Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Membran Proteinleri Ayhan ÜNLÜ

2
Hücre zarı ya da plazma zarı hücrenin organellerini ve sıvı içeriğini saran ve hücreye yapısal bütünlük sağlayan ayrıca çok önemli işlevleri yürüten seçici geçirgen bir yapıdır.

3
Yapısı: Hücre zarı baslıca protein ve fosfolipitlerden olusmus çift katlı bir sıvıdır. Fosfolipit tabaka membranın sıvı bölümünü olustururken, fosfolipitten olusmus bu sıvı tabaka içine gömülü halde bulunan proteinler ise mozaik bölümünü olustururlar.

4
Fosfolipit tabakası birbirine sırt sırta dayanmış molekül gruplarından oluşmaktadır. Kolesterol molekülleri protein-lipit tabakadaki sırt sırta vermiş molekülleri birbirine bağlamada iş görür. Zar proteinleri ise ya iyon transferinde yada karbonhidratlara yataklık etmede görev alır. Şekilde mavi renkli yapılar zar proteinleri olup içindeki kanallar zarın seçici geçirgen özelliğini belirler.

5
b) ÖZELLİKLERİ: Canlı-saydam ve esnektir Üzerinde por denilen delikler bulunur Seçici geçirgendir Zardaki proteinler enzim görevi yapar Akışkan bir yapıya sahiptir c)GÖREVLERİ: Hücreyi dağılmaktan korur Hücreyi dış etkilerden korur Madde alış-verişini sağlar
GÖREVLERİ: Hücreyi dağılmaktan korur. Hücreyi dış etkilerden korur. Madde alış-verişini sağlar.")
6
Yaklaşık olarak 6-10 nanometre kalınlığında olan hücre zarı, farklı hücre tiplerinde, değişik oranlarda lipid. protein ve karbonhidrat içerir. Hücre protein oranının fazla olması, hücrenin biyokimyasal etkinliğinin çok olduğunun bir göstergesidir. Zara yerleşik proteinler; almaç (reseptör-hormonlar ya da 'ileticiler' hücre yüzeyinde bulunan bu yapılara bağlanarak hücresel tepkimeleri başlatabilirler), enzim ya da zarın her iki yönüne geçişi düzenleyen bir kanal işlevi görürler. Bu bağlamda yüksek oranda protein içeren bir zarın işlevlerinin de o oranda fazla olduğu düşünülmektedir.
, enzim ya da zarın her iki yönüne geçişi düzenleyen bir kanal işlevi görürler. Bu bağlamda yüksek oranda protein içeren bir zarın işlevlerinin de o oranda fazla olduğu düşünülmektedir..")
7
Zar Lipidleri Hücre zarı iki tabakalı bir fosfolipid yapıdan oluşur. Her bir fosfolipid tabaka, hidrofilik (suyu seven) baş ve hidrofobik (suyu sevmeyen) kuyruk içerir. Fosfolipid tabakanın hidrofilik baş kısımları, zarın dış tabakasını oluştururken; hidrofobik kuyruk kısımları, diğer fosfolipid tabakanın hidrofobik bölümüyle karşılıklı durarak çift tabakalı hücre mebramnın iç kısımlarını oluştururlar.
 baş ve hidrofobik (suyu sevmeyen) kuyruk içerir. Fosfolipid tabakanın hidrofilik baş kısımları, zarın dış tabakasını oluştururken; hidrofobik kuyruk kısımları, diğer fosfolipid tabakanın hidrofobik bölümüyle karşılıklı durarak çift tabakalı hücre mebramnın iç kısımlarını oluştururlar.")
8
Zar lipidleri, büyük oranda fosfolipidlerden oluşsa da
Zar lipidleri, büyük oranda fosfolipidlerden oluşsa da. daha az oranda kolesterol ve diğer lipid tiplerini de içerir. Zarda bulunan kolesterolün en önemli görevi, yüksek ısı gibi iç ya da dış etkenlerin zar akışkanlığını artırmasını engelleyerek, zar dayanıklılığının sürdürülmesini sağlamaktır.

9
Zar Hareketleri Hücre zarı hareketsiz ve sabit bir yapı olmayıp, yapım ve yıkımla beraber, kendine özgü hareketin de sürekli olarak izlendiği özel bir organeldir. Zar hareketleri, her iki yana, kendi çevresinde ve boylamasına olabilir.

10
Zar Proteinleri Her ne kadar biyolojik zarların temel yapısı lipidler tarafından oluşturuIuyorsa da, hücrenin yaşamasıyla ilgili birçok özel fonksiyon proteinler tarafından gerçekleştirilir. Zarda yer alan proteinlerin tipleri ve miktarları oldukça farklıdır. Örneğin; Sinir aksonlarını kuşatan myelin zarlarda, zarın toplam kütlesinin % 25 inden daha azı proteinken, mitokondri, kloroplast gibi enerji üreten yapılarda bu miktar % 75 kadardır. Olağan zarlarda ise bu miktar yaklaşık % 50 dolaylarındadır. Lipid molekülleri, protein molekülleriyle kıyaslandığında, hacım olarak daha küçük olduğundan bu % 50 lik oranda bile çok fazla sayıda lipid molekülü vardır. Diğer bir deyişle zarlarda 1 protein molekülüne karşılık yaklaşık 50 lipid molekülü yer alır.

11
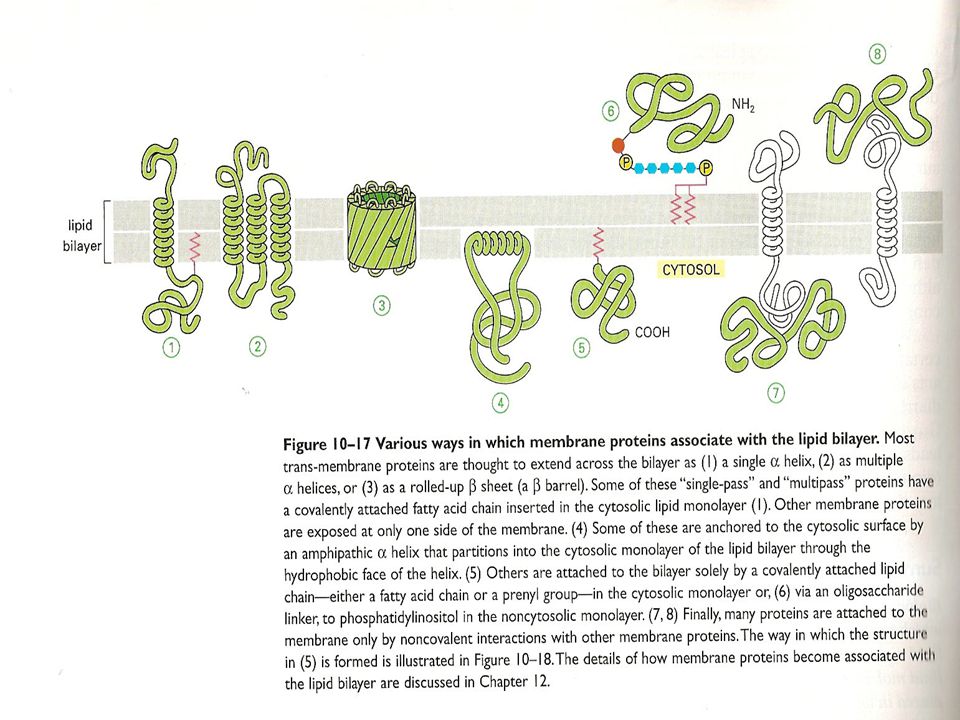
Proteinlerin Zarlardaks Yerleşim Şekilleri
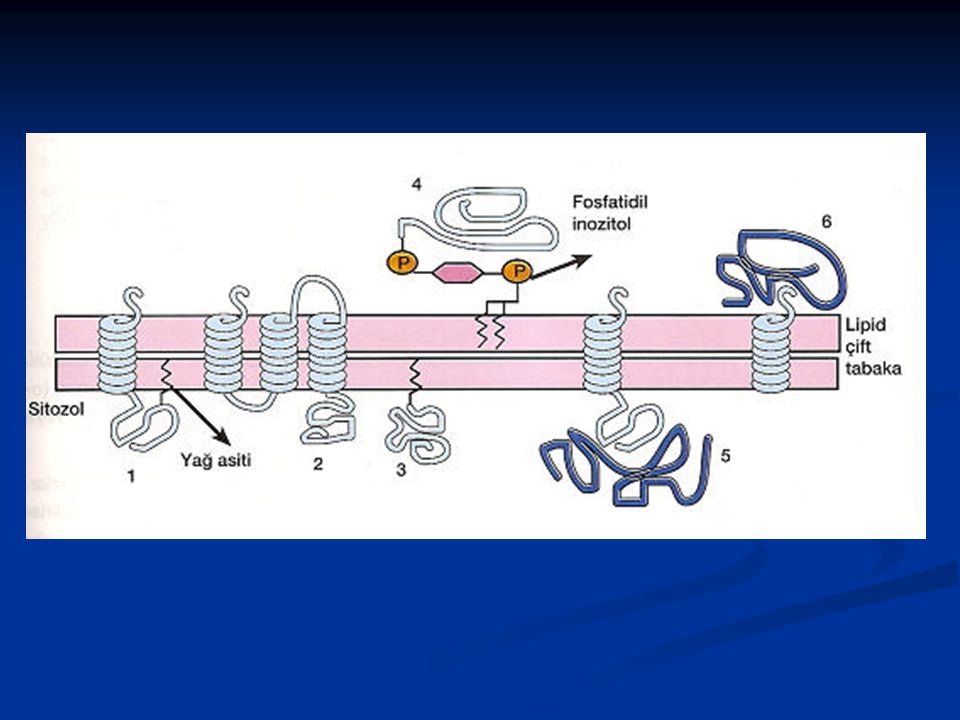
Birçok zar proteini, lipid tabakada bir taraftan diğer tarafa kadar uzanır. Transmembran proteinler olarak adlandırılan bu proteinler amfipatik özelliktedir, yani hem hidrofilik hem de hidrofobik bölgelere sahiptirler. Hidrofobik kısımları lipid tabakanın hidrofobik iç kısmıyla, hidrofilik kısımları ise lipid tabakanın her iki yanında yer alan su tabakasıyla ilişkidedir. Bu proteinlerin bazılarının hidrofobikliği kendilerine kovalent olarak bağlanmış olan ve lipid tabakanın sitoplazmik yarısı içine gömülü bulunan yağ asiti zincirleri nedeniyle artabilir. Zarların sitoplazmik yarısına bağlanan bazı proteinler sadece yağ asiti zincirleri aracılığıyla lipid tabakaya tutunur . Bazı hücre yüzey proteinleri ise küçük bir fosfolipid olan fosfatidil inositole kovalent olarak bağlanarak lipid tabakanın dış yüzünde yer alırlar

13
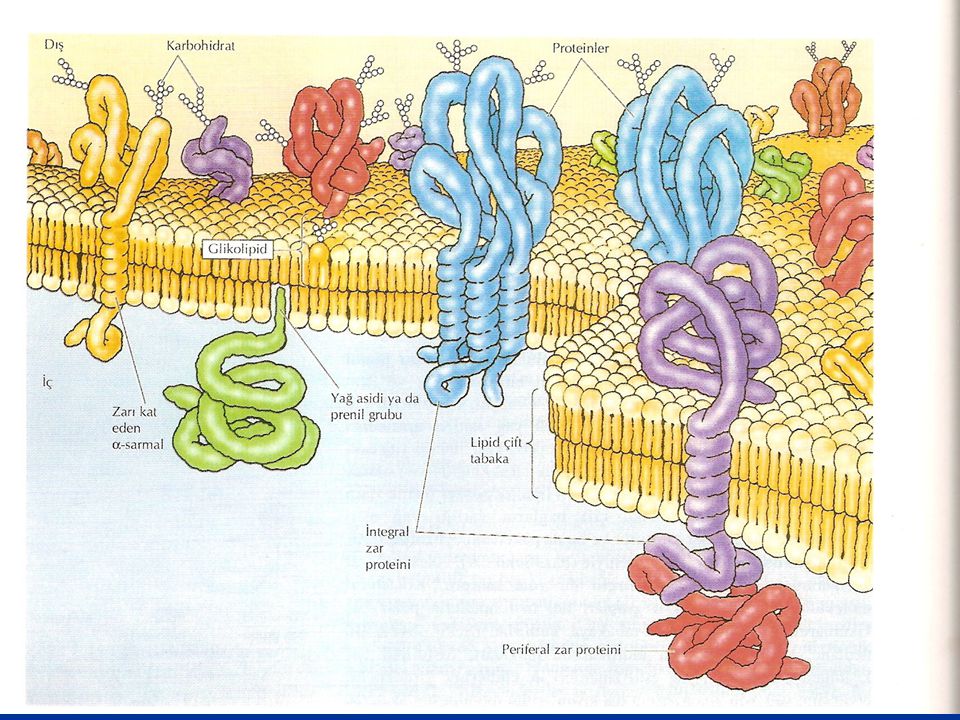
Bunların dışındaki proteinler ise zarın her hangi bir yüzeyinde yer alıp, hidrofobik bölgeye kadar uzanmazlar, zarda yer alan diğer proteinler ile nonkovalent bağlarla bağlanmış olarak bulunurlar. Bunlar ekstraksiyon metodlarıyla zarlardan çok kolay ekstrakte edilebilirler. Bu tip proteinlere periferal zar proteinleri adı verilir. Bunların aksine, transmembran proteinler, fosfatidil inositol'e bağlı proteinler ve yağ asitlerine bağlı proteinler bütün lipid tabaka deterjanlarla veya organik çözücülerle tahrip edilmek suretiyle elde edilebilirler. O nedenle transmembran proteinlere integral zar proteinleri adı da verilir.

14
Zar Proteinleri İntegral zar proteinleri Periferal zar proteinleri Hücre zarında çok farklı işleve sahip proteinler bulunur. Zar proteinlerini zardaki yerleşimlerine göre, "integral" ve "periferal" olarak ikiye ayırmak mümkün. İntegral proteinler, zarın her iki tarafına doğru yerleşim gösterirken, periferal proteinler, ya iç ya da dış tabakada bulunurlar.

15
Zar Proteinleri 1. İntegral zar proteinleri
a. Transmembran zar proteinleri b. Zarın sitozolik yüzüne bağlanan zar proteinleri c. Zarın dış yüzüne bağlanan zar proteinleri 2. Periferal zar proteinleri

16
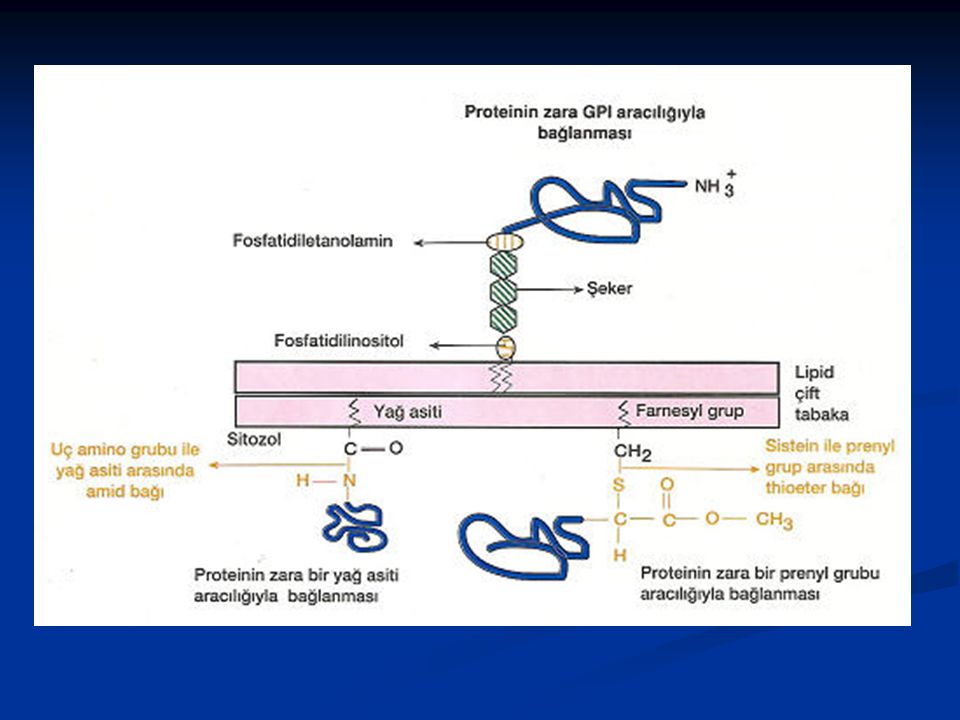
Yağ asitlerine kovalent olarak tutunan proteinler zarda bulunan lipidlere kovalent olarak bağlanır. Bu proteinler üç tip lipid grubuyla bağlanırlar. Birinci tip bağlanmada (açilasyon); bir yağ asiti zinciri (myristic veya palmitic acid) bir amid bağı aracılığıyla, sitozolik yüzde, proteinin amino ucundaki glisine kovalent olarak bağlanır. İkinci tip bağlanmada (prenilasyon); bir prenyl grubu (farnesyl veya daha uzun olan geranyigeranyl grubu) bir thioeter bağı ile, sitozolik yüzde, proteinin karboksil ucundaki 4. sırada yer alan sistein'e bağlanır. Bu prenilasyonu takiben uçtaki 3 amino asit ayrılır ve yeni oluşan karboksil uç metillenir, böylece protein zara yerleşmiş olur. Üçüncü tip bağlanmada; proteinler, zarın dış yüzünde yer alan glikozilfosfatidilinositol (glycosylphosphatidylinositol)(GPI)'a bağlanır. GPl'ın yapısı hücreler arasında farklılık gösterebilir. Bu yapı genel olarak fosfatidilinositol ve fosfatidiletanoamin ihtiva eder. Fosfatidilinositolün iki yağ asidi zarda uzanırken fosfatidiletanoamin bu lipid yapıyı proteine bağlar. Fosfatidilinositol ve fosfatidiletanoamin arasında ise sayısı hücre tipine göre değişen şeker rezüdüler yer alır.
; bir yağ asiti zinciri (myristic veya palmitic acid) bir amid bağı aracılığıyla, sitozolik yüzde, proteinin amino ucundaki glisine kovalent olarak bağlanır. İkinci tip bağlanmada (prenilasyon); bir prenyl grubu (farnesyl veya daha uzun olan geranyigeranyl grubu) bir thioeter bağı ile, sitozolik yüzde, proteinin karboksil ucundaki 4. sırada yer alan sistein e bağlanır. Bu prenilasyonu takiben uçtaki 3 amino asit ayrılır ve yeni oluşan karboksil uç metillenir, böylece protein zara yerleşmiş olur. Üçüncü tip bağlanmada; proteinler, zarın dış yüzünde yer alan glikozilfosfatidilinositol (glycosylphosphatidylinositol)(GPI) a bağlanır. GPl ın yapısı hücreler arasında farklılık gösterebilir. Bu yapı genel olarak fosfatidilinositol ve fosfatidiletanoamin ihtiva eder. Fosfatidilinositolün iki yağ asidi zarda uzanırken fosfatidiletanoamin bu lipid yapıyı proteine bağlar. Fosfatidilinositol ve fosfatidiletanoamin arasında ise sayısı hücre tipine göre değişen şeker rezüdüler yer alır.")
18
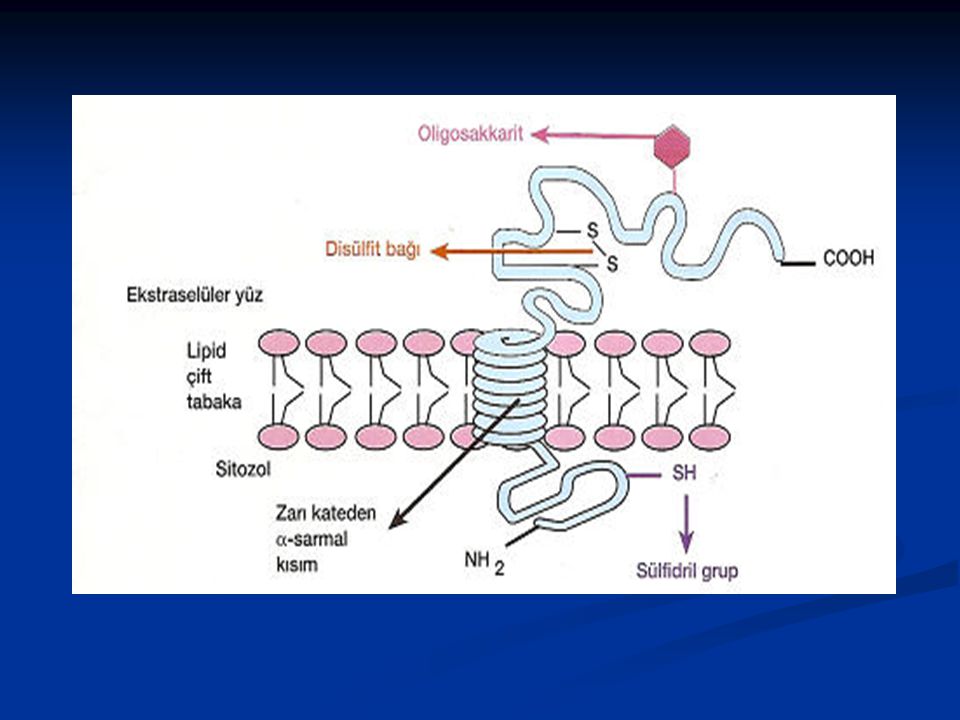
Transmembran proteinlerin, polipeptid zincirinin lipid çift tabakanın hidrofobik bölgesine gömülü olan kısmı nonpolar yan zincirli amino asitlerden oluşurlar. Bu proteinler, lipid çift tabakayı α-sarmal (α -heliks) halinde geçerler . Bu geçiş ya tek bir zincir şeklinde, tek geçişli (single-pass), veya birkaç tane α -sarmal zincir halinde, çok geçişli multipass olur . Transmembran proteinlerin çoğu glikozillenmiştir (şeker grupları eklenmiştir). Şeker molekülleri bu proteinlere endoplazmik retikulum ve Golgi Kompleksinde eklenir ve proteinlerin oligosakkarit zincirleri daima ekstraselüler yüzde yer alır. Proteinlerin sergiledikleri asimetri, sahip oldukları diğer ilave gruplar ve çeşitli bağlar tarafından daha da artırılır. Bunlardan, sülfidril (sulphydril = SH) grupları sitoplazmik yüzde, disülfit (S-S) bağları ekstraselüler yüzde yer alır
. Şeker molekülleri bu proteinlere endoplazmik retikulum ve Golgi Kompleksinde eklenir ve proteinlerin oligosakkarit zincirleri daima ekstraselüler yüzde yer alır. Proteinlerin sergiledikleri asimetri, sahip oldukları diğer ilave gruplar ve çeşitli bağlar tarafından daha da artırılır. Bunlardan, sülfidril (sulphydril = SH) grupları sitoplazmik yüzde, disülfit (S-S) bağları ekstraselüler yüzde yer alır.")
20
Spektrin (Specirin) Eritrositlerdeki protein moleküllerinin çoğu lipid tabakanın sitoplazmik yüzüne tutunmuş olan periferal zar proteinleridir. Bunların içinde de en bol bulunanı spektrin'dir. Spektrin yaklaşık 100 nm uzunlukta, uzun, ince, esnek, çubuk şeklinde olup zar proteinlerinin yaklaşık % 25 ini teşkil eder. Zarın altında yer alan sitoplazma iskeletinin esas elemanıdır. Spektrin iki büyük polipeptid zincirinden meydana gelmiştir. Bunlar: α-spektrin (~ dalton) ve β-spektrin (~ dalton) zincirlerdir. Esnek yapıdaki bu zincirler birbirlerine birkaç noktada nonkovalent olarak tutunmuştur. Spektrin heterodimerleri uç uca eklenerek 200 nm uzunlukta tetramerleri oluşturur, α ve β zincirler 106 amino asit uzunluğunda, çok sayıda tekrarlayan kısımlardan meydana gelmiştir.
 ve β-spektrin (~ dalton) zincirlerdir. Esnek yapıdaki bu zincirler birbirlerine birkaç noktada nonkovalent olarak tutunmuştur. Spektrin heterodimerleri uç uca eklenerek 200 nm uzunlukta tetramerleri oluşturur, α ve β zincirler 106 amino asit uzunluğunda, çok sayıda tekrarlayan kısımlardan meydana gelmiştir.")
21
Birkaç tane spektrin tetrameri, kısa bir aktin filament ve band 4
Birkaç tane spektrin tetrameri, kısa bir aktin filament ve band 4.1 adlı başka bir protein aracılığıyla birbirlerine bağlanarak zarın sitoplazmik yüzeyinde bir ağ gibi uzanır . Bu yapı özellikle kapillerlerden geçerken oluşan büyük basınca eritrositlerin dayanmasını sağlar. Yapılan çalışmalarda spektrin iskeletin zara ne şekilde tutunduğu radyoaktif işaretli spektrin kullanılarak araştırılmıştır. Buna göre; spektrinin zara bağlantısını geniş ölçüde bir hücre içi protein olan ankirin sağlar. Ankirin, (5-spektrin'e ve bir transmembran protein olan band 3' ün sitoplazmik kısmına aynı anda tutunur. Böylece spektrin zara tutunmuş olur. Spektrinin zara tutunmasında keza ikinci bir protein kümesi de yardım eder. Bu grupta, band 4.1 proteini spektrin ve aktin'e bağlanarak yine spektrinin zara tutunmasını sağlar. Bu grupta bazı durumlarda glikoforin (glycophorin)'in de yer aldığı gösterilmiştir.
 in de yer aldığı gösterilmiştir.")
22
Glikoforin (Glycophorin)
İnsan kırmızı kan hücre zarında bulunan ve bütün amino asit dizilişi tam olarak bilinen ilk protein Glikoforin'dir. Glikoforin, kütlesinin büyük kısmı zarın dış yüzeyinde bulunan, 131 amino asitten oluşmuş bir transmembran glikoproteindir.

24
Glikoforin (Glycophorin)
Glikoforin üç kısımdan meydana gelmiştir: 1. Amino uç kısım: Zarın dış yüzünde yer alan bu kısım 61 amino asitten oluşmuş bir zincir halindedir. Bu kısma 16 karbohidrat ünitesi bağlanmıştır. Karbohidratlardan 15 tanesi glikoforinin serin ve threonin amino asitlerine bağlanırken bir tanesi asparagin'e bağlanır. Bu oligosakkaritler genellikle negatif yüklü sialik asit grubu taşırlar. 61. pozisyondaki pozitif yüklü arginin, zarda yer alan, negatif yüklü bir fosfolipid baş grubuyla iyonik olarak etkileşir.

25
Glikoforin (Glycophorin)
2. Hidrofobik orta kısım: Glikoforini oluşturan 131 amino asitten, sıraları arasında yer alan 34 amino asitlik dizi proteinin zara tutunmasını sağlar. 66, 67, 70 ve 72. sıralarda yer alan yüklü amino asitler (histidin ve glutamik asit) hariç, diğerleri çoğunlukla hidrofobik amino asitlerden (fenilalanin, lösin, izolösin, valin, triptofan ve tirozin) meydana gelmiştir. Bu kısım bir α-sarmal halinde uzanarak zarı kateder.
 hariç, diğerleri çoğunlukla hidrofobik amino asitlerden (fenilalanin, lösin, izolösin, valin, triptofan ve tirozin) meydana gelmiştir. Bu kısım bir α-sarmal halinde uzanarak zarı kateder.")
26
Glikoforin (Glycophorin)
3. Karboksil uç kısım: Polar ve iyonize olmuş yan zincirlerce zengin olup sitoplazmada uzanır ve proteinin zara tutunmasına katkıda bulunur. Bu katkı, sitoplazmik yüzde yer alan fosfolipidlerin negatif yüklü baş kısımları ile glikoforinin 96, 97, 100 ve 101. sıralarında yer alan pozitif yüklü arginin ve lizin amino asitleri arasındaki iyonik ilişki sayesinde sağlanır. Glikoforin, sadece kırmızı kan hücre zarlarında bulunuyorsa da, onun yapısı lipid tabakayı bir α-sarmal halinde geçen transmembran proteinlerin yapısını açıklamada kullanılır. Bu nedenle bu tip proteinlere Tek geçişli zar proteinleri (single-pass transmembran protein) adı verilir.
 adı verilir.")
27
Band 3 Protein Glikoforin gibi Band 3 proteini de bir transmembran proteindir. Glikoforinden farklı olarak Band 3 proteini çok geçişli (multipass) bir zar proteinidir. Band 3 proteini lipid çift tabakayı 10 α-sarmal halinde kateden ve yaklaşık 900 amino asitten oluşmuş uzun bir proteindir. Bilindiği gibi eritrositlerin başlıca görevi akciğerlerden dokulara oksijen, dokulardan akciğere karbondioksit taşımaktır. Karbondioksitin oksijen ile değişimi bir anyon değiştirici transport proteini sayesinde olur. İşte eritrositlerdeki bu anyon transport proteinleri Band 3 proteinlerdir. Band 3 proteinleri dondurma-kırma elektron mikroskobi teknikleriyle incelendiğinde zar içi partiküller olarak görülür. Bu yolla incelendiğinde glikoforinin E, Band 3 proteinin ise P yüzde yer almaktadır
 bir zar proteinidir. Band 3 proteini lipid çift tabakayı 10 α-sarmal halinde kateden ve yaklaşık 900 amino asitten oluşmuş uzun bir proteindir. Bilindiği gibi eritrositlerin başlıca görevi akciğerlerden dokulara oksijen, dokulardan akciğere karbondioksit taşımaktır. Karbondioksitin oksijen ile değişimi bir anyon değiştirici transport proteini sayesinde olur. İşte eritrositlerdeki bu anyon transport proteinleri Band 3 proteinlerdir. Band 3 proteinleri dondurma-kırma elektron mikroskobi teknikleriyle incelendiğinde zar içi partiküller olarak görülür. Bu yolla incelendiğinde glikoforinin E, Band 3 proteinin ise P yüzde yer almaktadır.")
28
Bakteriorodopsin Yapısı çok iyi bilinen çok geçişli bir zar transport proteini de Bakteriorodopsin (Bacteriorhodopsin)'dir. Bu protein belirli bakterilerin zarlarında aktif proton (H+) pompası olarak görev yapar. Bir bakteri olan Halobacterium halobium'un zarı özel bir cins protein taşır. Bu protein Bakteriorodopsin (Bacteriorhodopsin)'dir. Bu protein, birbirine oldukça yakın olarak yer alan 7 α-sarmal halinde kıvrılmış olarak, lipid çift tabakada yer almaktadır. Her bir α -sarmal yaklaşık 25 amino asitten meydana gelir. Bu α -sarmal segmentler zarın her iki yanında hidrofilik segmentlerle birbirlerine bağlanırlar.
 dir. Bu protein belirli bakterilerin zarlarında aktif proton (H+) pompası olarak görev yapar. Bir bakteri olan Halobacterium halobium un zarı özel bir cins protein taşır. Bu protein Bakteriorodopsin (Bacteriorhodopsin) dir. Bu protein, birbirine oldukça yakın olarak yer alan 7 α-sarmal halinde kıvrılmış olarak, lipid çift tabakada yer almaktadır. Her bir α -sarmal yaklaşık 25 amino asitten meydana gelir. Bu α -sarmal segmentler zarın her iki yanında hidrofilik segmentlerle birbirlerine bağlanırlar.")
29
Membrane protein. Special proteins inserted in cellular membranes create pores that permit the passage of molecules across them. The bacterial protein shown here uses the energy from light (photons) to activate the pumping of protons across the plasma membrane. (Adapted from H. Luecke et al., Science 286:255–260, 1999.)
 to activate the pumping of protons across the plasma membrane. (Adapted from H. Luecke et al., Science 286:255–260, 1999.).")
30
Her bir bakteriorodopsin molekülü ayrıca retinal olarak adlandırılan, ışık absorbe edici bir kromofor (veya prostetik grup) ihtiva eder. Aynı yapıdaki bir kromofor omurgalıların retina çomak hücrelerinde de bulunur. Retinal, bakteriorodopsinin lizin yan zincirlerine kovalent olarak bağlanmıştır. Bir ışık fotonu tarafından uyarıldığında protein kompleksinde yapısal bir değişikliğe neden olarak bir veya iki protonun (H+) hücrenin dışına transferini gerçekleştirir. Bakteriyal fotosentetik reaksiyon merkezi bakterinin plazma zarında yer alır ve ışık enerjisini yakalayarak 1 nanosaniyeden daha kısa bir sürede zarı geçen yüksek enerjili elektronların oluşumunu sağlar. Bu elektronlarda daha sonra sitozolde ATP sentezlenmesi için elektron taşıma sistemi tarafından kullanılır.

31
Proteinler hücre zarlarının temel bileşenleri olup, hücrenin değişik zarlarının kütlesinin %25 - %75'ini oluşturur. Fosfolipidler zarların temel yapısal düzenini oluştururken, zar proteinleri hücrenin değişik zarlarının özgül işlevlerini yerine getirmektedir. Bu proteinler, zarla ilişkilerine göre iki temel sınıfta toplanırlar, Integral zar proteinleri lipid çift-tabakamn içine doğrudan gömülmüşlerdir. Periferal zar proteinleri lipid çift-tabakamn içine gömülmezler fakat zarla indirekt olarak, genelde integral zar proteinleri ile etkileşimler yoluyla, ilişkilidirler.

34
Lipid çift-tabakaları geçen tek diğer protein yapısı olarak, β-tabakaların fıçıya benzer bir yapı oluşturduktan β -fıçı yapısı bilinmektedir ki bu yapı, bakteri, kloroplastlar ve mitokondrilerin transmembran proteinlerinde görülmektedir.

35
Fosfolipidler gibi, transmembran proteinler, hidrofilik kısımları zarın her iki yüzeyindeki sulu ortamlar ile iç içe olan amfipatik moleküllerdir. Bazı transmembran proteinler zarı yalnızca bir kez geçmektedir; diğerlerinin, zan geçen çok sayıda bölgeleri vardır. Ökaryot plazma zarlarının transmembran proteinlerinin çoğu, karbohidratların bağlanmasıyla modifiye olmuşlardır; bu karbohidratlar, hücrenin dışı ile ilişkilidir ve hücre-hücre etkileşimlerine katılabilirler.

36
Zar Proteinlerinin Saf Olarak Elde Edilmesi
Genel olarak transmembran proteinler, lipid tabakayı tahrip ederek hidrofobik ilişkileri bozan çeşitli ajanlar tarafından çözündürülebilir. Bu ajanlar içinde en fazla kullanılan deterjanlardır. Bunlar suda misel şeklinde toplanma eğilimi gösteren amfipatik moleküllerdir. Zarlarla karıştırıldıklarında deterjanların hidrofobik uçları zar proteinlerinin hidrofobik bölgelerine bağlanır, böylece lipid moleküllerin yerini değiştirirler. Deterjan molekülünün diğer ucu polar olduğundan, protein molekülünün lipid tabakadan ayrılarak bir protein-deterjan kompleksi halinde solüsyon içerisine geçmesini sağlar. Bu komplekse nadiren çok kuvvetli bağlanmış lipid molekülleri de katılabilir. Deterjanlar dializ veya herhangi bir kromatografi tekniği kullanılarak uzaklaştırılmadan önce ortama fosfolipidler karıştırılır. Böylece sadece o proteini ihtiva eden özel bir fosfolipid çift tabaka elde edilir ve o proteinin istenen fonksiyonu bu ortamda araştırılır. Deterjanların polar uçları, ya sodyum dodesil sülfat (sodiurn dodecyl sulfate) (SDS) ta olduğu gibi elektiriksel olarak yüklü (iyonik) ya da triton'da olduğu gibi yüksüzdür (aniyonik).
 (SDS) ta olduğu gibi elektiriksel olarak yüklü (iyonik) ya da triton da olduğu gibi yüksüzdür (aniyonik).")
38
Zar Proteinlerinin Eritrositlerde İncelenmesi
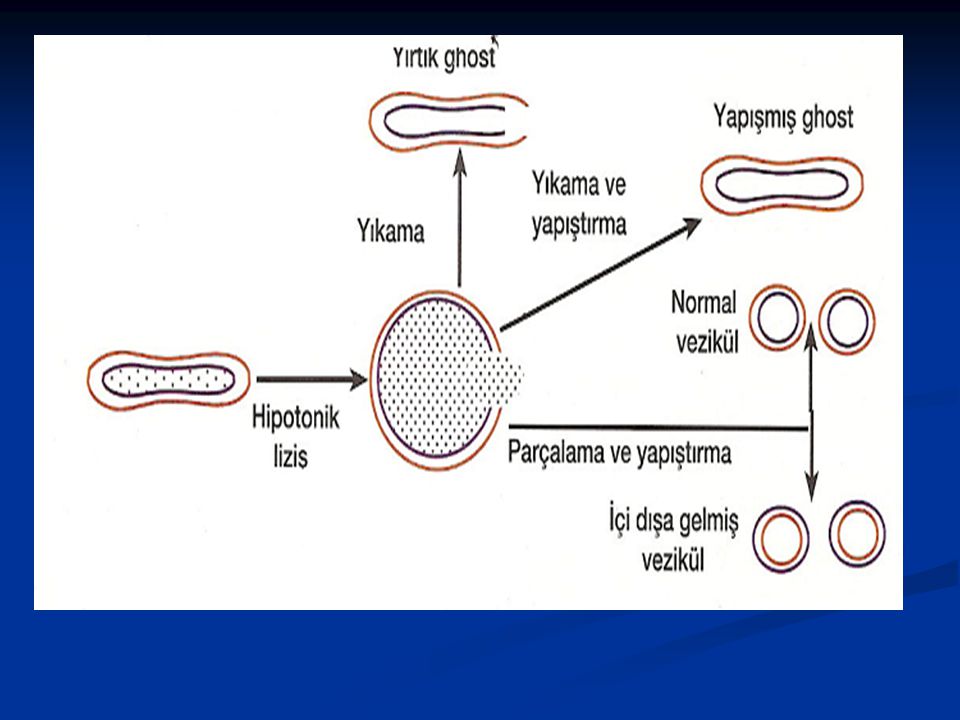
Ökaryotik hücrelerde hücre zarı ile yapılan çalışmalarda çoğunlukla kırmızı kan hücreleri kullanılır, çünkü: 1. Diğer hücrelerle karışma tehlikesi olmaksızın bol olarak elde edilebilir. 2. Çekirdek veya diğer organellere sahip olmadığından diğer iç zarlarla karışmadan, doğrudan saf olarak eritrosit zarı elde edilebilir. 3. Boş kırmızı kan hücre zarları veya kalıntıları (ghost) elde etmek çok kolaydır. Kan hücresinin sitoplazmasından daha düşük konsantrasyonlarda (hipotonik) solüsyonlar kullanıldığında, normal difüzyon kurallarına göre dış ortamdaki sıvı hücre içine girerek kırmızı kan hücrelerinin patlamasına (lysis) neden olur. 4. Zar kalıntıları ile fazla parçalanmadan, sadece tek noktadan yırtıkken çalışılabilir (böylece, çalışmada kullanılacak solüsyonlar isteğe göre zarın her iki yüzüyle temas edecektir), veya yırtığın kapanması sağlanır. Bu özellikten faydalanılıp, kullanılan değişik iyonik konsantrasyonlar sayesinde iç yüzü dışa gelmiş veziküller elde edilebilir.
 elde etmek çok kolaydır. Kan hücresinin sitoplazmasından daha düşük konsantrasyonlarda (hipotonik) solüsyonlar kullanıldığında, normal difüzyon kurallarına göre dış ortamdaki sıvı hücre içine girerek kırmızı kan hücrelerinin patlamasına (lysis) neden olur. 4. Zar kalıntıları ile fazla parçalanmadan, sadece tek noktadan yırtıkken çalışılabilir (böylece, çalışmada kullanılacak solüsyonlar isteğe göre zarın her iki yüzüyle temas edecektir), veya yırtığın kapanması sağlanır. Bu özellikten faydalanılıp, kullanılan değişik iyonik konsantrasyonlar sayesinde iç yüzü dışa gelmiş veziküller elde edilebilir.")
40
Bu tür çalışmalarla bazı zar proteinlerinin lipid tabakayı boydan boya geçtiği ve lipid çift tabakanın lipid kompozisyonunun birbirinden farklı olduğu gösterilebilmiştir. İnsan eritrosit zarlarında, SDS poliakrilamit jel elektroforezi ile yapılan çalışmalarla, molekül ağırlığı arasında değişen 15 büyük protein bandı elde edilmiştir Bu proteinlerden üçü, spektrin (spectrin), giikoforin (glycophorin) ve Band 3 toplam proteinlerin % 60 ı kadardır. Bu proteinlerin zarda diziliş şekilleri farklıdır bu yüzden bu 3 proteinin birleşme şekli proteinlerin zarla yapmış oldukları 3 farklı birleşme şeklinin örneği olarak açıklanır.
, giikoforin (glycophorin) ve Band 3 toplam proteinlerin % 60 ı kadardır. Bu proteinlerin zarda diziliş şekilleri farklıdır bu yüzden bu 3 proteinin birleşme şekli proteinlerin zarla yapmış oldukları 3 farklı birleşme şeklinin örneği olarak açıklanır..")
Benzer bir sunumlar












>")






 Nedir? Lipitlerdir.>")

