Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
MUTASYON ve MUTAJENLER
Doç. Dr. Öztürk ÖZDEMİR Aralık 2004

2
Mutasyon: Genomik yapıda (DNA ya da RNA) meydana gelen değişikliklerin tümüne denir
 meydana gelen değişikliklerin tümüne denir")
3
MUTASYON 1-Meydana geliş biçimi, tipi ve etiyolojisinde yatan etkenler 2-Meydana geldiği doku – hücre tipi 3-Meydana geldiği hücre – doku evresi 4- Meydana geldiği DNA dizileri – gen bölgesi 5- Büyüklüğü 6- Fenotipte rol alıp alamayacağı açısından değerlendirilmelidir.

4
MUTASYONLAR I- Fenotipteki etkilerine göre II- Büyüklüklerine göre
III- Meydana geliş biçimlerine göre IV- Genom-gen üzerindeki etkilerine göre ilk aşamada gruplandırılır.

5
I-Fenotipik etkilerine göre
Patojenik- patolojik mutasyonlar Non-patojenik- patolojik olmayan mutasyonlar - silent mutasyonlar - resessif mutasyonlar -non-genomik bölge mutasyonları

6
II-Büyüklüklerine göre mutasyonlar
Mikroskopik (makro mutasyonlar) -2000 kb ve daha büyük Submikroskopik (mikro mutasyonlar) - Bir tek baz yada - mikroskop düzeyinde değerlendirilmeyecek kadar küçük bant ya da subbant düzeyinde olamayan mutasyonlar
 kb ve daha büyük. Submikroskopik (mikro mutasyonlar) - Bir tek baz yada. - mikroskop düzeyinde değerlendirilmeyecek kadar küçük bant ya da subbant düzeyinde olamayan mutasyonlar.")
7
A-Mikroskopik mutasyonlar
Işık ya da elektron mikroskoplarla kolaylıkla ve kromozom düzeyinde saptanabilen: total kromozom, kromozom kolu, bant, subbant düzeyinde DNA bölgesini ilgilendiren 2000 kb büyüklüğündeki mutasyonlara denir (Makromutasyonlar).
.")
8
A- Mikroskopik mutasyonlar
Kromozomal Mutasyonlar, iki temel grupta toplanır; 1. Sayısal Kromozom Anomalileri: 1.a, Euploidi : Haploid yapının katları şeklindeki kromozom artışları - Monoploidi : n sayıda kromozom, insanda bu değer 23 tür - Diploidi : 2n - Triploidi : 3n - Tetraploidi : 4n - Pentaploidi : 5n ( insanda 115 kromozom durumu) 1.b, Aneuploidi : Haploid yapının katları şeklinde olmayan kromozom artışları (Hiperaneuploidi, n yada 2n+1) yada azalışları (hipoaneuploidi, n yada 2n-1) Örneğin: Down sendromu ,XX+21 Klinefelter sendromu 47, XXY Trizomi 13 sendromu 47, XX+13 Turner sendromu , X
 1.b, Aneuploidi : Haploid yapının katları şeklinde olmayan kromozom artışları (Hiperaneuploidi, n yada 2n+1) yada azalışları (hipoaneuploidi, n yada 2n-1) Örneğin: Down sendromu 47,XX+21. Klinefelter sendromu 47, XXY. Trizomi 13 sendromu 47, XX+13. Turner sendromu 45, X.")
9
2- Yapısal Kromozom Anomalileri 2. 1 Translokasyon;
2- Yapısal Kromozom Anomalileri 2.1 Translokasyon; -Birden fazla kromozomlararası parça alışverişi -Kromozomlar homolog ya da non-homolog olabilir -Transloke olunan parça kromozomun kısa ya da uzun kolu düzeyinde olabileceği gibi bant, subbant düzeyinde de olabilir Resiprokal translokasyon (Robertsonian) Non-resiprokal translokasyon Sentrik füzyon translokasyon 2.2 Delesyon; - Birden fazla kromozomda meydana gelebilir - Kromozomdan bir ya da birden fazla gen bölgesinin kaybına denir - Kromozomda en az iki kırılma bölgesinin olması gerekir 2.3. İnsersiyon; Herhangi bir kromozomun farklı bölgelerine başka bir kromozoma ait bant ya da subbant düzeyinde bir kısmının katılmasıdır.
 Non-resiprokal translokasyon Sentrik füzyon translokasyon 2.2 Delesyon; - Birden fazla kromozomda meydana gelebilir - Kromozomdan bir ya da birden fazla gen bölgesinin kaybına denir - Kromozomda en az iki kırılma bölgesinin olması gerekir 2.3. İnsersiyon; Herhangi bir kromozomun farklı bölgelerine başka bir kromozoma ait bant ya da subbant düzeyinde bir kısmının katılmasıdır.")
10
2. 4. İnversiyon;. - Perisentrik :sentromeri içeriyorsa
2.4. İnversiyon; - Perisentrik :sentromeri içeriyorsa - Parasentrik: sentromeri içermiyorsa 2.5. Dublikasyon; İzokromozom;Tam metasentrik yapıda kromozom Ring kromozom Gap (aralık) Frajil bölge yapısı ; FraXq – Martin Bell sendromu. İnsanda en yaygın frajil bölge mutasyonu gösteren kromozomlar ; 2q13,6p23,9p23, 12q13 ve 20p11 dir.
 2.9. Frajil bölge yapısı ; FraXq – Martin Bell sendromu. İnsanda en yaygın frajil bölge mutasyonu gösteren kromozomlar ; 2q13,6p23,9p23, 12q13 ve 20p11 dir.")
11
B- Submikroskopik mutasyonlar
Mikroskopla varlığı saptanamayan, bir tek baz (A;G;C ya da T olabilir) ya da 200 kb büyüklüğünde olan mutasyonlara denir (Mikromutasyonlar ya da nokta mutasyonlar).
 ya da 200 kb büyüklüğünde olan mutasyonlara denir (Mikromutasyonlar ya da nokta mutasyonlar).")
12
I- Baz düzeyinde Meydana Gelen Mutasyonlar 1
I- Baz düzeyinde Meydana Gelen Mutasyonlar 1.Transisyon ; Pürin – Pürin ya da Primidin –Primidin 2.Transversiyon ; Pürin – Primidin ya da Primidin –Pürin 3. İnsersiyon 4. Delesyon 5. Frame –Shift (çerçeve kayması) 6. Silen (Sessiz) mutasyon: DNA üçlü (triplet) kodonunda bir nokta mutasyona rağmen kodondan sentezlenen aminoasitde değişikliğe neden olmuyorsa denir. Ör: TTA TTG (Transisyonel silent mutasyon) lösin lösin
 6. Silen (Sessiz) mutasyon: DNA üçlü (triplet) kodonunda bir nokta mutasyona rağmen kodondan sentezlenen aminoasitde değişikliğe neden olmuyorsa denir. Ör: TTA TTG (Transisyonel silent mutasyon) lösin lösin.")
13
7. Missens (Kayıp) Mutasyon: DNA üçlü (triplet) kodonunda bir nokta mutasyon sonrasında kodondan sentezlenen aminoasitde değişikliğe neden oluyorsa denir. Ör: GCA GAA (Transversiyonel missens mutasyon) alanin glutamik asit 8. Nonsens Mutasyon: Üçlü (triplet) kodonda meydana gelen bir nokta mutasyon sonrasında kodon STOP kodona neden oluyorsa denir. Ör: TTA TGA (Transversiyonel missens mutasyon) alanin STOP
 Mutasyon: DNA üçlü (triplet) kodonunda bir nokta mutasyon sonrasında kodondan sentezlenen aminoasitde değişikliğe neden oluyorsa denir. Ör: GCA GAA (Transversiyonel missens mutasyon) alanin glutamik asit 8. Nonsens Mutasyon: Üçlü (triplet) kodonda meydana gelen bir nokta mutasyon sonrasında kodon STOP kodona neden oluyorsa denir. Ör: TTA TGA (Transversiyonel missens mutasyon) alanin STOP.")
14
II-Gen Düzeyinde Meydana Gelen Mutasyonlar
II-Gen Düzeyinde Meydana Gelen Mutasyonlar 1-Regülatör (düzenleyici) gen alt birimi mutasyonları 2- Promotor (idame ettirici) gen alt birimi mutasyonları 3- Yapısal gen alt birimi mutasyonları 4- Operatör gel alt birimi mutasyonları 5- Toplam gen delesyonları 6- Enhancer bölge mutasyonları 7- Silencer bölge mutasyonları
 gen alt birimi mutasyonları 2- Promotor (idame ettirici) gen alt birimi mutasyonları 3- Yapısal gen alt birimi mutasyonları 4- Operatör gel alt birimi mutasyonları 5- Toplam gen delesyonları 6- Enhancer bölge mutasyonları 7- Silencer bölge mutasyonları.")
15
Frame-shift Mutasyon 1 4 7 10 13 16 19 ATG GGA GCT CTA TTA ACC TAA
ATG GGA GCT CTA TTA ACC TAA met gly ala leu leu thr stop ATG GGG AGC TCT ATT AAC CTA ATT TGA met gly ser ser ile asn leu ile stop

16
Mis-sens Mutasyon (Transversiyonel-mis-sens nokta mutasyon)
ATG GGA GCT CTA TTA ACC TAA met gly ala leu leu thr stop ATG GGA GCT CTA TTT ACC TAA met gly ala leu phe thr stop

17
Non-sens / tRNA supressör Mutasyon (Transversiyonel-non-sens nokta mutasyon)
ATG GGA GCT CTA TTA ACC TAA met gly ala leu leu thr stop Non-sens ATG GGA GCT CTA TGA ACC TAA met gly ala leu Stop tRNA supressör met gly ala leu trp thr stop

18
Revers Mutasyon (Ters- birinci revers - mutasyon)
ATG GGA GCT CTA TTT ACC TAA met gly ala leu phe thr stop 2.mutasyon ATG GGA GCT CTA TTA ACC TAA met gly ala leu leu thr stop

19
Revers Mutasyon (İkinci bölge - second side - revers mutasyon)
ATG GGA GCT CTA TTT ACC TAA met gly ala leu phe thr stop 2.mutasyon ATG GGA GCT CTA CTT ACC TAA met gly ala leu leu thr stop

20
GEN DÜZEYİNDE GÖRÜLEN MUTASYONLARIN PATOJENİTE DÜZEYLERİ
Mutasyonun Gen fonksiyonu gendeki yerleşimi üzerindeki etkisi Yorum Multigen delesyonu GEOK Bileşik gen sendromları Toplam gen delesyonu GEOK Ağır klinik tablo Toplam ekson kaybı GEOK Unstabil protein sentezi Ekson içi delesyonu GEOK Prematür stop kodon oluşumu Toplam intron kaybı yok __ Splice bölge mutasyonu GEOK yada ekspresyonda azalma Modifiye protein sentezi Promotor bölge mutasyonu “ Promotor bölgenin toplam mutasyonu gen fonksiyonunu tamamen ortadan kaldırır Stop kodon mutasyonu Modifikasyon Modifiye protein sentezi PoliA sinyal bölge mutasyonu GEOK toplam delesyon non-fonksiyonel gene neden olur -kısmi delesyon yada insersiyon protein ekspresyonunu değiştirir

21
NOKTA MUTSYONU ORANLARI
Mutasyon Tipi Sayı % Büyük delesyonlar Kompleks rearrengements Büyük İnsersiyonlar ve dublikasyonlar Rpeat extepentions Küçük insersiyon ve delesyonlar Küçük insersiyonlar Küçük delesyonlar

22
NOKTA MUTSYONU ORANLARI
Mutasyon Tipi Sayı % Regülatör bölge/tek baz Splisiyonel/tek baz Nonsens Missens Silent nadir Frame-Shift nadir

23
III- MEYDANA GELİŞ MEKANİZMALARINA GÖRE MUTASYONLAR
1- Spontan – sporadik 2- Kromozomlararası yeni düzenlemelerle meydana (rearrangement) gelen de novo 3- Kalıtsal (ailesel) Mutasyonlar 4- İndüklenmiş Mutasyonlar (Akciğer kanserleri + polisiklikhidrokarbonlar)
 gelen de novo. 3- Kalıtsal (ailesel) Mutasyonlar. 4- İndüklenmiş Mutasyonlar (Akciğer kanserleri + polisiklikhidrokarbonlar)")
24
ORGANİZMA DÜZEYİNDE ETKİLİ MUTASYONLAR
Organizmada meydana gelebilecek herhangi bir mutasyon, fonksiyonel bir proteinin sentezini ilgilendiriyor ve organizma söz konusu mutant proteinin farklı sentezlenmesi yada sentezlenmemesini tolere edemiyor ve bu durum organizmanın ölümü ile sonuçlanıyorsa bu mutasyonlara “LETAL” mutasyonlar adı verilir.

25
Letal Mutasyonlar; 1- Oksotrofik (Auxotrophic) Mutasyonlar: Mutasyon temel bir aa gibi esensiyel bir metabolitin biyosentezini ilgilendiriyor ve bu metabolitin yokluğunda hücre yaşamını sürdüremiyorsa denir. Bu mutasyonlarda, ilgili metabolit dışarıdan verilirse mutasyon etkisi ortadan kaldırılır ve organizma yaşamına devam eder. 2- Protrofik Mutasyonlar: Dışarıdan ilgili metabolit temin edilse dahi mutajenik etkisi ortadan kaldırılamayan letal mutasyonlara denir. 3- Regülatör (düzenleyici) Mutasyonlar 4- Revers (ters) Mutasyonlar Geri İkinci revers Amber (Supressör tRNA)
 Mutasyonlar 4- Revers (ters) Mutasyonlar 4.1- Geri 4.2- İkinci revers 4.3- Amber (Supressör tRNA).")
26
MUTAJENLER I- Fiziksel Mutajenler a- Isı b-pH c- Işınlar
1-İyonize ışınlar (X ve gamma) 2-Non-iyonize (UV, 260 nm dalga boyu ışınlar) 3-Mor ötesi ışınlar II-Kimyasal Mutajenler a- Baz Analogları (5-Bromodeoksiuridin-BrdU, 6-thioguanin, 2-aminopürinler en yaygınları) b- Deaminasyon yapan ajanlar: DNA yapısında amino gruplarının kaybına neden olan ajanlar (Nitröz asidi, hidroksil aminler) c- Alkilleyici ajanlar: DNA yapısına alkil grubu takan ajanlar ( Kükürt, Nitrojen mustard, Etilenoksitler)
 2-Non-iyonize (UV, 260 nm dalga boyu ışınlar) 3-Mor ötesi ışınlar. II-Kimyasal Mutajenler. a- Baz Analogları (5-Bromodeoksiuridin-BrdU, 6-thioguanin, 2-aminopürinler en yaygınları) b- Deaminasyon yapan ajanlar: DNA yapısında amino gruplarının kaybına neden olan ajanlar (Nitröz asidi, hidroksil aminler) c- Alkilleyici ajanlar: DNA yapısına alkil grubu takan ajanlar ( Kükürt, Nitrojen mustard, Etilenoksitler)")
27
MUTAJENLER d- İnterkalasyon yapan ajanlar :Acridinlerin hepsi bu özelliktedir (Proflawin, acrilflavin ve acridin orange e- Demetilasyon yapan ajanlar: DNA’nın hipo yada demetilasyonuna neden olan ajanlar (5-azacytidine, 5aza-2-deoxycytidine) f- Çeşitli insersiyonlara neden olan ajanlar ( Bu grup ajanlar DNA replikasyonu esnasında-süresince pürin ve primidin bazları yerine DNA yapısına katılan çoğunlukla frame-shift mutasyonlara neden olan ajanlardır.(ethidium bromide-EtBr).
 f- Çeşitli insersiyonlara neden olan ajanlar ( Bu grup ajanlar DNA replikasyonu esnasında-süresince pürin ve primidin bazları yerine DNA yapısına katılan çoğunlukla frame-shift mutasyonlara neden olan ajanlardır.(ethidium bromide-EtBr).")
28
ANTİ MUTASYON MEKANİZMALAR
Ökaryotik hücreler kendileri için zararlı, “LETAL” etki-etkilere sahip mutasyonlara - mutajenlere iki mekanizmayla yanıt verirler. I-Mutasyon önleyici mekanizmalar a- Genomda Junk DNA’nın tutulumu (%98) b- Mutajenlere karşı detoksifikasyon mekanizması c- DNA’nın hücre içi organellerle sınırlandırılması II-Mutasyon giderici mekanizmalar a-Revers mutasyonlar b-Supressör tRNA mutasyonu c-DNA Repair (DNA tamiri) d- Silent mutasyon mekanizması e- Resessif allel sistemi f- Glioksilaz ile eksizyonel tamir
 b- Mutajenlere karşı detoksifikasyon mekanizması. c- DNA’nın hücre içi organellerle sınırlandırılması. II-Mutasyon giderici mekanizmalar. a-Revers mutasyonlar. b-Supressör tRNA mutasyonu. c-DNA Repair (DNA tamiri) d- Silent mutasyon mekanizması. e- Resessif allel sistemi. f- Glioksilaz ile eksizyonel tamir.")
29
mCpG Mutasyonu 1. Deaminasyon CpG UpG CpG 2. Deaminasyon
(+) (Glioksilaz aracılı eksizyonel tamir) 2. Deaminasyon mCpG TpG (tamir yok, hot stop mutasyon) (-) C : Sitozin mC : Metil sitozin, episitozin yada 5mC
 (Glioksilaz aracılı eksizyonel tamir) 2. Deaminasyon. mCpG TpG (tamir yok, hot stop mutasyon) (-) C : Sitozin. mC : Metil sitozin, episitozin yada 5mC.")
30
DNA REPAIR DNA da mutasyon meydana geldikten sonra onu ortadan kaldırmakla yükümlü işlemlerin tümüne DNA repair – DNA tamiri adını alır.Mutasyona uğramış bir DNA molekülü birkaç yolla tamir edilir; I-Direkt Tamir: FOTOREAKTİVASYON - Özellikle DNA yapısında en yaygın meydana gel “timin dimer” mutasyonunun giderildiği tamir mekanizmasıdır. Görünür ışın (güneş ışını) aracılığıyla başarılır. ______________________________ A T G A C A A G ıı ıı ıı ıı ııı ııı Görünür ışın (güneş) T A C T G T = T C Mutant DNA zinciri(Timin dimeri) Fotoreaktivasyon (aktif DNA fotoliaz inaktif DNA fatoliaz) _______________________________ A T G A C A A G ıı ıı ıı ıı ııı ıı ıı ııı Tamir edilmiş DNA zinciri T A C T G T T C Şekil 1. Direkt DNA tamir mekanizmasıyla (fotoreaktivasyon) Timin dimerlerinin tamir edilmesi
 aracılığıyla başarılır. ______________________________. A T G A C A A G. ıı ıı ıı ıı ııı ııı Görünür ışın (güneş) T A C T G T = T C. Mutant DNA zinciri(Timin dimeri) Fotoreaktivasyon (aktif DNA fotoliaz inaktif DNA fatoliaz) _______________________________. A T G A C A A G. ıı ıı ıı ıı ııı ıı ıı ııı Tamir edilmiş DNA zinciri. T A C T G T T C. Şekil 1. Direkt DNA tamir mekanizmasıyla (fotoreaktivasyon) Timin dimerlerinin tamir edilmesi.")
31
II- Eksizyonel tamir: DNA replikasyonunda yanlış eşleşmeler sonunda meydana gelen bazların bir eksonükleaz aktivite ile koparılıp tekrar DNA polimeraz I'in 5'3' polimerizasyon aktivitesiyle doğru bazın eklenmesi esasından ibarettir. Reaksiyonun son basamağında DNA ligaz görev yapacaktır. Ayrıca DNA yanlış eşleşmelerinin tamiri de (mis-match repair) bu grup içerisinde değerlendirilir. İnsanda DNA tamir mekanizmasının bozulduğu, görev yapmadığı durumlarda ortaya çıkan iki yaygın kalıtsal hastalık bilinmektedir. Ekzisyonel tamir üç basamakta başarılır: a- DNA polimeraz I’in yeni replike DNAyı taraması b- Yanlış bazı tanıyıp DNA yı o noktada kırmsı (ekzonükleaz aktivite) c- Doğru bazın transferi(polimeraz aktivite) ve DNA ligaz aktivitesi III- Post-transkripsiyonel tamir: DNA glioksilaz enzimi tarafından yapılan transkripsiyon sonrası tamir mekanizmasıdır. Modifikasyonel mutasyonlar dahil bütün mutasyonlar bu aşamada giderilir. Hastalık Etkili genin lokalizasyonu Klinik tablo 1. Ataxia telangiectasia q lenfoma 2. Xeroderma pigmentosum ERCC 3, 2q deri kanseri ERCC 5, 13q 22-34 Tablo I. DNA tamir mekanizmasının bozulmasına bağlı insanda meydana gelen genetik hastalıklar
 c- Doğru bazın transferi(polimeraz aktivite) ve DNA ligaz aktivitesi. III- Post-transkripsiyonel tamir: DNA glioksilaz enzimi tarafından yapılan transkripsiyon sonrası tamir mekanizmasıdır. Modifikasyonel mutasyonlar dahil bütün mutasyonlar bu aşamada giderilir. Hastalık Etkili genin lokalizasyonu Klinik tablo. 1. Ataxia telangiectasia 11q lenfoma. 2. Xeroderma pigmentosum ERCC 3, 2q21 deri kanseri. ERCC 5, 13q Tablo I. DNA tamir mekanizmasının bozulmasına bağlı insanda meydana gelen genetik hastalıklar")
32
MUTATIONS Wild-type strain (dominant allele)
Mutant strain (recessive allele) DNA RNA Normal PHENOTYPE (wild-type) Correct PROTEIN ( functional enzyme) Mutant DNA Altered RNA Mutant PHENOTYPE Defective PROTEIN (non-functional enzyme)
 DNA. RNA. Normal PHENOTYPE (wild-type) Correct PROTEIN ( functional enzyme) Mutant DNA. Altered RNA. Mutant PHENOTYPE. Defective PROTEIN (non-functional enzyme)")
33
MUTATIONS Wild-type strain (dominant allele)
Mutant strain (recessive allele) DNA Mutant DNA RNA While this is a common scenario, there are many exceptions .e.g.:- some mutants can be dominant some mutants produce functional proteins cause a gain of function, co-dominance etc. Altered RNA Correct PROTEIN ( functional enzyme) Defective PROTEIN (non-functional enzyme) Normal PHENOTYPE (wild-type) Mutant PHENOTYPE
 DNA. Mutant DNA. RNA. While this is a common scenario, there are many exceptions .e.g.:- some mutants can be dominant. some mutants produce functional proteins. cause a gain of function, co-dominance etc. Altered RNA. Correct PROTEIN ( functional enzyme) Defective PROTEIN (non-functional enzyme) Normal PHENOTYPE (wild-type) Mutant PHENOTYPE.")
34
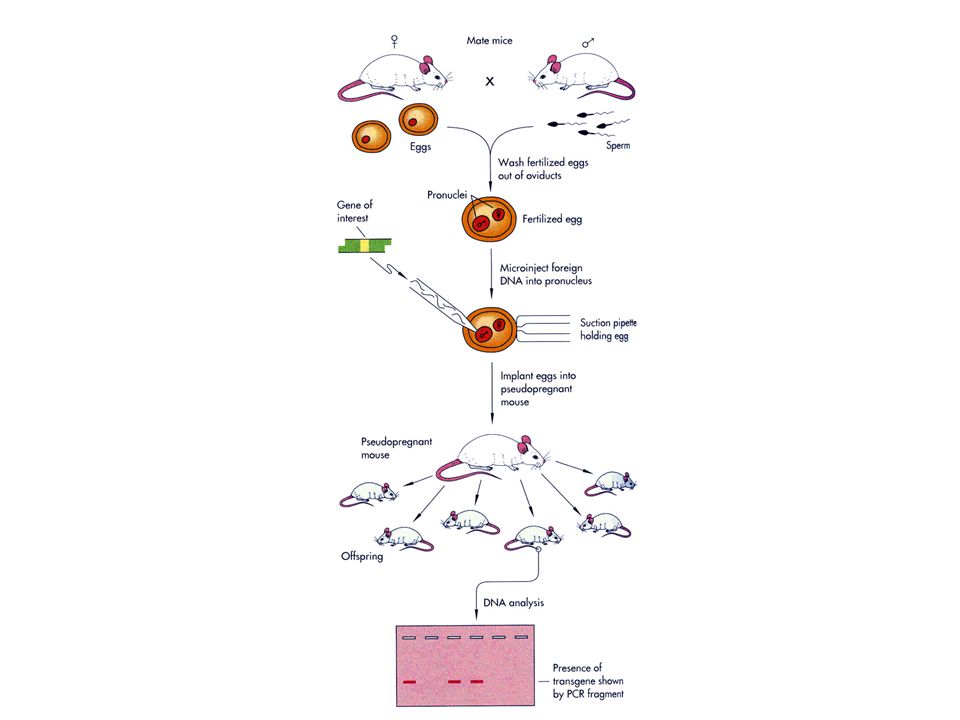
What are Mutations ? Mutations are results of changes to the normal DNA sequence for a gene Typical gene - a linear sequence of about 2000 base pairs AGCCGTGCTGTCGAAAACGTTCAGACTCATTGGCAATCCGAAGTCGGCA TCGGCACGACAGCTTTTGCAAGTCTGAGTAACCGTTAGGCTTCAGCCGT A mutant allele could result from change in only one of them - knocking out the function of that gene AGCCGTGCTGTCGAAAACTTTCAGACTCATTGGCAATCCGAAGTCGGCA TCGGCACGACAGCTTTTGAAAGTCTGAGTAACCGTTAGGCTTCAGCCGT

35
Some types of point mutations
5’ AUG UUA UUA ACU AAG 3’(RNA) met leu leu thr lys (protein) A silent mutation - no effect on phenotype AUG UUA UUG ACU AAG met leu leu thr lys
 met leu leu thr lys (protein) A silent mutation - no effect on phenotype. AUG UUA UUG ACU AAG. met leu leu thr lys.")
36
Some types of point mutations
AUG UUA UGA ACU AAG met leu stop 5’ AUG UUA UUA ACU AAG 3’(RNA) met leu leu thr lys (protein) A missense mutation -- may cause defective protein AUG UUA UUU ACU AAG met leu phe thr lys Changes ‘sense’ of one amino-acid A nonsense mutation -- will make shorter protein
 met leu leu thr lys (protein) A missense mutation -- may cause defective protein. AUG UUA UUU ACU AAG. met leu phe thr lys. Changes ‘sense’ of one amino-acid. A nonsense mutation -- will make shorter protein.")
37
Some types of point mutations
5’ AUG UUA UUA ACU AAG 3’(RNA) met leu leu thr lys (protein) A base substitution mutation AUG UUA UUU ACU AAG met leu phe thr lys An insertion or a deletion (frameshift) AUG UUA UUA ACU AAG met leu leu thr lys U AA G-- stop
 met leu leu thr lys (protein) A base substitution mutation. AUG UUA UUU ACU AAG. met leu phe thr lys. An insertion or a deletion (frameshift) AUG UUA UUA ACU AAG. met leu leu thr lys. U. AA G-- stop.")
38
Some types of point mutations
AUG UUA UUA ACU AAC met leu leu thr asn AUG UUU AUU AAC UAA C met phe ile asn stop ... Insertion of 1 base All amino acids now scrambled from this point on AUG UUU UAU UAA CUA AC met phe tyr stop Insertion of 2 bases All amino acids now scrambled from this point on AUG UUA UUA UUA ACU AAC met leu leu leu thr asn Insertion of 3 bases Amino acids now OK again

39
Summary : types of mutations
Mutations can be Silent - no change to protein Missense - change one a.a. for another Nonsense - cause premature stop signal frameshift - cause scrambled sequence of a.a’s Mutations can be:- Substitutions - change one base for another Insertions/Deletions - gain or loss of a base resulting in frameshifts

40
Base Substitutions A substitution mutation can be…
a transition A G C T purine purine pyrimidine pyrimidine a transversion A T G C purine pyrimidine Transversions are less likely because they result in a change in helix diameter

41
An example: sickle-cell anaemia
HA HS Allele DNA template strand -CTC- -CAC- -GAG- -GUG- mRNA amino acid #6 in chain of hemoglobin -glu- -val- (acidic) (aliphatic)
 (aliphatic)")
42
Chromosomal mutations
So far have been talking about point mutations - changes to individual base pairs. However, other mutations can involve large scale changes to chromosomes Deletions of large sections of a chromosome. Duplications of large sections of a chromosome Inversions (inverted sections of a chromosome). Translocations (exchanges of sections of non-homologous chromosomes) Transposons - bits of DNA that suddenly ‘jump’ to a new location - also knock out genes and cause mutation
. Translocations (exchanges of sections of non-homologous chromosomes) Transposons - bits of DNA that suddenly ‘jump’ to a new location - also knock out genes and cause mutation.")
43
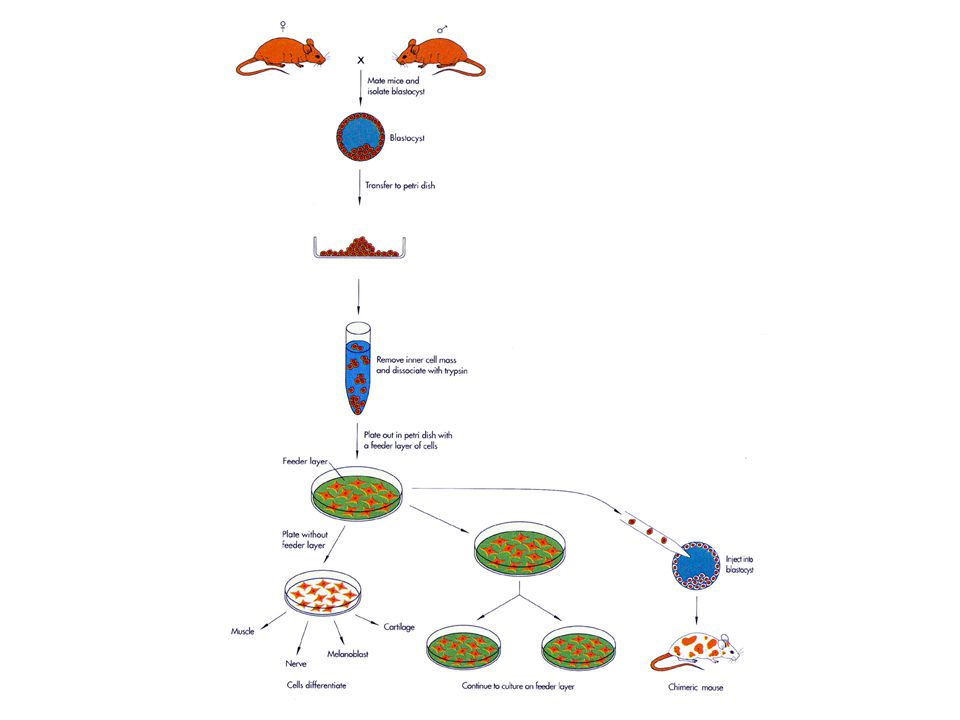
SOMATİK HÜCRE KALITIMI (epigenetik kalıtım) Doç.Dr.Öztürk ÖZDEMİR
“Aynı organizmaya ait hücrelerarası gen aksiyon farklılığını inceleyen genetik alt dalı”

44
GENETİK DÜZENLEMEDE – SOMATİK KALITIM EVRELERİ
1- Yumurta hücresi düzeyinde düzenleme : Yumurta hücresinde bulunan anterioposterior gradiyent farkı fertilizisyon öncesi yumurta hücresinden meydana gelecek embriyonun anteriyor ve posteriyor kısmını verecek bölgeler öncelikle belirlenmektedir. Burada sadece yumurta hücresiyle sınırlı bazı regülatör-modülatör proteinler yumurta-polarity ve segmentasyondan sorumlu (25 adet tanımlanmıştır) genler görev almaktadırlar. 2. Zigot evresinde düzenleme : Bu evrede yine çoğu yumurtadan orijin alan ve döllenmeyi takiben aktive olan zigotik-effekt genler olarak bilinen; remodelling faktörler, integrinler, transkripsiyonel faktörler ve kromatin bağlayıcı özgül proteinler gibi düzenleyici moleküllerin görev aldıkları saptanmıştır. Bu genlerin görevi yumurta ve sperm çekirdeklerinin kaynaşmasını sağlamak ve hücre bölünmesi öncesi görev yapan proteinlerin bazı regülasyonunda görev alırlar.
 genler görev almaktadırlar. 2. Zigot evresinde düzenleme : Bu evrede yine çoğu yumurtadan orijin alan ve döllenmeyi takiben aktive olan zigotik-effekt genler olarak bilinen; remodelling faktörler, integrinler, transkripsiyonel faktörler ve kromatin bağlayıcı özgül proteinler gibi düzenleyici moleküllerin görev aldıkları saptanmıştır. Bu genlerin görevi yumurta ve sperm çekirdeklerinin kaynaşmasını sağlamak ve hücre bölünmesi öncesi görev yapan proteinlerin bazı regülasyonunda görev alırlar.")
45
3. Gastrulasyon-Embriyogenez evresinde düzenleme : Bu evrede görev alan en önemli gen grubunun yine yumurta hücresine ait 8 çift oldukalı saptanan pair-rule ve 10 adet oldukarı saptanan segment polarity genlerdir. Bu gen grubu her tür için farklı olmakla birlikte embriyogenezin 2, 8 ve 16 hücrelik bölünme evrelerinde inaktive edilirler. Örneğin farelerde zigot 2, koyun ve insanda 16 hücrelik embriyo olana kadar görev yapmaktadırlar. Bu sayı türe göre değişmektedir. Zigot hücresinin maksimum 16 hücreye kadar bölünmesinden sorumlu gen grubudur. Bu genler sadece totipotent hücrelerde görev alırlar. 4. Fetus dönemi düzenleme : Bu dönem, fetus hücrelerine ait genlerin ifade edilmesiyle başlar. Bu dönemden sonra yumurta regülasyonu yerini fetus gen regülasyonuna terk eder. Görev yapan genler homeotik ya da homeodometik (hox) gen ailesi olarak adlandırılır. Diğer bir tanımla bu genler geniş bir aileden ibaret olup, yetişkin dokuların ilkin farklılaşmasından sorumlu oldukları için homeotik seçici genler olarak adlandırılırlar. Türlerarası somatik doku farklılaşmasından birinci dereceden bu gen grubu sorumludur. Bir dokunun normal ya da anormal bir şekilde farklılaşması bu gen grubunun normal ve zamanında fonksiyon yapmasına bağlıdır. İlk kez blastoderm evresinde aktive olurlar. Memelilerde 4 adet homolog homeotik kompleks genin varlığı saptanmıştır. Homeodometik seçici genleri meydana getiren homeodomain zincir genleri evrim süresince korunan ve en az varyasyon gösteren genlerdir. Herbiri 60 aa uzunluğunda protein sentezinden sorumlu bç uzunluğunda regülatör alt birimlerinden ibaret genlerdir. Genlerin regülatör alt birimlerini meydana getiren diziler, segmentasyonel ve yumurta -polarity gen ürünlerine özgül bağlantı bölgeler içerirler. Vücudun segmentasyonunda spesifik görev yapan bu gen grubudur. Moleküler mekanizmaları kesin olarak bilinmemekle birlikte regülatör alt birimleri aracılığıyla aktive ve inhibe edildikleri sanılmaktadır.
 gen ailesi olarak adlandırılır. Diğer bir tanımla bu genler geniş bir aileden ibaret olup, yetişkin dokuların ilkin farklılaşmasından sorumlu oldukları için homeotik seçici genler olarak adlandırılırlar. Türlerarası somatik doku farklılaşmasından birinci dereceden bu gen grubu sorumludur. Bir dokunun normal ya da anormal bir şekilde farklılaşması bu gen grubunun normal ve zamanında fonksiyon yapmasına bağlıdır. İlk kez blastoderm evresinde aktive olurlar. Memelilerde 4 adet homolog homeotik kompleks genin varlığı saptanmıştır. Homeodometik seçici genleri meydana getiren homeodomain zincir genleri evrim süresince korunan ve en az varyasyon gösteren genlerdir. Herbiri 60 aa uzunluğunda protein sentezinden sorumlu bç uzunluğunda regülatör alt birimlerinden ibaret genlerdir. Genlerin regülatör alt birimlerini meydana getiren diziler, segmentasyonel ve yumurta -polarity gen ürünlerine özgül bağlantı bölgeler içerirler. Vücudun segmentasyonunda spesifik görev yapan bu gen grubudur. Moleküler mekanizmaları kesin olarak bilinmemekle birlikte regülatör alt birimleri aracılığıyla aktive ve inhibe edildikleri sanılmaktadır..")
46
5-YETİŞKİN (ADULT) DÖNEM DÜZENLEME
1- Housekeeping genler 2- Doku spesifik genler 3- Alel spesifik genler 4- Diğer (ekspresyon farklılığı gösteren genler, onkogenler, TS genler vb)
")
47
EPİGENETİKTE (SOMATİK KALITIM) ETKİLİ MEKANİZMALAR
I- DNA METİLASYONU II- FOSFORİLASYON III- ASETİLASYON IV- UBİQUTİNASYON V- HIGH MOBIL NON-HISTON PROTEİNLER

48
DNA METİLASYONU - Hücre farklılaşması - Gen ekspresyonu - Yaşlanma
- DNA replikasyonunun başlatılması - DNA transkripsiyonunun başlatılması - DNA tamiri - Mutagenezis - İkili sarmal DNA stabilitesinin sağlanması - Lokal mutasyon oranının artırılması - Nükleer parçalanmanın engellenmesi - Kromozom paketlenmesi - Hücre farklılaşması - X-kromozom inaktivasyonu - Gen ekspresyonu - Yaşlanma - Tümör baskılayıcı gen inaktivasyonu ve proto-onkogen aktivasyonu aracılı onkogenezis - Genomun aktif gen ya da kondanse bölgeler şeklinde yapılanması ve yerleşimi - Apoptozis

49
DNA METİLASYONU Post-replikatif bir mekanizmadır
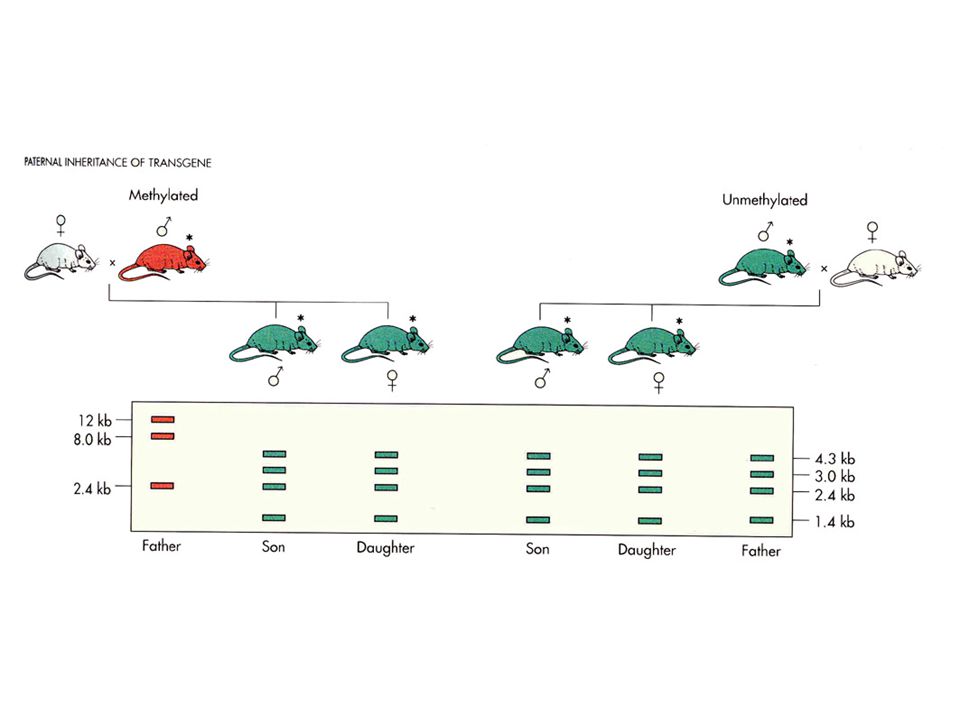
DNA düzeyinde yapılan modifikasyonla karakterize epigenetik mekanizmadır DNA metiltransferaz görev alır İnsanda % 90 oranında metillenenz nükleotidler mCpG dinükleotididir. DNA metilasyon oranı açısından; Ametile DNA (Ökromatik DNA, Housekeeping genler) Hipometile DNA (Fakültatif heterokromatik DNA, Pseudogenler, inaktif X) Undermetile DNA (bazı onkogenler) Metile DNA (Heterokromatik DNA, İnterkalar heterokromatik DNA, protoonkogenler) Hipermetile DNA (Sentromerik DNA, İnaktik Junk DNA) mC, 5-metilsitozin yada episitozin olarak adlandırılır Semikonservatif kalıtılır
 Hipometile DNA (Fakültatif heterokromatik DNA, Pseudogenler, inaktif X) Undermetile DNA (bazı onkogenler) Metile DNA (Heterokromatik DNA, İnterkalar heterokromatik DNA, protoonkogenler) Hipermetile DNA (Sentromerik DNA, İnaktik Junk DNA) mC, 5-metilsitozin yada episitozin olarak adlandırılır. Semikonservatif kalıtılır.")
50
ELEMANLARI Metil vericisi SAM (S adenozil methionin)
- Substrat template DNA Enzim DNA Metil transferaz SAM metil grubunu kaybedince SAH (S adenozin homosistein)’e dönüşür. Ökaryot ve prokaryot hücrelerin herikisinde en yaygın metillenen baz sitozin ( C) dir. Prokaryotlarda CCGG dizilerindeki ilk sitozin, ökaryotlarda ise CpG dinükleotidlerdeki ilk sitozin en yaygın metillenen bazdır. DNA yapısından metil grubunun koparılmasında görev alan enzim DNA Mtaz dır.
’e dönüşür. Ökaryot ve prokaryot hücrelerin herikisinde en yaygın metillenen baz sitozin ( C) dir. Prokaryotlarda CCGG dizilerindeki ilk sitozin, ökaryotlarda ise CpG dinükleotidlerdeki ilk sitozin en yaygın metillenen bazdır. DNA yapısından metil grubunun koparılmasında görev alan enzim DNA Mtaz dır.")
51
METİLASYONUN ONKOGENEZDE ÖNEMİ
Bütün onkogenler ökaryotik hücrelerde öncelikle DNA düzeyinde modifiye edilerek inaktive dilir DNA metisyonu görev alır Onkogenler hipermetile durumda inaktif durumdadırlar Onkogen hipermetile ya /yada metillenerek inaktive edilir, protoonkogene dönüştürülür. Fosforilasyon, ubiqutinasyon, yüksek mobiliteye sahip non-histon proteinlerin varlığı ve asetilasyon ise nükleoproteinler düzeyinde (histon –non-histon) yapılan epigenetik modifikasyon mekanizmaları olur gen ekspresyonu farklılaşmasında rol alan en önemli mekanizmalardır. Tümör supressör (20 adet) genler ÖR : p53 DNA hipermetilasyonu sonucu ekspresiyonel olarak inaktive edilir, hücre onkogeneze girer. Bu genler normal hücrelerde aktif genlerdir, inaktif durumda hücrede kansere neden olurlar. Genlerin inaktivasyonları da DNA hipermetilasyonu ile olur.
 yapılan epigenetik modifikasyon mekanizmaları olur gen ekspresyonu farklılaşmasında rol alan en önemli mekanizmalardır. Tümör supressör (20 adet) genler ÖR : p53 DNA hipermetilasyonu sonucu ekspresiyonel olarak inaktive edilir, hücre onkogeneze girer. Bu genler normal hücrelerde aktif genlerdir, inaktif durumda hücrede kansere neden olurlar. Genlerin inaktivasyonları da DNA hipermetilasyonu ile olur.")
52
ASETİLASYON 1- DNP düzeyinde yapılan bir modifikason şeklidir.
2- H3 ve H4 en yaygın asetillenen histon proteinelerdir 3- N-asetil lizin en sık asetillenen aa dir. 4- Asetillasyon gen aksiyonu ile doğru ilişiktedir. 5- Kromozomların “aktif gen” bölgelerindeki H3 ve H4 proteinlerine ait lizin aa leri hiper asetillenmiş formdalar.

53
UBİQUİTİNASYON Histon proteinlerin C (Karboksil) terminaline yakın lizin aminoasitlerine ubiquitin adında küçük proteinlerin aktarılması ile karakterize modifikasyon şeklidir. En sık H2A ve H2B ubiquitine edilir. Ubiquitine H2A toplam histon proteinlerin % 10 ve Ubiquitine H2B ise toplam histon proteinlerin %1-2’sini teşkil eder. Ubiquitine proteinler, stoplazmada yapılan protein parçalanmalarında sinyal görevi yapmaktadırlar. Mitoza giren hücrelerde bölünme süresince kromatin fibrilinin 30 ºA çapta kalmasında görev yapabileceği sanılmaktadır. Transkripsiyonel aktif gen bölgelerinde ubiquitine histon proteinlerin yaygın oldukları, gen regülasyonunda görev alabilecekleri sanılmaktadır.
 terminaline yakın lizin aminoasitlerine ubiquitin adında küçük proteinlerin aktarılması ile karakterize modifikasyon şeklidir. En sık H2A ve H2B ubiquitine edilir. Ubiquitine H2A toplam histon proteinlerin % 10 ve. Ubiquitine H2B ise toplam histon proteinlerin %1-2’sini teşkil eder. Ubiquitine proteinler, stoplazmada yapılan protein parçalanmalarında sinyal görevi yapmaktadırlar. Mitoza giren hücrelerde bölünme süresince kromatin fibrilinin 30 ºA çapta kalmasında görev yapabileceği sanılmaktadır. Transkripsiyonel aktif gen bölgelerinde ubiquitine histon proteinlerin yaygın oldukları, gen regülasyonunda görev alabilecekleri sanılmaktadır.")
54
HİGH MOBİL NON-HİSTON PROTEİNLER
1- Dört tipi vardır 2- Tamamı non-histon proteinelerden ibarettir. 3- Non mutabıldırlar (mutasyonu tolere edemezler) 4- Polipeptit zincir yapısında asimetrik bulunurlar 5- Yüksel mobil aktiviteye sahiptirler, DNA zinciri boyunca kolay hareket etme, ve pozisyon değiştirme yeteneğindedirler. 6- DNA ya asimetrik yerleşirler 7- Dizi özgüllüğü gösterirler 8- Yüklü [ (+), (-)] aminoasitlerce zengindirler
 4- Polipeptit zincir yapısında asimetrik bulunurlar. 5- Yüksel mobil aktiviteye sahiptirler, DNA zinciri boyunca kolay hareket etme, ve pozisyon değiştirme yeteneğindedirler. 6- DNA ya asimetrik yerleşirler. 7- Dizi özgüllüğü gösterirler. 8- Yüklü [ (+), (-)] aminoasitlerce zengindirler.")
55
HİGH MOBİL NON-HİSTON PROTEİNLER
I- Kromatin bağlayıcı enzimler: histon proteinlerin post-transkripsiyonunda görev alırlar DNA tamir sentez replikasyon nükleazlar proteazlar bağlayıcı motifler metilazlar ubiquitin transferaz fosfor-fosfat transferaz ADP ribozil transferaz

56
HİGH MOBİL NON-HİSTON PROTEİNLER
II- HMGP Proteinler Tipi bağlandığı bölge olası görevi HMGP1/2 proteinler interkalar DNA DNA replikasyonu ve tamir - genel transkripsiyon faktörü - DNA loop stabilizasyonu HMGP 14/ nükleozom transkripsiyon başlama noktaları HMGP 1/Y özgül diziler kromatin kondensasyonu A/T ce zengin bölgeler genel transkripsiyon faktörü - gen amplifikasyonu

57
HİGH MOBİL NON-HİSTON PROTEİNLER
III- Transkripsiyon faktörler lösin zipper zink finger helix loop helix HMGP adaptör proteinler helix turn helix

58
HİGH MOBİL NON-HİSTON PROTEİNLER
IV- Kromozom yapıcı proteineler - kromomerler - diğer quaterner birim elemanları

59
HÜCRE ÖLÜM MEKANİZMALARI
Apoptozis Nekrozis Sitotoksisite

60
Tablo I. Apoptozda etkili basamaklara genel bakış.
Tablo I. Apoptozda etkili basamaklara genel bakış. Uyarıcılar Upstream Caspase Aktivasyonu Mitokondriyal membranında potansiyal kayıp ROS üretiminde artış Asidifikasyon Downstream Caspase Aktivasyonu Kromatin condensasyonu Fosfatidilserin translokasyonu Hücre membran permeabilitesinde artış DNA fregmantasyonu Apoptatik body oluşumları Fagositozis (ölüm)
")
61
APOPTOZİS Kontrol edilen – proğramlı hücre ölümüdür
Fizyolojik bir process olup istenmeyen yada yararsız hücrelerin ölümünden sorumlu mekanizmadır. yüksek canlılarda özellikle gelişme ve doku farklılaşması dönemlerinde görev yapar”fiyolojik apoptozis”. Farklılaşmasını tamamlamış doku –hücrelerde meydana gelirse “patolojik apoptozis “adlandırılır.

62
APOPTOZİS EVRELERİ Membran blebbing
Kromatin(çekirdek)kompertmentalizasyonu Sitoplazma kondensasyonu DNA fragmentasyonu Mitokondri membran yapı bozukluğu Apoptotik body oluşumu Fagositozis
kompertmentalizasyonu. Sitoplazma kondensasyonu. DNA fragmentasyonu. Mitokondri membran yapı bozukluğu. Apoptotik body oluşumu. Fagositozis.")
63
APOPTOZİS 1- Özel grup hücrelerde meydana gelir
2- Hormonal değişim ve büyüme faktörlerinin yokluğu gibi fizyolojik sitimülasyona bağlı gelişir 3- Apoptotik body ler makrofaj ve diğer komşu hücrelerce fagosite edilir 4- İnflamasyonel yanıt görülmez.

64
APOPTOZİS ÖZELLİKLERİ
Enzimatik basamakları düzenlenebilen mekanizmadır 37 C’de meydana gelen ATP bağımlı bir mekanizmadır. Agaroz elektroforezde “ladder” yapı gösterir Mitokondri membran değişiklikleri mevcuttur: -fosfatidilserin translokasyonu -AIF ve sitokrom C sekresyonu

65
APOPTOZİS GÖREVLERİ Embriyogenezis Doku homeostazisi İmmün tolerans
Sinir hücrelerinin gelişimi Normal hücre gelişimi Endokrin bağımlı doku atrofisi Primer gonad - seks gelişimi Metamorfozis

66
APOPTOZİS TETİK ÇEKİCİLERİ
Hücre yüzey reseptör ölümleri (CD95, APO 1, Fas ve ras aktivasyonu) Fosfatidilserin translokasyonu, extrinsik matiriks değişimi 11 farklı intrasellüler Cystein proteaz enzimlerin sitozole salınımı (Caspase 8 ve 9) AIF salınımı Ca ve Mg bağımlı oligonükleozomal endonükleaz aktivasyonu
 Fosfatidilserin translokasyonu, extrinsik matiriks değişimi. 11 farklı intrasellüler Cystein proteaz enzimlerin sitozole salınımı (Caspase 8 ve 9) AIF salınımı. Ca ve Mg bağımlı oligonükleozomal endonükleaz aktivasyonu.")
67
NEKROZİS Kazaen hücre ölümüdür Patolojik bir process tir
İstenmeyen hücre ölüm mekanizmasıdır Hücrenin çok ciddi bir fiziksel yada kimyasal ajanlara maruz kaldığı durumda kendi siteği dışında gelişen bir ölüm mekanizmasıdır. İnflamasyonel yanıt mevcut (yangı) Makrofajlarla fagositozis görülür Homeostazis yokluğu en önemli etkendir
 Makrofajlarla fagositozis görülür. Homeostazis yokluğu en önemli etkendir.")
68
NEKROZİS ÖZELLİKLERİ Vesikül oluşumu görülmez
Homeostazis regülasyonu ortadan kalkmıştır Enrji gereksinimi yoktur Hücre özgüllüğü yoktur “Smear DNA” yapısına sahiptir + 4 C’de meydana gelir Hücre ölümünün son basamağında rastgele DNA parçalanması görülür Vesikül oluşumu görülmez Smoot mitokondri ve hücre membran yapısı Sitoplazma vemitokondri membran yapısında irreversible swelling (şişme) Total hücre ölümü ile sonlanır
 Total hücre ölümü ile sonlanır.")
69
NEKROZİS ETKENLERİ metabolik zehirlenmeler ischemia hipoksi Hipertermi
litik viruslar complemen ataklar - homeostasis gerilemesi(hücreye su ve iyon geçişinde düzensizlikler)
")
70
SİTOTOKSİSİTE İlaç Kozmetikler Çeşitli yiyecekler
Ağır kimyasal bileşenler gibi toksik etkenlerin neden olduğu hücre ölüm mekanizmasıdır. Patolojik bir mekanizmadır T-hücreler aracılı fagositozis bu mekanizmaya dahil edilir MHC reaksiyonların tamamı sitototoksisite ölümdür Apoptotik body fagositozu yine bir sitotoksisite ölümdür olarak kabül görür.

Benzer bir sunumlar













>")








 Doç.Dr.Öztürk ÖZDEMİR>")
