moleküllere ve mitokondri fonksiyonu için önemli olan diğer metabolitlere geçirgen olmayan özelleşmiş bir yapıdır. iyonların veya moleküllerin bu membrandan geçebilmesi için özelleşmiş taşıyıcılara veya transport sistemlerine gerek vardır. Mitokondri iç zarı alışılmamış şekilde proteinden zengindir ve bunların yarısı elektron transportu ve oksidatif fosforilasyon ile direkt olarak ilgilidir. Mitokondri iç zarı çok kıvrımlıdır Bu kıvrımlara krista adı verilir ve membranın yüzey alanını ileri derecede arttırırlar.
 taşımasına rağmen, mitokondri iç zarı H+, Na+, K+ da dahil olmak üzere küçük iyonların çoğuna, ATP, ADP, pirüvat gibi küçük moleküllere ve mitokondri fonksiyonu için önemli olan diğer metabolitlere geçirgen olmayan özelleşmiş bir yapıdır. iyonların veya moleküllerin bu membrandan geçebilmesi için özelleşmiş taşıyıcılara veya transport sistemlerine gerek vardır. Mitokondri iç zarı alışılmamış şekilde proteinden zengindir ve bunların yarısı elektron transportu ve oksidatif fosforilasyon ile direkt olarak ilgilidir. Mitokondri iç zarı çok kıvrımlıdır Bu kıvrımlara krista adı verilir ve membranın yüzey alanını ileri derecede arttırırlar.")
29
ATP sentetaz kompleksleri
Bu protein kompleksleri iç membran partikülleri olarak adlandırılırlar ve mitokondri iç zarının iç yüzeyine bağlanmışlardır. Mitokondri matriksine doğru uzanan küreler şeklinde görülürler.

30
Mitokondri matriksi Mitokondri içindeki jel-kıvamındaki bu soIüsyon pirüvat, amino asitler ve yağ asitlerinin oksidasyonundan sorumlu enzimleri ve trikarboksilik asid (TCA) siklusu enzimlerini içerir. Üre ve hem sentezinin bir bölümü mitokondri matriksinde gerçekleşir. Ayrıca ek olarak NAD+ ve FAD (hidrojen alıcısı olarak gerekli olan koenzimlerin yükseltgenmiş şekilleri ) ile ATP oluşumunda gerekli olan ADP ve Pi içerir.
 siklusu enzimlerini içerir. Üre ve hem sentezinin bir bölümü mitokondri matriksinde gerçekleşir. Ayrıca ek olarak NAD+ ve FAD (hidrojen alıcısı olarak gerekli olan koenzimlerin yükseltgenmiş şekilleri ) ile ATP oluşumunda gerekli olan ADP ve Pi içerir.")
32
Zincirin organizasyonu
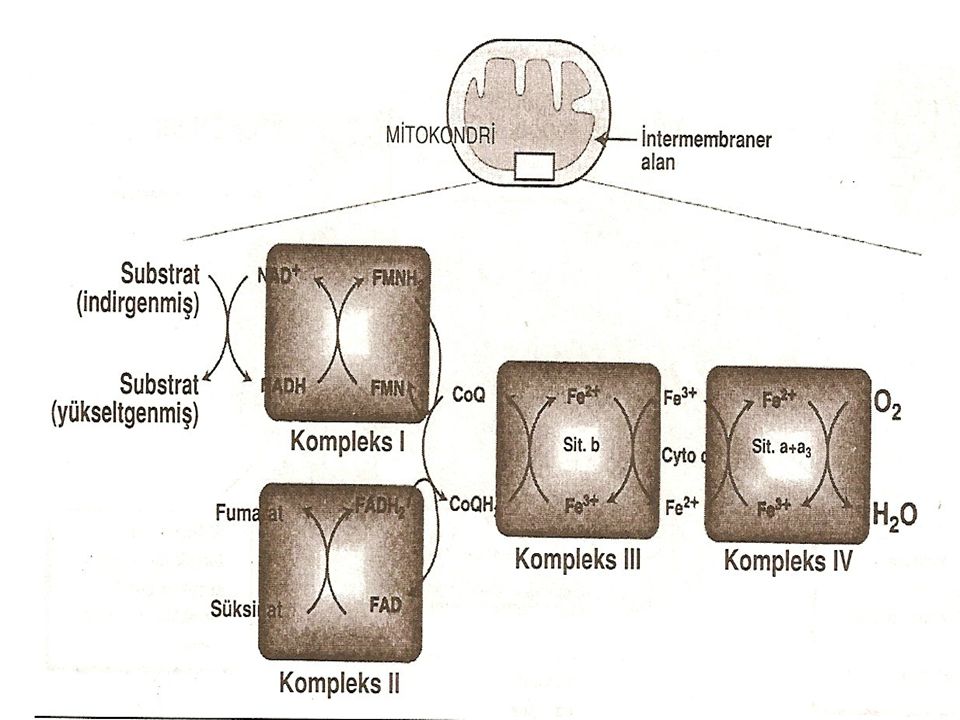
Mitokondri iç zarı beş ayrı enzim kompleksine parçalanabilir ve bunlara kompleks I, II, III, IV ve V adı verilir. Kompleks I-IV'ün her birisi elektron transport zincirinin bir bölümünü içerir. Kompleks V ise ATP sentezini katalize eder. Her kompleks elektronlarını koenzim Q ve sitokom C gibi hareketli elektron taşıyıcılarına verir. Elektron transport zincirinin her taşıyıcısı bir elektron vericisinden elektronları alır ve daha sonra en sonunda su oluşturmak için oksijen ve protonlarla birleşmek üzere zincirdeki bir sonraki taşıyıcıya verir. Elektron transport işlemini solunum zinciri yapan bu oksijen gereksinimidir ve vücudun oksijen tüketiminin en büyük bölümü burada gerçekleşir.

34
Elektron transport zinciri reaksiyonları
Koenzim Q dışında bu zincirin tüm üyeleri proteindir. Bunlar bazı dehidrogenazlarda olduğu gibi enzim olarak işlev yapabilirler, demir-kükürt merkezinin bir bölümü olarak demir taşıyabilirler, sitokromlarda olduğu gibi bir porfirin halkası ile koordine olabilirler veya sitokrom a+a3 de olduğu gibi bakır taşıyabilirler.

35
NADH oluşumu NAD+, substratlarından iki hidrojen atomu Çıkaran dehidrogenazlar tarafından NADH'a indirgenir. Her iki elektron, fakat sadece bir NADH ve ortama salınan bir serbest proton, H+ oluşturmak üzere NAD+ ye aktarılır.

36
NADH dehidrogenaz Serbest proton ve NADH tarafından taşınan hidrid iyonu daha sonra mitokondri iç zarında yerleşmiş bir enzim kompleksi olan NADH dehidrogenaza aktarılır. Bu kompleks kendisine sıkıca bağlanmış bir molekül flavin mononükleotid içerir ve bu koenzim iki hidrojen atomunu (2e+2H+) alarak FMNH2 haline gelir. NADH dehidrogenaz ayrıca demir-kükürt merkezleri oluşturmak üzere kükürt atomları ile çiftler oluşturmuş birkaç demir atomu da içerir. Bunlar hidrojen atomlarının zincirin bir sonraki üyesi olan ubikinona (koenzim Q olarak bilinir) aktarılması için gereklidir.
 alarak FMNH2 haline gelir. NADH dehidrogenaz ayrıca demir-kükürt merkezleri oluşturmak üzere kükürt atomları ile çiftler oluşturmuş birkaç demir atomu da içerir. Bunlar hidrojen atomlarının zincirin bir sonraki üyesi olan ubikinona (koenzim Q olarak bilinir) aktarılması için gereklidir.")
37
Koenzim Q Koenzim Q, uzun bir izoprenoid kuyruğu olan bir kinon türevidir. Biyolojik sistemlerde çok yaygın olarak bulunduğundan (ubiquitous) ubikinon olarak da adlandırılır. Koenzim Q, hidrojenlerini hem NADH dehidrogenaz tarafından oluşturulmuş FMNH2 den hem de süksinat dehidrogenaz ve asil CoA dehidrogenaz tarafından oluşturulmuş FADH2'den alabilir.
 ubikinon olarak da adlandırılır. Koenzim Q, hidrojenlerini hem NADH dehidrogenaz tarafından oluşturulmuş FMNH2 den hem de süksinat dehidrogenaz ve asil CoA dehidrogenaz tarafından oluşturulmuş FADH2 den alabilir.")
38
Sitokromlar Elektron transport zincirinin geri kalan üyeleri sitokromlardır. Her biri bir atom demir taşıyan porfirin halkasından oluşmuş bir hem grubu içerirler. Hemoglobinin hem grubundan farklı olarak sitokromun demir atomu, reversibi elektron taşıyıcı görevinin gereği olarak reversibi olarak ferrik (Fe+3) şeklinden ferro (Fe+2) şekline dönüşür. Elektronlar zincirde koenzim Q'dan-sitokrom b. c ve a+a3'e doğru ilerlerler
 şeklinden ferro (Fe+2) şekline dönüşür. Elektronlar zincirde koenzim Q dan-sitokrom b. c ve a+a3 e doğru ilerlerler.")
39
Sitokrom a+a3 Bu sitokrom hem halkasının demirinin moleküler oksijen ile direkt olarak reaksiyona girebileceği bir serbest bağ yapısına sahip olan tek elektron taşıyıcısıdır. Bu bölgede, taşınmış elektronlar, moleküler oksijen ve serbest protonlar su oluşturmak üzere bir araya getirilirler. Sitokrom a+a3 (sitokrom oksidaz da denir) bu kompleks reaksiyonun gerçekleşebilmesi için gerekli olan bağlı bakır atomları da içerir.
 bu kompleks reaksiyonun gerçekleşebilmesi için gerekli olan bağlı bakır atomları da içerir.")
41
inhibitörler Bu bileşikler zincirin bir komponentine bağlanıp oksidasyon / redüksiyon reaksiyonunu engelleyerek elektron geçişini önlerler. Bu nedenle bu engelin gerisindeki elektron taşıyıcıları tamamen indirgenmiş, ilerisindekiler ise yükseltgen'miş şekilde bulunurlar.

42
Elektron transportu sırasında serbest enerji salınımı
Elektronlar, elektron transport zinciri boyunca bir elektron vericisinden (indirgeyici ajan veya redüktan) bir'elektron alıcısına (yükseltgeyici ajan veya oksidan) aktarıldıkça serbest enerji salınır. Elektronlar değişik şekillerde aktarılabilir.
 bir elektron alıcısına (yükseltgeyici ajan veya oksidan) aktarıldıkça serbest enerji salınır. Elektronlar değişik şekillerde aktarılabilir.")
43
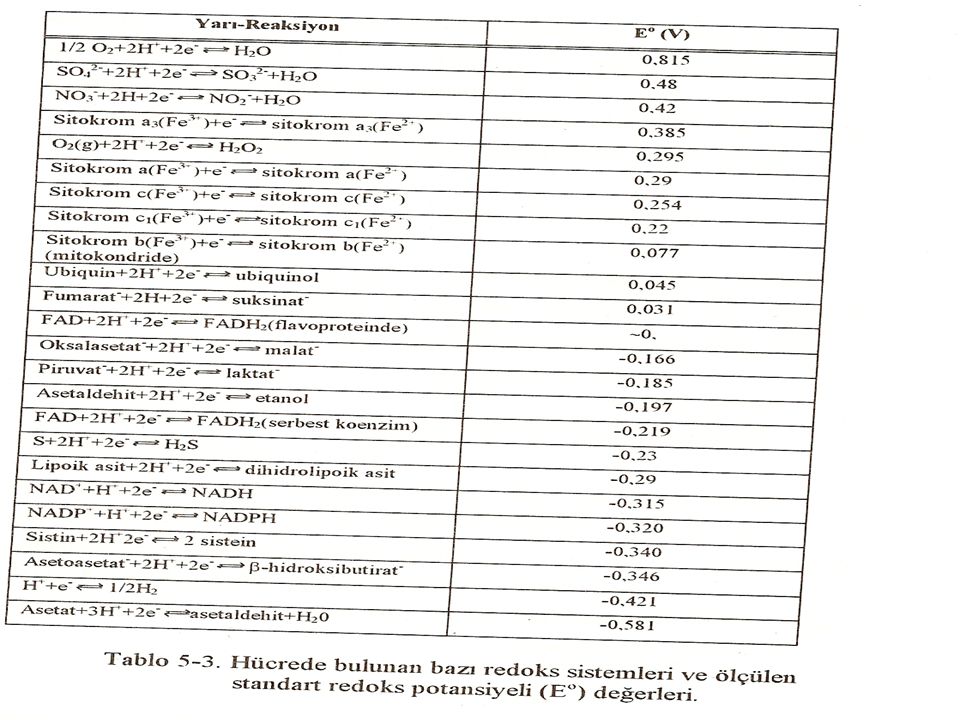
Redoks çiftleri Bir bileşiğin oksidasyonuna (elektron kaybı) mutlaka ikinci bir maddenin redüksiyonu (elektron kazanımı) eşlik eder. Örneğin, Şekilde NADH'ın NAD+'ye oksidasyonuna FAD'nin FADH2'ye redüksiyonunun eşlik ettiğini göstermektedir. Böyle oksidasyon-redüksiyon reaksiyonları iki reaksiyonun toplamı olarak yazılabilir: bir izole oksidasyon reaksiyonu ve ayrı bir redüksiyon reaksiyonu NAD+ ve NADH ile FAD ve FADH2 redoks çiftlerini oluştururlar. Redoks çiftleri elektron kaybetme eğilimlerinde farklılık gösterirler. Bu eğilim o redoks çiftinin özelliğidir ve kantitatif olarak birimi volt olan Eo sabitesi (standart redüksiyon potansiyeli) ile tanımlanır.
 mutlaka ikinci bir maddenin redüksiyonu (elektron kazanımı) eşlik eder. Örneğin, Şekilde NADH ın NAD+ ye oksidasyonuna FAD nin FADH2 ye redüksiyonunun eşlik ettiğini göstermektedir. Böyle oksidasyon-redüksiyon reaksiyonları iki reaksiyonun toplamı olarak yazılabilir: bir izole oksidasyon reaksiyonu ve ayrı bir redüksiyon reaksiyonu NAD+ ve NADH ile FAD ve FADH2 redoks çiftlerini oluştururlar. Redoks çiftleri elektron kaybetme eğilimlerinde farklılık gösterirler. Bu eğilim o redoks çiftinin özelliğidir ve kantitatif olarak birimi volt olan Eo sabitesi (standart redüksiyon potansiyeli) ile tanımlanır.")
44
Standart redüksiyon potansiyeli (Eo)
Çeşitli redoks çiftlerinin redüksiyon potansiyelleri en negatif Eo'dan en pozitife kadar sıralanabilir. Bir redoks çiftinin standart redüksiyon potansiyeli ne kadar daha fazla negatif ise o çiftin redüktan üyesinin elektron kaybetme eğilimi o kadar fazladır. Eo ne kadar daha fazla pozitif ise o çiftin oksidan üyesinin elektron kabul etme eğilimi o kadar fazladır. Bu nedenle elektronlar daha negatif Eo'a sahip çiftten daha pozitif Eo'a sahip çifte doğru kayarlar.

47
ATP'nin G0'ı ATP'nin terminal fosfat grubunun hidrolizinin standart serbest enerjisi cal/mol'dür. Bir çift elektronun NADH'dan elektron transport zinciri yolu ile oksijene taşınması cal üretir ve bu nedenle 3 ADP ve 3 Pi'dan 3 ATP oluşumuna yetecek kadar enerji sağlar (3x7,300= 21,900 cal) artan kaloriler ısı olarak salınır. [Not: Bir çift elektronun elektron transport zinciri yolu ile FADH2 veya FMNH2 den oksijene taşınması 2ADP ve 2Pi'dan 2 ATP oluşumuna yetecek kadar enerji üretir.]
 artan kaloriler ısı olarak salınır. [Not: Bir çift elektronun elektron transport zinciri yolu ile FADH2 veya FMNH2 den oksijene taşınması 2ADP ve 2Pi dan 2 ATP oluşumuna yetecek kadar enerji üretir.]")
48
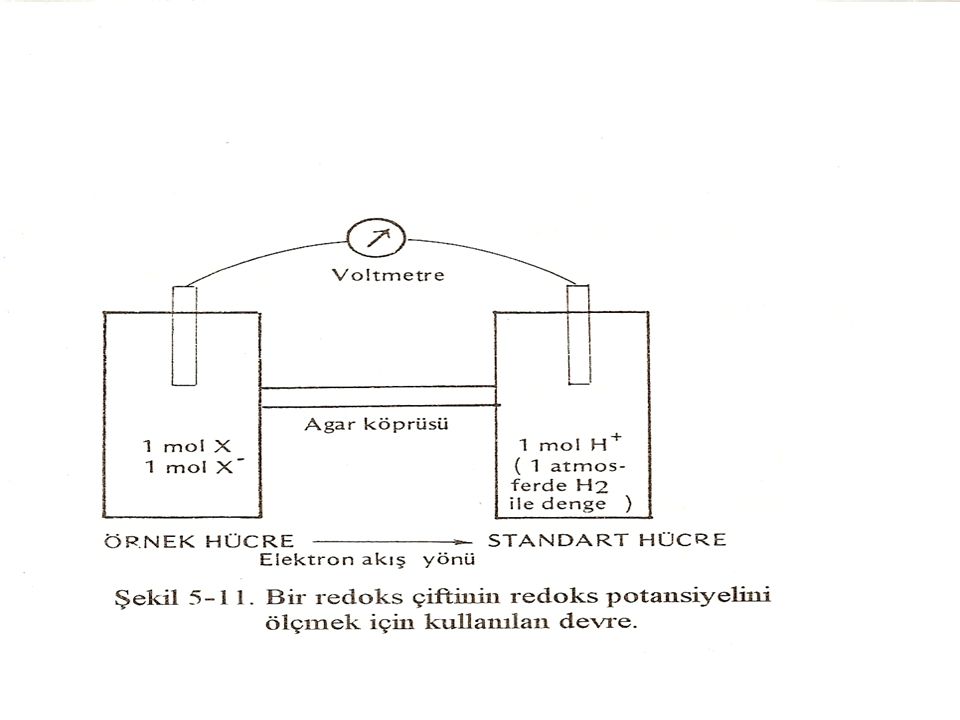
Redoks potansiyeli ve ölçülmesi
Bir maddenin O2 ile birleşmesi, ya da hidrojen (elektron) yitirmesi oksitlenme (yükseltgenme) bunun tersi ise indirgenme (redüklenme) olarak tanımlanır. O2, oksitleyici bir madde olarak hücre solunumu ile H20'ye indirgenir. Çok zayıf bir oksitleyici olan H+ ise çok güçlü bir indirgeyici olan H2 ye dönüşebilir. O2 – H2O ve H+ - H2 iki önemli redoks çiftini oluşturmaktadır. Genelde, bir maddenin oksitlenmiş (X) ya da indirgenmiş (X-) biçimleri, (X) ve (X-) çifti, redoks çifti olarak tanımlanır. Hücre içinde metabolik önemi olan birçok redoks çifti (sistemi)vardır'
 yitirmesi oksitlenme (yükseltgenme) bunun tersi ise indirgenme (redüklenme) olarak tanımlanır. O2, oksitleyici bir madde olarak hücre solunumu ile H20 ye indirgenir. Çok zayıf bir oksitleyici olan H+ ise çok güçlü bir indirgeyici olan H2 ye dönüşebilir. O2 – H2O ve H+ - H2 iki önemli redoks çiftini oluşturmaktadır. Genelde, bir maddenin oksitlenmiş (X) ya da indirgenmiş (X-) biçimleri, (X) ve (X-) çifti, redoks çifti olarak tanımlanır. Hücre içinde metabolik önemi olan birçok redoks çifti (sistemi)vardır")
49
Fe (demir) ve Cu (bakır) redoks çiftleri olup Fe ' den 1 elektron Cu ' ya geçmiştir.Redoks çiftleri arasında elektron alıp verme potansiyeline " Redoks potansiyeli " denir.Elektron transfer zincirinde ise redoks çiftleri, potansiyeli küçük olandan büyük olana doğru sıralanmıştır.Böylelikle elektron seri bir şekilde akmaktadır.Redoks çiftleri ile elektron akımı, bitkilerin kloroplastlarındaki " Sitokrom " moleküllerinde de görülür.ATP ise, elektronun bir redoks çiftinden diğerine geçişi sırasında sentezlenir.
 ve Cu (bakır) redoks çiftleri olup Fe den 1 elektron Cu ya geçmiştir.Redoks çiftleri arasında elektron alıp verme potansiyeline Redoks potansiyeli denir.Elektron transfer zincirinde ise redoks çiftleri, potansiyeli küçük olandan büyük olana doğru sıralanmıştır.Böylelikle elektron seri bir şekilde akmaktadır.Redoks çiftleri ile elektron akımı, bitkilerin kloroplastlarındaki Sitokrom moleküllerinde de görülür.ATP ise, elektronun bir redoks çiftinden diğerine geçişi sırasında sentezlenir.")
Benzer bir sunumlar

















>")








