Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Amino asitlerin Karbon İskeletinin Katabolizması Prof Dr A Binnur Erbağcı Gaziantep Üniversitesi Tıbbi Biyokimya AD

2
Protein katabolizması C ve H N ve H CO2 ve H2O amonyak (toksik) endergonik üre (toksik değil)
 endergonik üre (toksik değil)")
3
Beslenme Sindirim Ince barsaktan emilim Yeni doku ve kan proteinlerinin sentezi amino asit havuzu Doku ve kan Proteinlerinin yıkımı esansiyel olmayan aa Diğer azotlu bileşikler Esansiyel olmayan aminoasitlerin biyosentezi Glukoneogenez ve lipogenez Enerji üretimi

4
Amino asitlerin karbon iskeletinin katabolizması
Sentez/ yıkım reaksiyonları farklı olabildiği için ayrı düzenlenebilirler. Bireyin fizyolojik durumu karbon iskeletinin akıbetini belirler Açlık: Karaciğer – C iskeletleri glukoz, keton cisimleri veya CO2 Tokluk: Glikojen ve TAG’e dönüşerek depo edilirler. Karaciğer tüm aa metabolizma enzimlerinin bulunduğu tek dokudur.

5
Amino Asitlerin Karbon İskeleti
Amino asitler deamine olarak a-keto asitlere dönüştüklerinde direkt olarak veya ilave reaksiyonlarla majör metabolik yollara girerler (ör., Krebs Döngüsü). Karbon iskeletlerinin dönüştükleri ürünlere göre 2 sınıfta incelenirler. Glukojenik ketojenik
. Karbon iskeletlerinin dönüştükleri ürünlere göre 2 sınıfta incelenirler. Glukojenik. ketojenik.")
6
Glukojenik aa’lerin karbon iskeletleri:
piruvat, veya 4-C veya 5-C Krebs Döngüsü metabolitlerine dönüşür, glukoneogenez öncülüdürler. 1-Glukoz düzeyleri düşük olduğunda glukojenik amino asitler glukoneogenez için majör karbon kaynağıdırlar. 2-Enerji sağlamak üzere katabolize olabilirler. 3-Enerji depolanması için glikojen veya yağ asitlerine dönüşebilirler.

7
Ketojenik aa’lerin karbon iskeletleri: asetil-CoA, veya
asetoasetata yıkılır. Asetil CoA ve öncülü, glukoza dönüşemez. neogenez öncülü olan oksaloasetata dönüşemez (Krebs döngüsüne giren her 2-C asetil kalıntısı CO2 olarak ayrılır). ketojenik amino asitlerin karbon iskeletleri Krebs Döngüsünde yıkılarak enerji sağlayabilir veya keton cisimlerine ve yağ asitlerine dönüşebilir.
. ketojenik amino asitlerin karbon iskeletleri Krebs Döngüsünde yıkılarak enerji sağlayabilir veya keton cisimlerine ve yağ asitlerine dönüşebilir.")
8
Karbon iskeletlerinin dönüştüğü ara ürünlere göre
Glukojenik aaler Piruvat Alfa-keto glutarat Süksinil KoA Fumarat Okzaloasetat Ketojenik aa’ler Asetoasetat Asetil KoA HMG KoA

9
C iskeletleri azot içermeyan karbohidrat veya lipidlere dönüşür
Böylece her aa’in karbon iskeletinin tümü veya bir bölümü Karbohidrata (13) Lipidlere ve keton cisimlerine (2) Her ikisine (6) dönüşebilir.
 Lipidlere ve keton cisimlerine (2) Her ikisine (6) dönüşebilir.")
10
Fazla olduklarında veya açlıkta bazı amino asitler tercihen glukoza bazıları ise yağ veya keton cisimlerine dönüşmektedir. citrate 20 Amino asit de su ve CO2’ye yıkılabilir malat isocitrat succinate succinat

11
Krebs döngüsü ara ürünlerine dönüşen aa’ler
Enerji sağlamak üzere krebs döngüsünde yıkılabilirler. Açlıkta glukoza dönüşerek kan glukoz düzeyini artırırlar Glukoz ihtiyacı yoksa glikojen şeklinde depo edilirler. Glikojen kapasitesi dolu ise lipitlere dönüşerek depolanabilirler (önemsiz miktarda).
.")
12
Krebs döngüsü ara ürünlerine dönüşen aa’ler
Enerji sağlamak üzere krebs döngüsünde yıkılabilirler. Açlıkta glukoza dönüşerek kan glukoz düzeyini artırırlar Glukoz ihtiyacı yoksa glikojen şeklinde depo edilirler. Glikojen kapasitesi dolu ise lipitlere dönüşerek depolanabilirler (önemsiz miktarda).
.")
13
Glikojen Kan İhtiyaç fazlası YA Açlık Glikojen
citrate Glikojen depolama kapasitesi aşıldığında ne olur? malate isocitrat succinat

14
Ketojenik aa’ler Açlıkta keton cisimlerine dönüşebilir ve keton cismi kullanan dokularda enerji sağlayabilirler. Enerji sağlamak üzere krebs döngüsünde yıkılabilirler. Enerji gereksinimi olmadığında lipitlere dönüşerek depolanabilirler. Piruvat dehidrogenaz reaksiyonu geri dönüşümsüz olduğu için karbohidratlara dönüşemezler.

15
YA Fazla protein citrate Açlıkta malate isocitrate succinate

16
Hem ketojenik hem de glukojenik olan aa’ler
Her iki grup bileşiğe de dönüşebilirler Enerji sağlamak üzere krebs döngüsünde yıkılabilirler.

17
YA FA citrate malate isocitrate succinate

18
Piruvata dönüşen aa’ler
Piruvat asetil KoA öncülü olduğu için piruvata dönüşen aa’ler glukojenik grubunda olmalarına rağmen lipidlere de dönüşebilir. Açlıkta glukoza İhtiyaç fazlası olduğunda lipidlere dönüşürler. Enerji sağlamak üzere krebs döngüsünde yıkılabilirler.

19
Karaciğerde Açlık citrate malate isocitrate succinate

20
İhtiyaç fazlası YA citrate malate isocitrate succinate

21
Glukojenik ve ketojenik
Alanin Arginin Aspartat Asparagin Sistein Glutamat Glisin Histidin Prolin, OH-prolin Metionin Serin Valin Glukojenik ve ketojenik İzolösin Fenilalanin Triptofan Tirozin Treonin Ketojenik Lösin Lizin

22
Oksaloasetat ailesi Aspartat ve asparagin oksaloasetata dönüşürler
Enzimler: Asparaginaz ve aspartat aminotransferaz (AST)
")
23
Asparagin amido grubunu Asparaginaz reaksiyonunda kaybederek aspartata dönüşür.

24
Aspartat da oksaloasetata, dönüşebilir ör., transaminasyon ile

25
-keto glutarat ailesi
Glutamin, Glutamat Prolin Arginin Histidin -keto glutarata dönüşürler.

26
Glutamat ve glutamin a-ketoglutarata dönüşür
Glutaminaz reaksiyonu asparaginaza benzer +3HN-CH-COO- CONH2 CH2 COO- NH4+ Glutamin Glutamat Glutaminaz H2O Asparagine Aspartate Asparaginase Glutamat glutamat dehidrogenazla a-ketoglutarat ve amonyağa dönüşür O=C-COO- COO- CH2 +3HN-CH-COO- NH4+ NAD+ NADH

27
Prolin yıkımı sentezinin tersine benzer reaksiyonları ile gerçekleşir
Prolin, sentezinin tersine benzer yollarla glutamat ve -ketoglutarata dönüşür. Arginin ve ornitin glutamata dönüşür.

28
Histidin Histidin katabolizması ile glutamat ve -ketoglutarata dönüşür. Histidin katabolizmasında önemli bir basamak, FİGLU oluşumudur.

29
N-formiminoglutamat (FİGLU), formimino grubunu tetrahidrofolata (THF), aktarır, kendisi glutamata dönüşürken N5-formimino-THF oluşturur.
, formimino grubunu tetrahidrofolata (THF), aktarır, kendisi glutamata dönüşürken N5-formimino-THF oluşturur.")
30
Histidinin glutamata yıkılışı yolunda son reaksiyon kofaktör olarak tetrahidrofolat gerektirir.
Tetrahidrofolat (THF), yapısında bir pteridin halkası tasıyan folat’ın (B grubu vitamin) indirgenmiş formudur. Yapısında birden fazla olabilen glutamat kalıntısı taşır.
, yapısında bir pteridin halkası tasıyan folat’ın (B grubu vitamin) indirgenmiş formudur. Yapısında birden fazla olabilen glutamat kalıntısı taşır.")
31
THF N5 veya N10 ‘a bağlı veya ikisi arasında köprü oluşturacak şekilde bulunan tek-C ünitesi taşıyıcısıdır. Bu C üniteleri farklı oksidasyon durumlarında olabilir. Şekilde görülen N10 r-aminobenzoik asite aittir. R glutamat kalıntılarıdır. Hücresel THF havuzu farklı formlarda THF içerir ve bunlar farklı reaksiyonlarda kullanılır.

32
Histidin bu reaksiyonda tek karbonlu bir grubunu (formimino) diğer biosentez tepkimelerinde kullanılmak üzere THF’a aktarmıştır. Bu nedenle histidin tek karbon kaynağı olan bir amino asittir.

33
Folat Eksikliğinde idrarla Figlu atılır
Folat eksikliğinde bu reaksiyon kısmen veya tümü ile kesintiye uğrar ve idrarla FİGLU atılır.

34
a-ketoglutarat ailesi
arginin glutamin glutamat histidin TH folat & prolin.

35
Piruvat Ailesi Alanin, Serin, Glisin, sistein, treonin (2C atomu)
Triptofan (+Asetoasetil CoA) ve OH-prolin 3C’lu keto asit olan piruvat oluşturur.
 ve OH-prolin 3C’lu keto asit olan piruvat oluşturur.")
36
Alanin transaminasyonla piruvata dönüşür.

37
Glisin ve Serin birbirine dönüşebilir
Bu nedenle katabolizmaları birbirine dönüşerek olabilir. Serin Hidroksimetil Transferaz Serin Glisin THF N5N1OCH2THF

38
SERİN Serin için başlıca yıkım yolu glisine dönüşüm ile olur.
Bu dönüşüm serinin metilen grubunun THF’a aktarılmasını içerir. THF de bu bir karbonlu grubu diğer biyosentez reaksiyonlarında başka alıcılara aktarır. Bu nedenle serin bir karbon metabolizmasının önemli kaynaklarından birini oluşturur. Serin en önemli ‘bir karbon’ kaynağıdır.

39
Alternatif yol piruvata dönüşümdür.
Glisin de serine dönüşerek piruvata yıkılabilir. Not: bu reaksiyon serin, glisin ve treonin’in glukojenik olma nedeni

40
Glisin yıkım ürünleri Glisin çeşitli öncül bileşiklerden sentezlenebildiği gibi, katabolizması ile de farklı ürünler oluşabilir: Serin CO2, NH4, metilen (-CH2) Glioksalat
 Glioksalat.")
41
Glisin sentaz kompleksi
Glisin ve serin için majör yıkım yolu geri dönüşümlü bir tepkime olan glisin sentaz kompleksi ile olur (Karaciğer mitokondria). Ürün CO2 ve amonyaktır Glisin de “bir karbon” kaynağıdır. H4 Folat N5N1OCH2THF Glisin+NAD CO2+NH4+NADH+H Glisin sentaz (PLP)
. Ürün CO2 ve amonyaktır. Glisin de bir karbon kaynağıdır. H4 Folat. N5N1OCH2THF. Glisin+NAD. CO2+NH4+NADH+H. Glisin sentaz (PLP)")
42
Glisin glioksalata yıkılabilir
Glioksalat idrarla atılan oksalata oksitlenebilir. Oksalat üriner sistem taşları patogenezinde rol oynar. Glioksalat Glisin Amino asit -ketoasit

43
Sistin ve Sistein Sistin, sistin redüktaz ile sisteine dönüşür. Bu reaksiyondan sonra sistin ve sistein katabolizması kesişmektedir. Sistin redüktaz 2 Sistein Sistin NADH+H NAD Sistein iki farklı yolla piruvata dönüşebilir

44
Aldolaz treonin dehidrogenaz SİSTEİN GLİSİN ALANİN SERİN TREONİN
Piruvata dönüşümleri a-Amino-b- ketobutirat Asetaldehit Treonin Aldolaz Asetil-KoA Glisin N5,N10-Methylene-THF Süksinil KoA serin hidroksi metil- transferaz THF glisin sentaz Glisin Sistein Alanin Serin ALT serin dehidrataz ÇEŞİTLİ YOLLAR H2S SO3-2 SCN- Piruvat Willmore 2002

45
Piruvat Ailesi Alanin Triptofan serin glisin treonin Tetrahidrofolat
Sistin ve sistein OH prolin Triptofan serin glisin treonin

46
Fumarat Ailesi Fenil alanin ve tirozin katabolizması ile fumarat oluşur (Ürün olarak asetoasetat da oluştuğu için farklı başlıkta incelenecek)
")
47
Süksinil Koenzim A Ailesi
Methionin Treonin (piruvat+ asetil KoA) Valin İzolösin (+asetil CoA) Propionil-CoA, bu metabolik yollarda oluşan bir ara üründür, tek sayılı C içeren yağ asitlerinin b-oksidasyonu ile de oluşur.
 Valin. İzolösin (+asetil CoA) Propionil-CoA, bu metabolik yollarda oluşan bir ara üründür, tek sayılı C içeren yağ asitlerinin b-oksidasyonu ile de oluşur.")
48
SAM aktif metionindir Metionin süksinil CoA ailesinin en önemli üyesidir. Katabolizması serinden sistein sentez yollarını da içerir. İlk basamakta ATP ve metyonin kondansasyonu ile S-adenozil metyonin (SAM) yani aktif metyonin oluşur.
 yani aktif metyonin oluşur.")
49
.

50
Homosistein + Serin Sistatyonin sentaz Sistatyonin Sistein NH4 -Keto bütirat Propionil CoA

51
Propionil-CoA biotin ve B12 vitaminlerini gerektiren reaksiyonlarla süksinil-CoA’ya dönüşür.

52
Metilmalonil-CoA Mutaz’ın prostetik grubu koenzim B12‘dir (vitamin B12, kobalamin) .
 .")
53
Dallı zincirli aa katabolizması
Valin Lösin İzolösin Başlangıç reaksiyonlar her 3 aa için aynı olmasına karşın farklı son ürünler oluşur. Yıkımları yağ asitlerinin katabolizmasına benzerlik gösterir.

54
Oksidatif Dekarboksilasyon
Lösin Valin İzolösin Transaminasyon Oksidatif Dekarboksilasyon Dehidrojenasyon Asetil CoA Asetoasetat Propionil CoA Metil malonil CoA B12 koenzim Süksinil CoA

55
12 Amino asit Asetil CoA Oluşturur
Piruvat oluşturan aa’ler (7 aa): Alanin, serin, glisin, OH-prolin, sistein, treonin, triptofan piruvat dehidrojenaz ile asetil CoA’ya dönüşebilirler. (non-ketojenik aa’lerdir.) Diğer (7 aa): Fenilalanin, Tirozin, Triptofan, Treonin, Lizin, İzolösin ve Lösin ise önce piruvat oluşturmadan Asetil CoA’ya dönüşürler, yani ketojenik özellik gösterirler. treonin, triptofan Triptofan Treonin
: Alanin, serin, glisin, OH-prolin, sistein, treonin, triptofan piruvat dehidrojenaz ile asetil CoA’ya dönüşebilirler. (non-ketojenik aa’lerdir.) Diğer (7 aa): Fenilalanin, Tirozin, Triptofan, Treonin, Lizin, İzolösin ve Lösin ise önce piruvat oluşturmadan Asetil CoA’ya dönüşürler, yani ketojenik özellik gösterirler. treonin, triptofan. Triptofan. Treonin.")
56
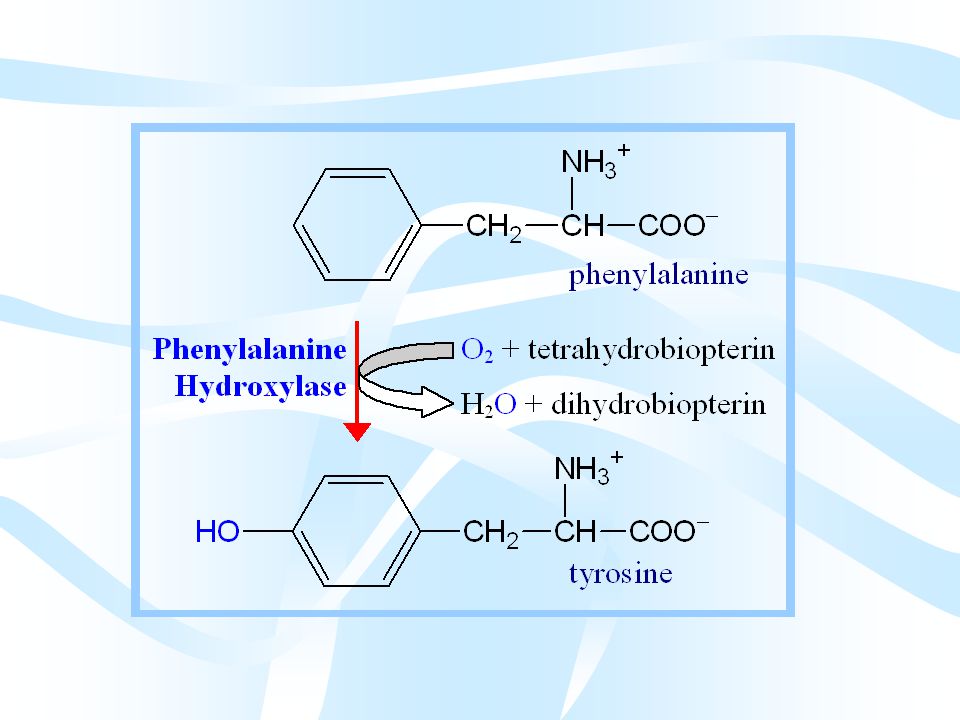
Fenilalanin Tirozine Dönüşür
Fenilalanin’in tirozine dönüşümü tirozin sentezinde gördüğümüz fenil alanin hidroksilaz reaksiyonudur. Fenilalanin’in tirozine dönüşümünden sonra katabolizmaları birlikte seyreder. Bu katabolik yolla ilgili sık görülen kalıtsal metabolizma bozuklukları vardır.

58
Tirozin ve fenil alanin katabolizmasının son ürünleri fumarat ve asetoasetattır.
Bu nedenle bu iki aa hem ketojenik hem glukojenik özelliktedirler.

59
p-hidroksifenilpiruvat
Tirozin p-hidroksifenilpiruvat Homogentisat Maleilasetoasetat Fumarilasetoasetat Fumarat + Asetoasetat Tirozin Aminotransferase Hidroksifenil piruvat oksidaz Homogentisat Fumarilasetoasetat hidrolaz

60
Triptofan Katabolizması sırasında Alanin açığa çıktığı, son ürün asetoasetat olduğu için Triptofan ketojenik ve glukojenik bir aa’tir. İlk basamağı triptofan oksijenaz (triptofan pirolaz) kataliz eder.
 kataliz eder.")
61
Triptofan İlk basamağı triptofan oksijenaz (triptofan pirolaz) kataliz eder. Bu enzim adrenokortikal steroidler ile indüklenebilir. O2 N-formil kinürenin Triptofan triptofan oksijenaz

62
Triptofan N formil kinürenin ‘Bir karbon’ ünitesi ayrılır Kinürenin Ksantürenat OH-kinürenin PLP, Kinüreninaz Alanin OH-antranilat Piruvat -keto adipat Asetoasetil CoA

63
Nikotinat mono nükleotid (NMN)
")
64
PLP Eksikliğinde idrarla ksantürenat atılımı artar.
Piridoksal Fosfat (PLP, B6) eksikliğinde Kinüreninaz enzimini aktivitesi engellenir. Kinürenin bileşiklerinin metabolizmasının engellenmesi sonucu Ksantürenat fazla miktarda oluşur ve idrarla atılımı artar.
 eksikliğinde Kinüreninaz enzimini aktivitesi engellenir. Kinürenin bileşiklerinin metabolizmasının engellenmesi sonucu Ksantürenat fazla miktarda oluşur ve idrarla atılımı artar.")
65
citrate malate isocitrate succinate succinate

66
Bir Karbon metabolizması
Bir Karbon metabolizması organik moleküllerin metabolizması ile açığa çıkan “bir Karbonlu” ünitelerin aracı moleküller ile biyosentez reaksiyonlarında kullanılmasıdır. Bu aracı moleküllere bir karbon taşıyıcıları diyoruz; THF, SAM ve B12 vitamini (kobalamin)’dir. (Tek karbonlu bir ünite olan CO2 tek karbon havuzu üyesi olarak kabul edilmez)
’dir. (Tek karbonlu bir ünite olan CO2 tek karbon havuzu üyesi olarak kabul edilmez)")
67
THF: farklı üniteler taşır
THF’ın en önemli bir karbon kaynakları seringlisin dönüşümü glisin katabolizması histidin katabolizması ve triptofan katabolizmasıdır THF bir karbonlu üniteleri nükleotid sentezi, homosisteinin remetilasyonu gibi reaksiyonlara aktarır.

68
SAM en önemli metil grubu vericisidir
SAM metioninden geri dönüşümsüz bir tepkime ile sentezlenir. N5-metil-tetrahidrofolat (N5-metil-THF)’tan 1000 kat daha güçlü bir metil grubu vericisidir. SAM metil grubunu fosfolipid sentezi, nükleotid sentezi, aa türevlerinin sentezi (kreatin, poliamin, aktif aminler...), DNA metilasyonu, post translasyonel protein metilasyonu reaksiyonlarına aktarır.
’tan 1000 kat daha güçlü bir metil grubu vericisidir. SAM metil grubunu fosfolipid sentezi, nükleotid sentezi, aa türevlerinin sentezi (kreatin, poliamin, aktif aminler...), DNA metilasyonu, post translasyonel protein metilasyonu reaksiyonlarına aktarır.")
69
Vitamin B12 Bu dersimizde vit B12’nin kofaktör olarak rol aldığı iki önemli reaksiyonu gördük. 1- metil malonil KoA’nın süksinil KoA’ya dönüşümü; Tek sayılı C ünitesi taşıyan yağ asitlerinin ve süksinil KoA’ya dönüşen aa’lerin glukoneogeneze girişi için önemli bir reaksiyondur. B12 eksikliğinde metilmalonik asidüri meydana gelir.

70
Vitamin B12 (devam) 2- Metionin’in homosisteinden rejenerasyonu: B12 türevi olan metil kobalamin ve N5-metil-tetrahidrofolat (N5-metil-THF) metioninin homosisteinden yeniden sentezinde gereklidirler. Bu reaksiyon bloke olduğunda idrarla homosistin atılımı meydana gelir: homosistinüri bu reaksiyon aynı zamanda metil-THF’ın THF’a dönüşümü için önemli. B12 vitamini eksikliğinde metil-THF’ın THF’a dönüşemez ve THF nükleik asit sentezine giremez. Buna folat tuzağı denilir.
 metioninin homosisteinden yeniden sentezinde gereklidirler. Bu reaksiyon bloke olduğunda idrarla homosistin atılımı meydana gelir: homosistinüri. bu reaksiyon aynı zamanda metil-THF’ın THF’a dönüşümü için önemli. B12 vitamini eksikliğinde metil-THF’ın THF’a dönüşemez ve THF nükleik asit sentezine giremez. Buna folat tuzağı denilir.")
71
Homosistein Metabolizması
R PPi+Pi S-Adenosyl Methionine (SAM) R-CH3 ATP 2 1 DMG S-Adenosyl Homocysteine Diet Methionine 5 Betaine H2O 4 3 THF B12 Homocysteine B6 Ado 8 6 N5-CH3-THF Cystathionine 9 7 B6 N5,510-CH2-THF Cysteine
 R-CH3. ATP DMG. S-Adenosyl Homocysteine. Diet. Methionine. 5. Betaine. H2O THF. B12. Homocysteine. B6. Ado N5-CH3-THF. Cystathionine B6. N5,510-CH2-THF. Cysteine.")
72
İdrarla neler atılıyordu?
Eksik vit ilgili Aa Atılan THF PLP B12 Histidin Figlu Triptofan Kinüreninat Metionin Homosistin Metionin Metilmalonik asit Valin izolösin

74
Orotik asit aşağıdaki metabolik yolların hangisinde oluşur?
A) Pürin bazlarının sentezi B) Pirimidin bazlarının sentezi C) Sfingolipidlerin sentezi D) Fosfogliseridlerin sentezi E) Üre sentezi
 Pürin bazlarının sentezi. B) Pirimidin bazlarının sentezi. C) Sfingolipidlerin sentezi. D) Fosfogliseridlerin sentezi. E) Üre sentezi.")
75
32. Bir hastanın idrarında metilmalonik asit düzeylerinin artması aşağıdaki B vitaminlerinden hangisinin eksikliğini gösterir? A) Vitamin B6 B) Vitamin B12 C) Riboflavin D) Folik asit E) Biotin
 Vitamin B6 B) Vitamin B12 C) Riboflavin D) Folik asit E) Biotin.")
76
Aşağıdakilerden hangisi üre sentezinin başlaması için mutlaka gereklidir?
A) N-asetil glutamat B) Asetoasetat C) Okzaloasetat D) a keto glutarat E) Piruvat
 N-asetil glutamat. B) Asetoasetat. C) Okzaloasetat. D) a keto glutarat. E) Piruvat.")
Benzer bir sunumlar