Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Amino asit-Peptid ve Proteinler 1
Prof Dr A Binnur Erbağcı Tıbbi Biyokimya AD.

2
Proteinler amino asit polimerleridir
Proteinler, canlı hücrelerde en fazla bulunan makromoleküller; yapı taşları olan amino asitlerin özgün kovalent bağlarla bağlanarak oluşturdukları zincirlerden meydana gelirler. ribozom

3
Amino asitler Doğada 400 kadar farklı amino asit bulunmasına rağmen...
Proteinler aynı 20 amino asitin farklı kombinasyonlarından meydana gelir. Protein yapısında bulunan amino asitlere standart veya primer amino asitler denilir. Bu 20 aa’i kodlayan özel genetik kodlar (kodonlar bulunur). 20 farklı amino asit : protein çeşitliliğinin üst düzeyde olmasını sağlar
. 20 farklı amino asit : protein çeşitliliğinin üst düzeyde olmasını sağlar.")
4
2 amino asitten meydana gelen bir peptid:
AA1 AA2 x = farklı molekül AA1 AA2 AA3 20 x x = farklı molekül 100 amino asitten meydana gelen protein için olasılık Evrendeki toplam atom sayısı

5
Protein: H, C, N, O, S atomlarından meydana gelirler

6
Amino asitlerin ortak yapısal özellikleri
Aynı C atomuna bağlı H atomu Karboksil (-COOH) grubu Amino (-NH2) grubu R grubu : değişken grup, yapısı, elektrik yükü ve büyüklüğü farklılık gösterir
 grubu. Amino (-NH2) grubu. R grubu : değişken grup, yapısı, elektrik yükü ve büyüklüğü farklılık gösterir.")
7
R NH2 -C-COOH H R NH2 -C-COOH H
Amino asitlerin ortak yapısal özellikleri asit NH2 -C-COOH R H NH2 -C-COOH R H Ortak yapı amino

8
C atomlarının adlandırılması
+NH3-CH-COO- CH2 COO-

9
C atomlarının adlandırılması
Bu amino asitlere (alfa) amino asitler denilir çünkü fonksiyonel grubun (-NH2) bağlandığı C atomu –COOH grubuna göre alfa pozisyonundadır. R grubunda varsa diğer C atomları sırası ile , , , C atomları olarak adlandırılır.
 amino asitler denilir çünkü fonksiyonel grubun (-NH2) bağlandığı C atomu –COOH grubuna göre alfa pozisyonundadır. R grubunda varsa diğer C atomları sırası ile , , , C atomları olarak adlandırılır.")
10
The amino group can be located on the hydrocarbon chain with a Greek letter, , , , or Figure 25.1 α β

11
Amino asitler tetrahedral yapılardır

12
Hatırlayalım;İzomerler
Kimyasal formülü AYNI ama atomların düzenlemeleri farklı olan moleküllere izomerler denilir. Yapısal izomerler: atomların birbirine bağlanışı farklıdır. Stereo izomerler bir C atomuna bağlı 4 grup birbirinden farklı olduğunda meydana gelir. Stereo izomerler; benzer veya aynı kimyasal özellikler ama farklı (bazı) biyolojik özellikler
 biyolojik özellikler.")
13
Amino asitlerin ortak özellikleri-1
Glisin dışında tüm standart aa’lerin C atomları asimetriktir yani 4 farklı grup bağlanmıştır. Böyle C atomlarına kiral merkez denilir. Bu moleküllerin ayna görüntüsü molekül üzerine tam olarak oturmaz.

14
Properties of Molecules

15
Amino asitlerin ortak özellikleri-2
Kiral merkezi olan tüm aa’ler aynı zamanda optikçe aktiftir. Yani polarize (doğru halinde) ışığın yönünü sağa veya sola çevirirler, Glisin dışında tüm amino asitler optikçe aktiftir.
 ışığın yönünü sağa veya sola çevirirler, Glisin dışında tüm amino asitler optikçe aktiftir.")
16
Amino asitlerin ortak özellikleri-3
Aa’lerin 2n kadar izomeri vardır. n: asimetrik C atomu sayısı (kiral merkez) n=1 ise, izomer=2 İzomerlerin adlandırılması: gliseraldehitin –OH grubu örnek alınarak yapılır.
 n=1 ise, izomer=2. İzomerlerin adlandırılması: gliseraldehitin –OH grubu örnek alınarak yapılır.")
17
Amino asitlerde ise -NH2 grubu izomerizmi belirler
Amino asitlerde ise -NH2 grubu izomerizmi belirler. -NH2 grubu sağda ise D-aa, solda ise L-aa denilir.

19
Protein yapısında L-aa’ler vardır
Protein yapısında yer alan aa’lerin tümü L-aa’lerdir. Proteinlerin karekteristik 3 boyutlu yapıları bileşenleri olan aa’lerin aynı sterokimyasal yapıda olmasını gerektirir. Protein sentezinde rol alan enzimler sadece L-aa’leri yapısına katar.

20
Amino asitlerin ortak özellikleri-4
Sulu çözeltilerde aa’lerin amino ve karboksil grupları ortam pH’sına göre iyonize olarak asit veya baz gibi davranabilirler: Bu özellikteki moleküllere amfoterik moleküller denilir.

21
Amino asitlerin iyonizasyonu
Sulu çözeltilerde iyonize olabilen karboksil ve amino gruplarının iki formu protonik denge halinde bulunur R-COOH R-COO- + H+ (pK1) R-NH3+ R-NH2 + H+ (pK2) pK2 pK1
 R-NH3+ R-NH2 + H+ (pK2) pK2. pK1.")
22
Amino asitlerin iyonizasyonu
R-COOH R-COO- + H+ (pK1) R-NH3+ R-NH2 + H+ (pK2) R-COOH ve R-NH3+ protonlu veya zayıf asidik bileşenlerdir. R-COO- ve R-NH2 konjuge bazlar yani proton alıcılarıdır. pK arasında değiştiğinden karboksil grupları nötral pH’ta tüme yakın iyonizedir (COO- ) pK2 ise arasında olduğundan amino gruplarıda nötral pH’ta tüme yakın iyonizedir (NH3+).
 R-NH3+ R-NH2 + H+ (pK2) R-COOH ve R-NH3+ protonlu veya zayıf asidik bileşenlerdir. R-COO- ve R-NH2 konjuge bazlar yani proton alıcılarıdır. pK arasında değiştiğinden karboksil grupları nötral pH’ta tüme yakın iyonizedir (COO- ) pK2 ise arasında olduğundan amino gruplarıda nötral pH’ta tüme yakın iyonizedir (NH3+).")
23
Fizyolojik pH’ta tüm aa’ler hem pozitif hem de negatif yük taşırlar
Böyle moleküllere zwitteriyon denilir.

24
Amino asitler tampon görevi yapabilir
Amino asitlerin titrasyon eğrisi incelendiğinde amino, karboksil gruplarının ve eğer varsa R gruplarındaki iyonize olabilen yapıların pKa’ları ile uyumlu pH’ta tamponlama yapabildiği görülür. Bu eğriler yardımı ile karboksil ve amino gruplarının pKa’ları saptanabilir.

25
NH3+ COOH COO- NH2 100% pH = 5.97

26
NH3+ COOH COO- NH2 50% pH = 5.97

27
Birinci proton/hidrojen’in ayrışması tamamlanmıştır.
COOH COO- NH2 100% pH = 5.97 Birinci proton/hidrojen’in ayrışması tamamlanmıştır.

28
NH3+ COOH COO- NH2 50% pH = 5.97

29
NH3+ COOH COO- NH2 100% İkinci proton / hidrojen’in ayrışması tamamlanmıştır. pH = 5.97

30
R NH2 -C-COOH H bu yapı hangi pH’ta olur? Cevap: hiçbir pH
Karboksil grubu amino grubundan daha kuvvetli bir asittir (pKa’ya bak)ve protononu daha önce kaybeder. NH2 -C-COOH R H asidik pH’ta COOH , NH3+ nötral pH’ta COO- , NH3+ bazik pH’ta COO- , NH2
ve protononu. daha önce. kaybeder. NH2 -C-COOH. R. H. asidik pH’ta COOH , NH3+ nötral pH’ta COO- , NH3+ bazik pH’ta COO- , NH2.")
31
Amino asitlerin izoelektrik pH’ı (pI)
İzoelektrik pH bir molekülün toplam yükünün 0 olduğu yani pozitif ve negatif yüklerin birbirine eş olduğu pH’tır. Sadece 2 iyonize olabilen grup taşıyan aa’lerde pI= pK1 + pK2 /2 2’den fazla iyonize olan grup varsa pI toplam yükün 0 olduğu forma komşu iki pK’nın ortalamasıdır.

33
amino asitlerin kodları
Alanin Ala A Sistein Cys C Aspartik asit Asp D Glutamik asit Glu E Fenil alanin Phe F Glisin Gly G Histidin His H Isolösin Ile I Lizin Lys K Lösin Leu L Metyonin Met M Asparagin Asn N Prolin Pro P Glutamin Gln Q Arginin Arg R Serin Ser S Threonin Thr T Valin Val V Triptofan Trp W Tirozin Tyr Y standart aa’lerin Birisi 3 harfli, Diğeri tek harfli 2 ayrı kısa kodu vardır.

34
Karbonil Hidroksil Metil

35
O NH NH2 Amino C C Amid CH3

36
karboksil fenil

37
Imidazol

38
Sulfidril

39
N H +NH3-CH-COO- CH2 İndol

40
NH C NH2+ NH2 Guanido grubu

41
Amino asitlerin sınıflaması-1 (R gruplarına göre)
Alifatik Hidroksilik Sülfürlü Asidik R grupları ve amidleri Bazik R grupları Aromatik İmino asit

42
Alifatik aa’ler-1 R grupları hidrokarbon yapıdadır.
Valin, lösin ve izolösine topluca dallı zincirli aa’ler denilir. Dallı zincirli aa’ler non-polar yapıda ve hidrofobiktir (biyolojik pH‘ta su ile etkileşme eğilimleri düşüktür) Glisin ve Alanin R grubunun küçük olması (H atomu ve metil grubu) nedeni ile polardır ve hidrofobik değildir. Glisin Alanin Valin Lösin izolösin
 Glisin ve Alanin R grubunun küçük olması (H atomu ve metil grubu) nedeni ile polardır ve hidrofobik değildir. Glisin. Alanin. Valin. Lösin. izolösin.")
43
glisin Gly G alanin Ala A valin lösin Leu L İzo-lösin Ile I 2.4 9.8
9.9 valin Val V 2.2 9.7 lösin Leu L 2.3 İzo-lösin Ile I

44
Alifatik aa’ler-2 Bu aa’ler (glisin hariç) protein yapılarının hidrofobik etkileşimlerinden sorumludurlar. Glisin R grubunun küçük olması nedeni ile proteinin o noktasına bükülebilme özelliği kazandırır.

45
Hidroksilli aa’ler Serin Treonin Tirozin
-OH grupları bazı enzimlerin aktif merkezinde bulunur ve katalitik aktivitenin düzenlenmesinde yer alır. Bazı enzimler ve proteinler fosforile/defosforile olarak aktifleşirler. Proteinlerde fosforilasyon hidroksilli aa’lerin R grupları üzerinden olur. Serin ve treonin, proteinlere karbohidrat eklenme noktasıdır. Serin Treonin Tirozin

46
Serin Ser S 2.2 9.2 13 treonin Thr T 2.1 9.1 tirozin Tyr 10.1

47
Sülfürlü amino asitler-2
S atomu aa/lerin yapısında iki farklı şekilde bulunabilir. 1) Metiyonin aa’i metil grubu vericisi olan SAM (S-adenozil metiyonin) yapısında bulunur. S atomu metil grubunun kolaylıkla verilmesini sağlar. 2) -SH grubu şeklinde sistein yapısında bulunur. –SH bazı enzimlerin aktif merkezinde katalitik rol oynar Sistein Metiyonin
 Metiyonin aa’i metil grubu vericisi olan SAM (S-adenozil metiyonin) yapısında bulunur. S atomu metil grubunun kolaylıkla verilmesini sağlar. 2) -SH grubu şeklinde sistein yapısında bulunur. –SH bazı enzimlerin aktif merkezinde katalitik rol oynar. Sistein. Metiyonin.")
48
Sistein Cys C 1.9 10.8 8.3 Metiyonin Met M 2.1 9.3

49
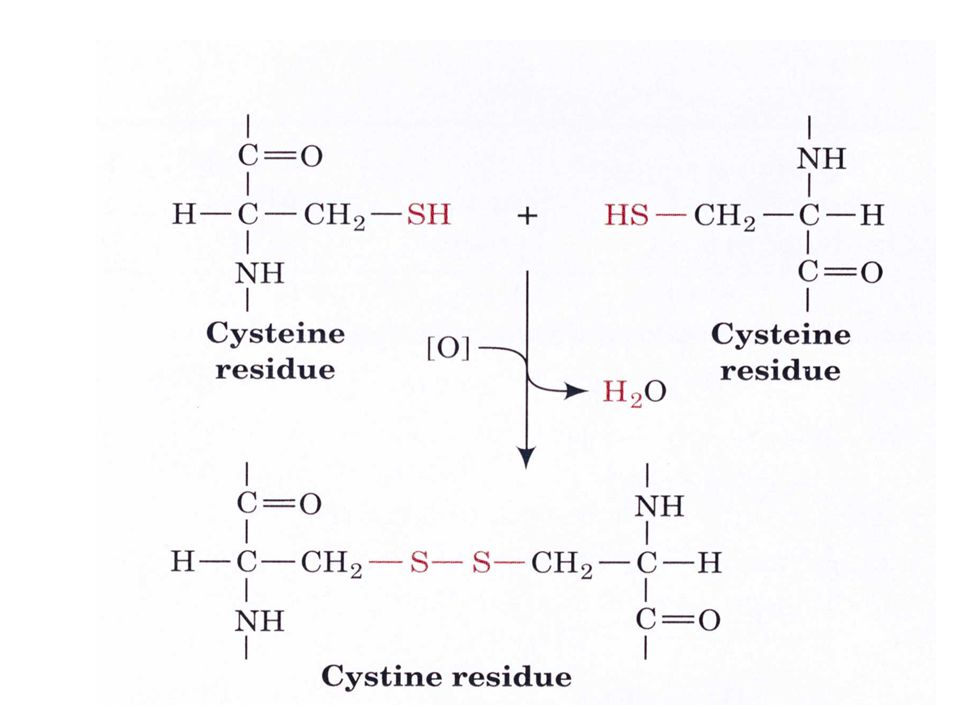
Sülfürlü amino asitler-2
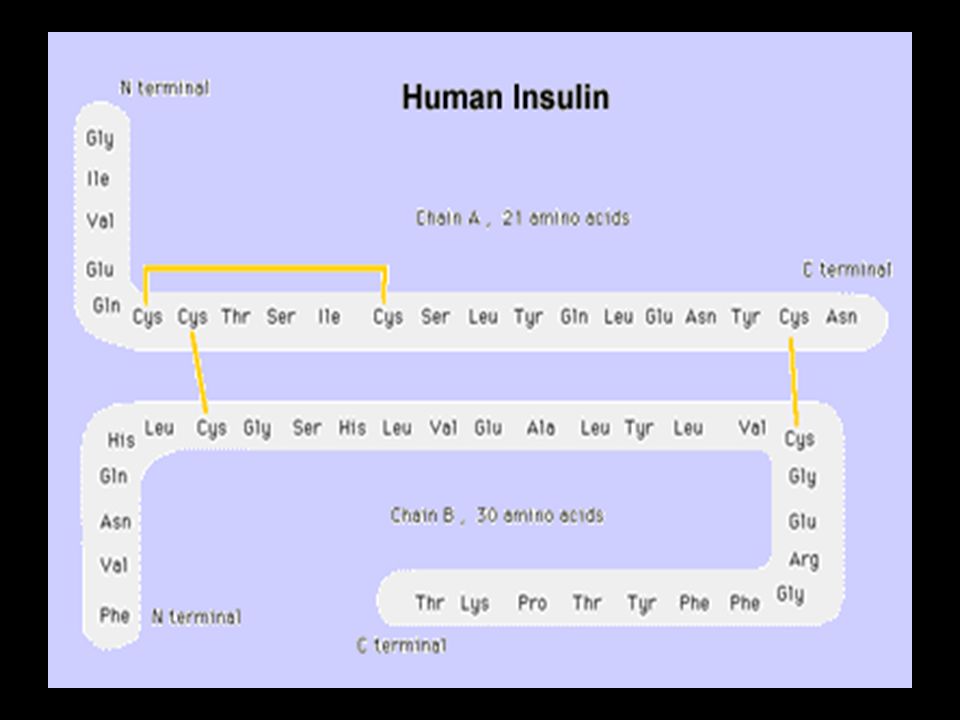
İki sistein molekülü okside olarak dimerik bir aa olan sistini oluşturur. Bu yapıya disülfit bağı adı verilir. Disülfit bağları hücre dışı proteinlerin stabilizasyonunda rol alan kovalent bağlardır. Sistin keratin adı verilen epitel hücreleri (deri), tırnak ve saç proteinin yapısında bulunur.
, tırnak ve saç proteinin yapısında bulunur.")
52
Asidik yan gruplar Asidik aa/lerin R gruplarında ilave bir –COOH grubu bulunur. pH 7’de net negatif yüke sahiptirler. Bazik ve asidik aa’lerin yan grupları iyonik bağlar oluşumu ile spesifik protein konformasyonlarını stabilize ederler. Enzimlerin aktif merkezinde bulunarak enzimatik katalizde rol alırlar. Aspartik asit Glutamik

53
Asidik yan grupların amidleri
R gruplarında ilave bir amid yapısı bulunur. Yüksüz ama polar özellik gösterirler Asparajin proteinlere karbohidrat eklenme noktasıdır. Asparajin Glutamin

54
Aspartik asit Asp D 2.0 9.9 3.9 Asparajin Asn N 2.1 8.8 Glutamik asit Glu E 9.5 4.1 Glutamin Gln Q 2.2 9.1

55
Bazik amino asitler-1 pH 7’de net pozitif yüke sahip bu amino asitler asidik aa’lerle iyonik bağ oluşumu ve proteinlerin 3 boyutlu yapısının stabilizasyonunda yer alırlar. Bazı enzimlerin aktif merkezinde katalitik rol üstlenirler. Arginin Lizin Histidin

56
Arginin Arg R 1.8 9.0 12.5 Lizin Lys K 2.2 9.2 10.8 Histidin His H 6.0

57
Bazik amino asitler-2 Arginin guanido grubu
Lizin pozisyonunda bir amino grubu ve histidin imidazol halkası içerir. Histidin R grubu (imidazol halkası) pKa’sı fizyolojik pH’a yakın (6.0) olduğu için fizyolojik pH’ta tamponlama yapabilen tek aa histidindir. Proteinlerin tamponlama yapabilme özellikleri histidin içerikleri ile ilişkilidir.
 pKa’sı fizyolojik pH’a yakın (6.0) olduğu için fizyolojik pH’ta tamponlama yapabilen tek aa histidindir. Proteinlerin tamponlama yapabilme özellikleri histidin içerikleri ile ilişkilidir.")
58
Aromatik amino asitler-1
Yan zincirlerinde aromatik halka içerirler Tirozin (aynı zamanda hidroksilli) Histidin (aynı zamanda bazik) Fenil alanin (benzen halkası) Triptofan (indol halkası)
 Histidin (aynı zamanda bazik) Fenil alanin (benzen halkası) Triptofan (indol halkası)")
59
Aromatik amino asitler-2
Fenilalanin, triptofan ve tirozin non-polar (hidrofobik) yapıdadır ve genellikle proteinlerin iç kısmında yer alırlar. Tirozin –OH grubu ile H bağı oluşturabilir.
 yapıdadır ve genellikle proteinlerin iç kısmında yer alırlar. Tirozin –OH grubu ile H bağı oluşturabilir.")
60
Fenil-alanin Phe F 2.2 9.2 Triptofan Trp W 2.4 9.4

61
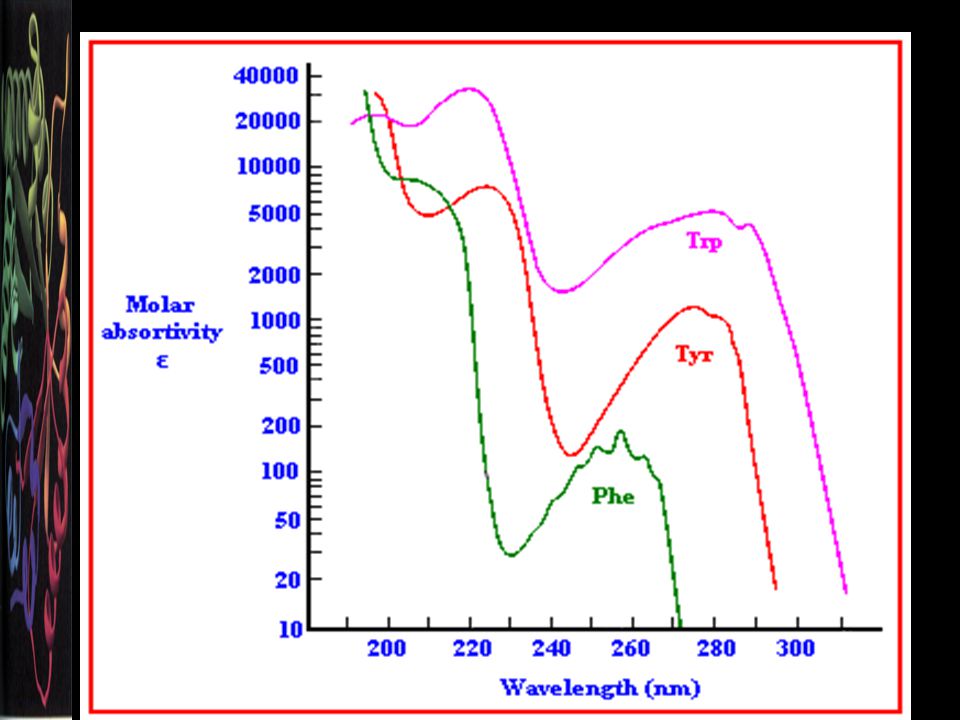
Aromatik amino asitler-3
Aa’ler renksizdir. 240 nm üzeri ultraviole ve görünür ışığı absorbe etmezler. Aksine en çok triptofan olmak üzere, tirozin ve az oranda fenil alanin 280 nm’de ışığı absorbe ederler. Çözeltilerde protein varlığını göstermek için bu özellikten yararlanılır.

63
İmino asit Halkalı yapısı ve amino yerine imino grubu (sekonder amin) olan prolin bu grupta yer alır. Prolin halkası rijit (bükülmez) özelliktedir. Kemik tendon gibi yapıların proteini olan kollojende bol miktarda bulunur. Ayrıca kollojen yapısında protein sentezlendikten sonra bazı prolinler enzimatik olarak hidroksillenir: hidroksiprolin de bir imino asittir.
 özelliktedir. Kemik tendon gibi yapıların proteini olan kollojende bol miktarda bulunur. Ayrıca kollojen yapısında protein sentezlendikten sonra bazı prolinler enzimatik olarak hidroksillenir: hidroksiprolin de bir imino asittir.")
64
Prolin Pro P 2.0 10.6

65
Amino asitlerin polaritelerine göre sınıflaması
Biyolojik ortamda davranışları hakkında fikir verir. Polarite polar grup varlığı ve hidrokarbon yapının büyüklüğü ile ilgilidir. Non-polar (hidrofobik) Polar ve yüksüz Polar asidik Polar bazik
 Polar ve yüksüz. Polar asidik. Polar bazik.")
66
Polar Non-Polar Hidrofobik Triptofan Fenilalanin Izolösin Tirozin Lösin Valin Metiyonin Yüklü Arginin (+) Glutamik a (-) Aspartik a (-) Lizin (+) Histidin (+) Yüksüz Sistein Prolin Serin Glutamin Asparagin Ortada Glisin Treonin Alanin
 Glutamik a (-) Aspartik a (-) Lizin (+) Histidin (+) Yüksüz. Sistein. Prolin. Serin. Glutamin. Asparagin. Ortada. Glisin. Treonin. Alanin.")
67
Hidrofobik İndeks Arginin -11.2 Glisin 0 Glutamik A -9.9 Threonin 0.4
Aspartik A -7.4 Lizin -4.2 Histidin -3.3 Sistein -2.8 Prolin -0.5 Serin -0.3 Glutamin -0.3 Asparagin -0.2 Glisin 0 Threonin 0.4 Alanin 0.5 Metiyonin 1.3 Valin 1.5 Lösin 1.8 Tirozin 2.3 Izolösin 2.5 Fenilalanin 2.5 Triptofan 3.4

68
21. Amino Asit

69
NON-STANDART AA’LER Genetik kodları yok enzimatik olarak sentezlenirler. 1) Protein yapısında bulunmayan, metabolik yollarda ara ürün olarak oluşan aa’ler Ör: ornitin, sitrüllin (üre sentezi) 2) Farklı işlevleri olan aa’ler Ör: Tiroid hormonları, kreatin (kas dokusunda) 3) Protein çatısının oluşmasından sonra değişikliğe uğrayan aaler Ör: 4-OH prolin (kollojen yapısında)
 Protein yapısında bulunmayan, metabolik yollarda ara ürün olarak oluşan aa’ler. Ör: ornitin, sitrüllin (üre sentezi) 2) Farklı işlevleri olan aa’ler. Ör: Tiroid hormonları, kreatin (kas dokusunda) 3) Protein çatısının oluşmasından sonra değişikliğe uğrayan aaler. Ör: 4-OH prolin (kollojen yapısında)")
70
PROTEİNLERİN SENTEZ SONRASI DÜZENLENMELERİ
1) Protein katlanması 2) Proteolitik işlenme 3) Yan grupların eklenmesi 4) Aminoasitlerin sentez sonrası değişimleri (Post-translasyonel modifikasyonlar) Gama karboksilasyon Metilasyon Hidroksilasyon Fosforilasyon Asetilasyon Adenilasyon Üridilasyon ADP ribozilasyon -Ubikuinasyon
 Protein katlanması. 2) Proteolitik işlenme. 3) Yan grupların eklenmesi. 4) Aminoasitlerin sentez sonrası değişimleri. (Post-translasyonel modifikasyonlar) Gama karboksilasyon. Metilasyon. Hidroksilasyon. Fosforilasyon. Asetilasyon. Adenilasyon. Üridilasyon. ADP ribozilasyon. -Ubikuinasyon.")
71
g-karboksiglutamat- Ca++ bağlar
Aspartat ve glutamat kalıntılarına karboksil grubu eklenebilir. Bazı pıhtılaşma proteinlerinde (Faktör 2, 7, 9 ve 10, protein S ve C) Ve kemik proteini osteokalsinde bulunur. Güçlü kalsiyum bağlar. Karboksilasyon için K vitamini gereklidir.
 Ve kemik proteini osteokalsinde. bulunur. Güçlü kalsiyum. bağlar. Karboksilasyon. için K vitamini gereklidir.")
72
Metilasyon Hidroksilasyon
Lizin, histidin, glutamat gibi aaler metillenir. Kas proteinleri Hidroksilasyon OH Prolin ve OH lizin Kollajen OHlizin ve kh yan gruplar

73
FOSFORİLASYON-DEFOSFORİLASYON
Serin, tirozin ve treonin kalıntılarının OH gruplarına ATP’den fosfat grubu aktarılır. Proteinlerin etkinleşmesi ve Enzim aktivitesinin düzenlenmesinde yer alır.

74
Glikojen, laktoz sentezi
Bakteri toksinleri, G proteinleri Bakteri toksinleri, G proteinleri

75
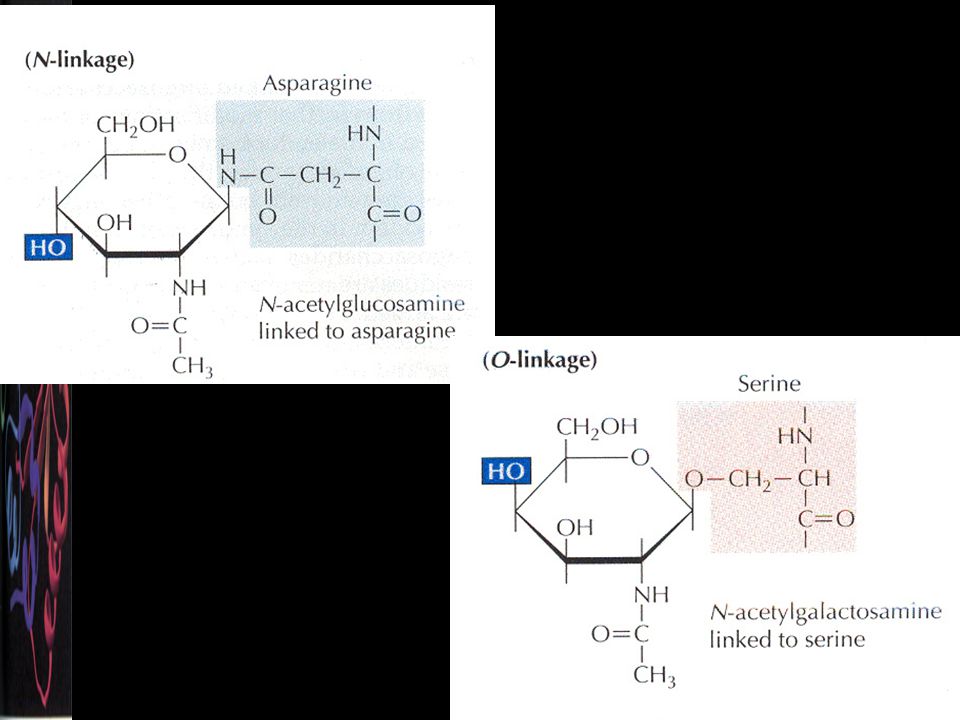
Karbohidrat yan grupların eklenmesi
Proteinlerin post-translasyonel değişim işlemlerinden biri. Karbohidrat yan gruplar sentez sırasında veya sonra kovalent olarak enzimatik yolla eklenir. İki tip glikozilasyon tanımlanmıştır: Asparajin, N-glikozilasyon (ER) Serin, treonin veya OH-lizin, O-glikozilasyon (Golgi) Sitoplazmik proteinler nadiren glikozillenirler. Salgı proteinleri, lizozomal enzimler, çoğu hücre yüzey proteinleri
 Serin, treonin veya OH-lizin, O-glikozilasyon (Golgi) Sitoplazmik proteinler nadiren glikozillenirler. Salgı proteinleri, lizozomal enzimler, çoğu hücre yüzey proteinleri.")
77
Amino asit tepkimeleri
Karboksil grubu tepkimeleri; esterifikasyon, tuz oluşumu, açilasyon Amino grubu tepkimeleri: bazıları aa’lerin varlığının gösterilmesi, miktarının saptanması ve proteinlerin aa dizisinin belirlenmesinde kullanılır. R grubu tepkimeleri: her aa için farklıdır, o aa’in nicel ve nitel analizi için kullanılır

78
Amino grubu tepkimeleri-1
Ninhidrin reaksiyonu: alfa aa’lerin ninhidrin ile tepkimesi sonucu mavi-mor renkli ürün oluşur. Prolin ve OH-Prolin ninhidrin ile sarı kompleks oluşturur. Aa’ler dışındaki aminler ve amonyak da ninhidrin ile reaksiyona girer ancak CO2 oluşmaz. Ürün başlangıç aa’e göre değişmez; tüm aa’ler aynı ürünü verirler.

79
O O O H R H * O O H N H H O O O O C O 2 H 2 O N R * H O O

80
Amino grubu tepkimeleri-2
Diğer amino grubu tepkimelerinde ise ürün aa’in R grubunu taşıdığı için aa’in tanınmasına olanak tanır: Sanger reaktifi (1-floro,2-4 dinitrobenzen) Hassas yöntemler, ürünleri zor koşullarda stabil ama peptid dizi analizine uygun değil Edman reak : protein ve peptid dizi analizine uygundur.
 Hassas yöntemler, ürünleri zor koşullarda stabil ama peptid dizi analizine uygun değil. Edman reak : protein ve peptid dizi analizine uygundur.")
81
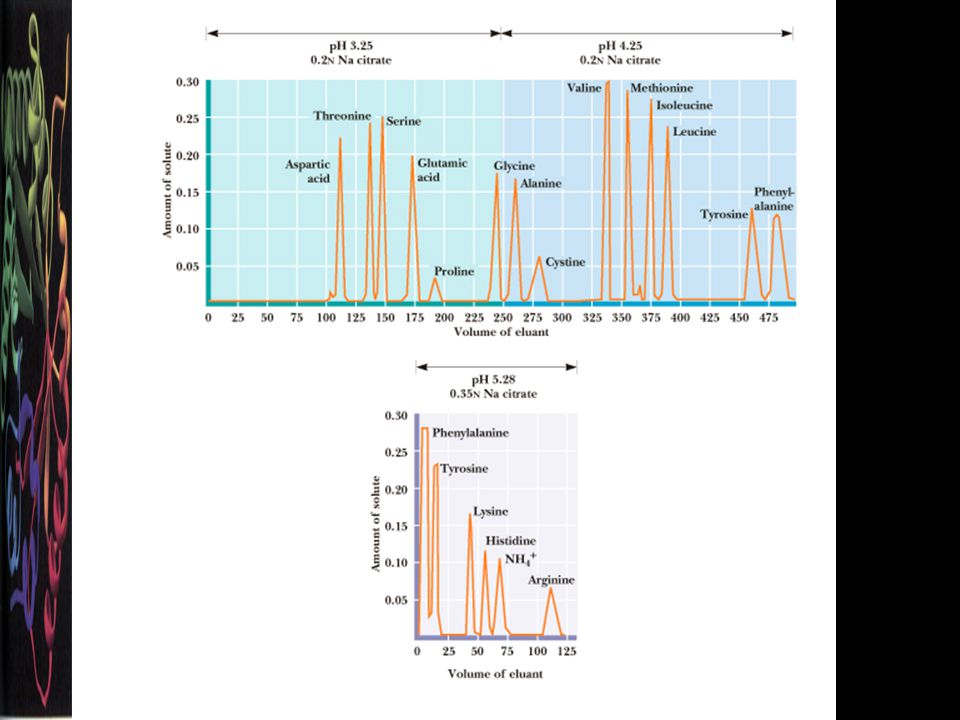
Amino asitlerin ayırım teknikleri
A) Polarite farklılıklarına göre ayırım Kağıt kromatografisi İnce tabaka kromatografisi B) Belirli bir pH’taki yük farklılıklarına göre ayırım İyon değiştirici kromatografi Yüksek voltajlı elektroforez HPLC: Gelişmiş kromatografi cihazı C) Kütleye göre ayırım Kütle spektrometresi; PTMlar da saptanabilir.
 Polarite farklılıklarına göre ayırım. Kağıt kromatografisi. İnce tabaka kromatografisi. B) Belirli bir pH’taki yük farklılıklarına göre ayırım. İyon değiştirici kromatografi. Yüksek voltajlı elektroforez. HPLC: Gelişmiş kromatografi cihazı. C) Kütleye göre ayırım. Kütle spektrometresi; PTMlar da saptanabilir.")
82
İyon Değiştirici Kromatografi

85
elektroforez ÖR: pH 5.5

86
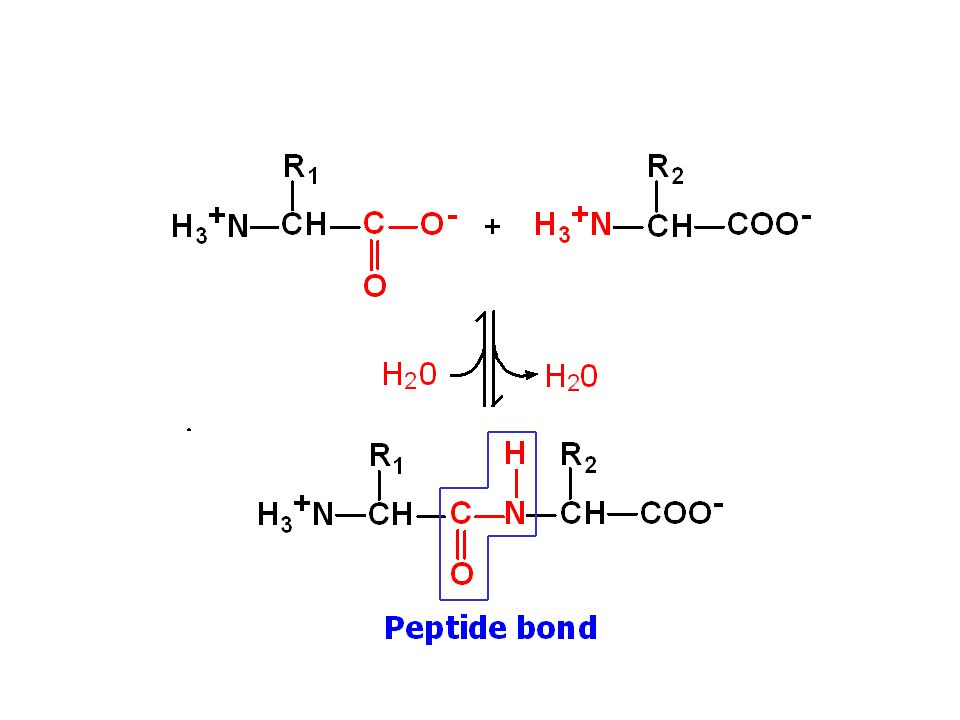
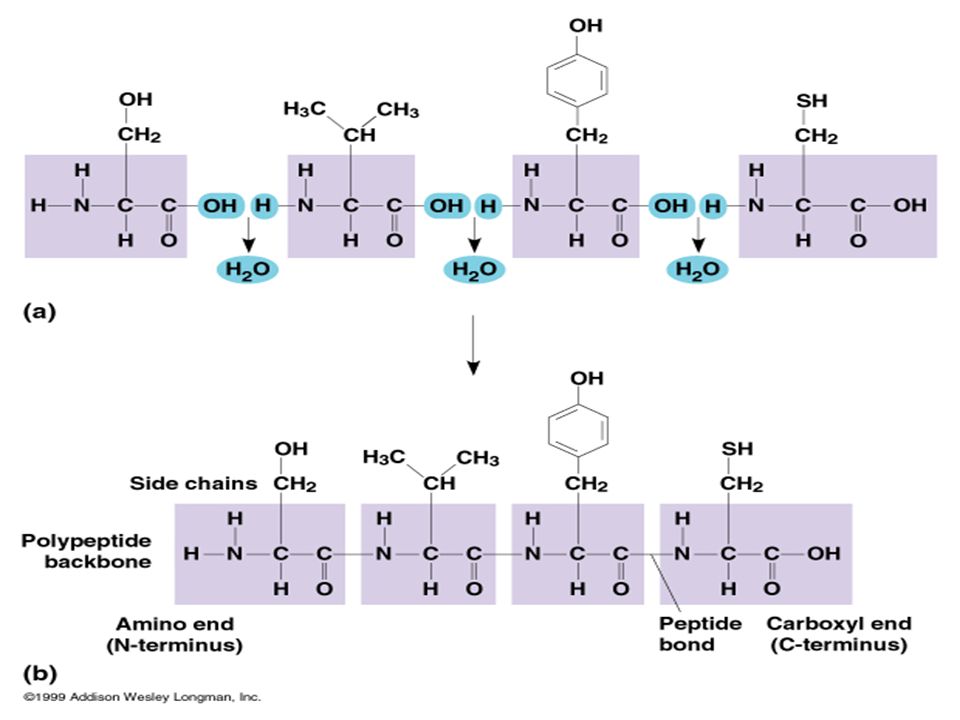
Amino asitlerin en önemli tepkimeleri peptit bağı oluşumudur
Bir aa’in amino grubu ile diğer bir aa’in karboksil grupları arasında bir molekül su açığa çıkışı ile oluşan kovalent bağa peptit bağı denilir.

88
Peptid bağı oluşumu: Peptid bağı aslında bir AMİD BAĞIdır
kondansasyon

89
Peptid bağı oluşumu Peptid bağı oluşumu bir kondansasyon tepkimesidir (tersi hidroliz). Fizyolojik pH’ta bu reaksiyon kendiliğinden meydana gelmez çünkü endergoniktir DG=21 kjul/mol’dür. Peptidler ribozomlarda depo enerji karşılığında sentezlenir. Peptid bağlarının hidrolizi ise egzergonik bir reaksiyon olmasına rağmen aktivasyon enerjisi yüksek olduğu için peptid ve proteinler oldukça stabildirler.

90
Peptid bağının özellikleri
Peptid bağları fizyolojik pH’ta polar ama yüksüzdür. Kısmi çift bağ özelliğindedir Düzlemseldir Bükülmez Transtır

91
1-Peptid bağının elektrik yükü
Peptid bağı oluşumu ile bir (-) ve bir (+) yük kaybedilmiş olur. Bu nedenle peptid bağı yüksüzdür ama elektron paylaşımı eş olmadığı için polardır. Peptidler ise iki ucundaki serbest amino , karboksil ve R gruplarındaki yükler nedeni ile yüklü moleküllerdir: polielektrolitlerdir.
 ve bir (+) yük kaybedilmiş olur. Bu nedenle peptid bağı yüksüzdür ama elektron paylaşımı eş olmadığı için polardır. Peptidler ise iki ucundaki serbest amino , karboksil ve R gruplarındaki yükler nedeni ile yüklü moleküllerdir: polielektrolitlerdir.")
92
2-Peptid bağları kısmi çift bağ karakterindedir
Hem oksijen hem de azot atomları elektronegatif atomlardır. Ancak oksijen azottan biraz daha elektronegatiftir. Bu nedenle çift bağ (yani paylaşılan 2. elektron) C ile O arasında gösterilir. Oysa gerçekte bu elektronun paylaşımı ve çift bağın yeri rezonans göstermektedir. Buna kısmi çift bağ denilir
 C ile O arasında gösterilir. Oysa gerçekte bu elektronun paylaşımı ve çift bağın yeri rezonans göstermektedir. Buna kısmi çift bağ denilir.")
93
Peptid bağında rezonans
Peptide bond resonance

94
3-Peptid bağı düzlemsel ve bükülmezdir
Kısmi çift bağ normal bir çift bağdan daha kısadır. Bu peptid bağının rijit (bükülmez) ve planar (düzlemsel) olmasına yol açar. C ve N atomlarını birleştiren peptid bağında rotasyon olmadığı için bu 4 atom aynı düzlemde bulunur.
 ve planar (düzlemsel) olmasına yol açar. C ve N atomlarını birleştiren peptid bağında rotasyon olmadığı için bu 4 atom aynı düzlemde bulunur.")
95
4- Peptid bağı transtır Prolinler %10’u sistir sis trans
C1 N2 1 C2 N1 oxygen trans Tercih edilendir sis

96
PEPTİDLER Peptidler aa oligo- veya poli-merleridir.
Bir kaç aa –binlerce aa İki aa: dipeptid Birkaç aa: oligopeptid Çok sayıda aa: polipeptid Karboksil grubu peptid bağı yapısına giren bir aa’e amino asit kalıntısı denilir ve adının sonuna -il eki getirilir. Adlandırma amino ucundan başlar Bu kurala göre peptid bağına karboksil grubu ile aspartatın, amino grubu ile alaninin katıldığı bir dipeptid aspartil-alanin dipeptididir.

97
Aspartil-alanin dipeptidi
Karboksil ucu Amino ucu

99
Peptidler Biyolojik aktivitesi olan peptidler: bazı hormon ve nörotransmitterler peptid yapısındadır Sentetik olarak üretilen peptidler: antibiyotik, tatlandırıcı Tek zincirden oluşur ve MA genellikle Da altındadır. Proteinlerin çatısını oluşturan peptidler.

100
Proteinlerin işlevleri
Taşıyıcı: vitamin, hormon, besin, oksijen Katalitik: enzimler Besin ve depo: ferritin (Fe), ovalbumin Kontraktil, motil: aktin, miyozin Yapısal: kollojen, elastin, keratin, fibroin Savunma: Immunoglobulinler Düzenleyici: hormonlar Çeşitli ve yaşam için zorunlu
, ovalbumin. Kontraktil, motil: aktin, miyozin. Yapısal: kollojen, elastin, keratin, fibroin. Savunma: Immunoglobulinler. Düzenleyici: hormonlar. Çeşitli ve yaşam için zorunlu.")
101
Proteinler büyük moleküllerdir
Tek veya fazla polipeptid zinciri Bir polipeptidden oluşan her bir kısma subünite veya protomer denilir. Subüniteler eş ise homodi-, homotetra- merik protein denilir. Subüniteler eş olmadığında hetero- öntakısı ile söylenir. Aa sayısı aa arasında değişir. Aa sayısı MA/110

104
Proteinler aa dışı yapılar da bulundurabilir
Bazı proteinler aa dışında kimyasal gruplar taşır: konjuge, kompleks veya birleşik proteinler denilir. Aa dışındaki kısma prostetik grup adı verilir. Prostetik gruplar protein işlevi ile ilgilidirler. Sadece aa’ten oluşan proteinler basit proteinler olarak adlandırılır.

105
Sınıf Prostetik grup Örnek Lipoprotein lipid HDL Glikoprotein karbohidrat İmmunglobulin G Hemoprotein Hem grubu hemoglobin Flavoprotein Flavin nükleotid Süksinat dehidrojenaz Metalloprotein demir ferritin

106
Şekilllerine göre sınıflama
1) Globüler proteinler: peptid zincirinin kıvrılıp katlanması ile kompakt bir yapı oluştururlar. Uzunluk/genişlik oranı (aksiyal oran) 10’dan küçüktür. Suda çözünür. Çoğu plazma proteini globülerdir 2) Fibröz proteinler: katlanmalar minimum, aksiyal oran 10’un üzerinde Daha çok yapısal proteinlerdir. İçerdikleri aa’lerin çeşitliliği düşüktür Düzenli bir sekonder yapıları vardır. Suda çözünmez.
 Globüler proteinler: peptid zincirinin kıvrılıp katlanması ile kompakt bir yapı oluştururlar. Uzunluk/genişlik oranı (aksiyal oran) 10’dan küçüktür. Suda çözünür. Çoğu plazma proteini globülerdir. 2) Fibröz proteinler: katlanmalar minimum, aksiyal oran 10’un üzerinde. Daha çok yapısal proteinlerdir. İçerdikleri aa’lerin çeşitliliği düşüktür. Düzenli bir sekonder yapıları vardır. Suda çözünmez.")
107
Globüler protein Fibröz protein

108
Bir proteini oluşturan aa’lerin çok sayıda
konformasyonu mümkündür. Ancak sulu çözeltilerde belirli konformasyonların oluştuğu görülmüştür. Peptid ve proteinlerin özgün konformasyonları fizyolojik işlevleri için zorunludur.

109
Konformasyon nedir? Bir molekülü (proteini) oluşturan aa dizisi değişmeden, tüm atomların diğer tüm atomlarla ilişkisinin belirlediği 3 boyutlu yapı. Non-kovalent güçlerin kırılıp yeniden yapılması ile değişebilir. Konfigürasyon: Bir grup atom arasındaki geometrik ilişki. Konfigürasyon değişimi kovalent bağların kırılıp yeniden yapılması ile mümkündür. Örneğin D ve L konfigürasyonlar.
 oluşturan aa dizisi değişmeden, tüm atomların diğer tüm atomlarla ilişkisinin belirlediği 3 boyutlu yapı. Non-kovalent güçlerin kırılıp yeniden yapılması ile değişebilir. Konfigürasyon: Bir grup atom arasındaki geometrik ilişki. Konfigürasyon değişimi kovalent bağların kırılıp yeniden yapılması ile mümkündür. Örneğin D ve L konfigürasyonlar.")
110
SORU: Proteinler neden katlanır?
BİYOKİMYASAL REAKSİYONLAR TERMODİNAMİK YASALARA UYGUN OLARAK GERÇEKLEŞİR

111
Protein konformasyonlarını stabilize eden güçler
Bir sistem en az serbest enerji (DG) gerektiren şekilde bulunma eğilimindedir. Bunu ısının azalması (entalpi DH) ve düzensizliğin artması (entropi DS) destekler. DG= DH-T DS Bu iki koşul sağlandığında ‘olay’ (burada protein konformasyonu) kendiliğinden gerçekleşir.
 gerektiren şekilde bulunma eğilimindedir. Bunu ısının azalması (entalpi DH) ve düzensizliğin artması (entropi DS) destekler. DG= DH-T DS. Bu iki koşul sağlandığında ‘olay’ (burada protein konformasyonu) kendiliğinden gerçekleşir.")
112
GENEL KURAL iki atom arasında bağ oluştuğunda ısı şeklinde bir miktar enerji kaybedilir. Bu serbest enerji açısından istenilen bir durumdur. Bağların artması yapıları stabilize eder (- entalpi, istenir). iki atom arasında bağ oluştuğunda düzensizlik azalır (- entropi, istenmez).
. iki atom arasında bağ oluştuğunda düzensizlik azalır (- entropi, istenmez).")
114
Sulu ortamda protein molekülün davranışına etkili 4 faktör:
Suyun ısısı, suyun düzensizliği, protein molekülünün ısısı ve protein molekülünün düzensizliğidir.

115
Proteinin bağ oluşumu ile katlanması…
protein molekülünün düzensizliği azalır (- entropi, istenmez). protein molekülünün ısısı azalır (- entalpi, istenir).
. protein molekülünün ısısı azalır (- entalpi, istenir).")
116
su ile hidrofobik yapıların bir araya gelmesi suyun düzensizliğini azaltır.
Protein katlanması su ile hidrofobik yapıların temasını azaltır yani suyun düzensizliği artar (+ entropi, istenir). Sonuç 1: Protein ve su kendileri ve birbirleri ile maksimum sayıda H bağı oluşturmalıdır (- entalpi) Sonuç 2: hidrofobik aaler proteinin iç kısmına gömülmelidir (+ entropi)
. Sonuç 1: Protein ve su kendileri ve birbirleri ile maksimum sayıda H bağı oluşturmalıdır (- entalpi) Sonuç 2: hidrofobik aaler proteinin iç kısmına gömülmelidir (+ entropi)")
118
Protein konformasyonlarını stabilize eden güçler
Hidrojen bağları Hidrofobik etkileşimler Elektrostatik etkileşimler Van der waals etkileşimleri

119
Protein yapısının hiyerarsik incelenmesi

120
Primer yapı Amino asit dizisi ve disülfit bağlarının yeri bir proteinin primer yapısını oluşturur. Genetik olarak belirlenir. Peptid bağları ve S-S bağları ile stabilizedir. Tümü kovalent bağlardır.

121
Sekonder yapı Birincil yapıyı oluşturan aa’ler arasında zincir içi H bağları aracılığı ile sağlanan lokal etkileşimlerle oluşan düzenli yapılar. Sekonder yapılar protein işlevi için önem taşır. Fibröz proteinlerin sekonder yapıları işlevleri için özelleşmiştir.

122
Sekonder yapılar Alfa heliks Beta pileli düzlemsel yapı Beta dönüş
İlmik (halka) Kollajen heliks Düzensiz yapılar
 Kollajen heliks. Düzensiz yapılar.")
123
-Helix Polipeptid yapısındaki her C atomu eş miktarda kıvrılırsa -heliks meydana gelir. Bu kıvrılma her peptid bağını oluşturan N’un 4 önceki peptid bağını oluşturan karbonil O’i ile oluşturduğu H bağı ile sağlanır. Bir tam dönüş 3.6aa içerir ve 0.54 nm yüksekliğindedir. Maksimum sayıda H bağı nedeni ile en düşük enerjili konformasyondur, kendiliğinden meydana gelir. Diğer non-kovalent bağlarla da desteklenir.

124
zincir-içi hidrojen bağları

125
-Heliks Globüler proteinlerin ¼’ü a heliks oluşturur.
a keratin başlıca a heliks yapısındadır. Ala sık Gly destabilize eder Pro bulunmaz

126
Sekonder yapılar 2: Beta pileli düzlemsel yapı
a-Helikse benzer yönleri: Maksimum sayıda H bağı ile oluşur. H bağları yine farklı 2 peptid bağını oluşturan N ve karbonil O’i arasında meydana gelir. a-Heliksten farklı yönleri: H bağları polipeptid eksenine paralel değildir. H bağlarını oluşturan aa’lerin primer yapıdaki konumları birbirine uzak olabilir Katlanma az

127
-düzlemsel yapı R grupları polipeptid çatısının bir üstünde bir altında olduğu için pileli görünüm verir. Alfa heliksten farklı olarak katlanmanın az olduğu düzlemsel bir yapıdır.

128
-heliks b-düzlemsel yapı
-pleated sheets

129
Fibröz proteinlerin sekonder yapıları görevleri için özelleşmiştir
Saç ve tırnakta bulunan a-keratin a-heliks yapısındadır. Kemik ve tendonda bulunan kollajen 3’lü heliks gerilime dayanıklılığı sağlar İpekte bulunan b-fibroin b-pilili düzlemsel yapısı ile yumuşak, bükülebilir, ancak gerilmeye dayanıklıdır.

130
Tersiyer yapılar Katlanma, bükülme vb. ile primer yapıda birbirinden uzak konumda bulunabilen aa’lerin etkileşimi ile oluşan 3 boyutlu yapı. Non-kovalent etkileşimler; H bağları, elektrostatik etkileşimler, hidrofobik etkileşimler, van der waals etkileşimleri Ve S-S bağları ile stabilize olur

131
Tersiyer ve kuarterner yapıları
stabilize eden güçler

132
Süpersekonder yapılar (motifler)
Çoğu globüler proteinde belirli sekonder yapıların bir düzen içinde bir araya gelmesi ile motifler oluşur. Üçüncül (tersiyer) yapı elemanları olarak kabul edilirler.
 yapı elemanları olarak kabul edilirler.")
133
Motifler Heliks-ilmik-heliks 4’lü heliks demeti
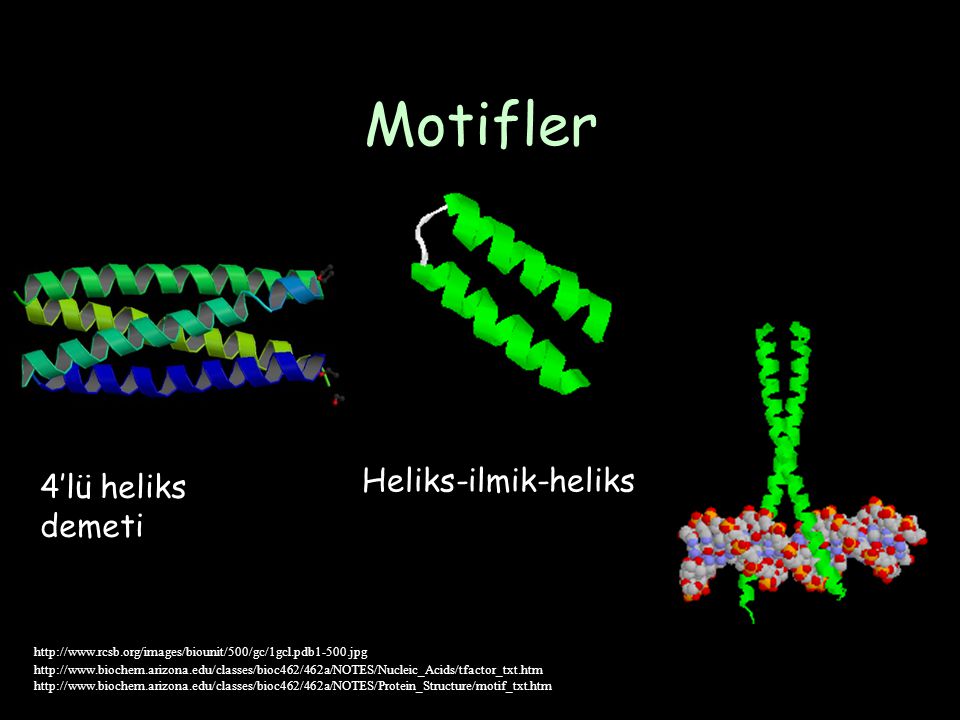
134
Kısımlar (domain) Genellikle ligand bağlamak gibi özel işlevler üstlenen yapısal birimlerin oluşturduğu 3 boyutlu yapı
 Genellikle ligand bağlamak gibi özel işlevler üstlenen yapısal birimlerin oluşturduğu 3 boyutlu yapı.")
135
Domainler alfa/beta barrel beta barrel
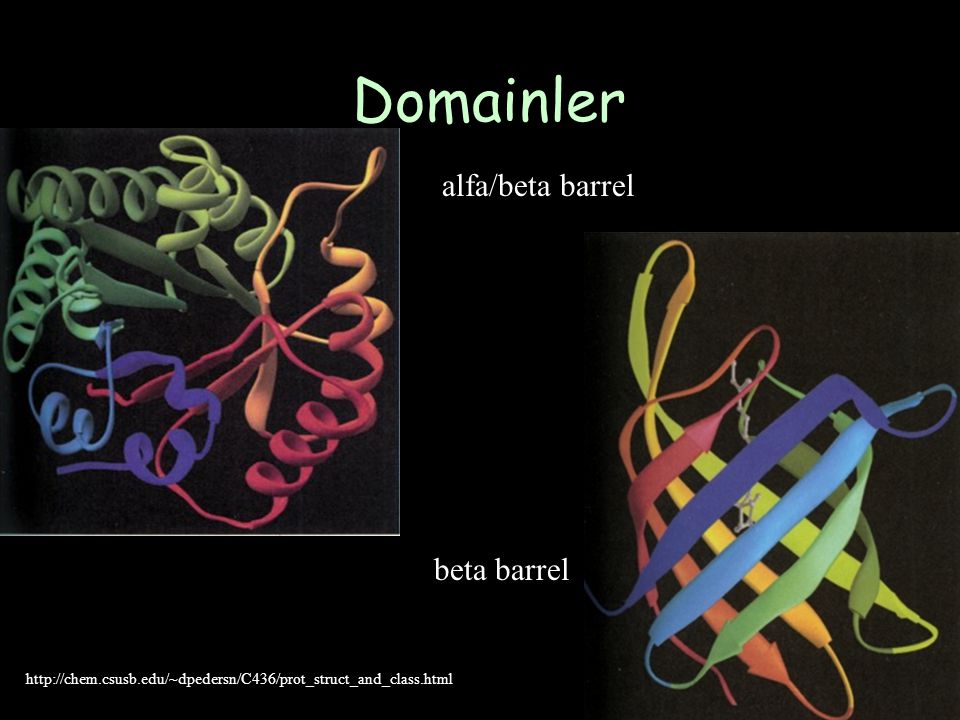
136
Kuarterner yapı Birden fazla polipeptid zincirinden oluşan proteinler için söz konusudur. Subünitelerin birbiri ile ilişkisini kapsar. Tersiyer yapıda olduğu gibi non-kovalent etkileşimler ve S-S bağları ile stabilize olur.

137
4o kuarterner yapı Monomerik protein (kuarterner yapı yok)
Dimerik protein (homodimer) Dimerik protein (heterodimer) heterotetramer heteropolimerik protein
 Dimerik protein (heterodimer) heterotetramer. heteropolimerik protein.")
139
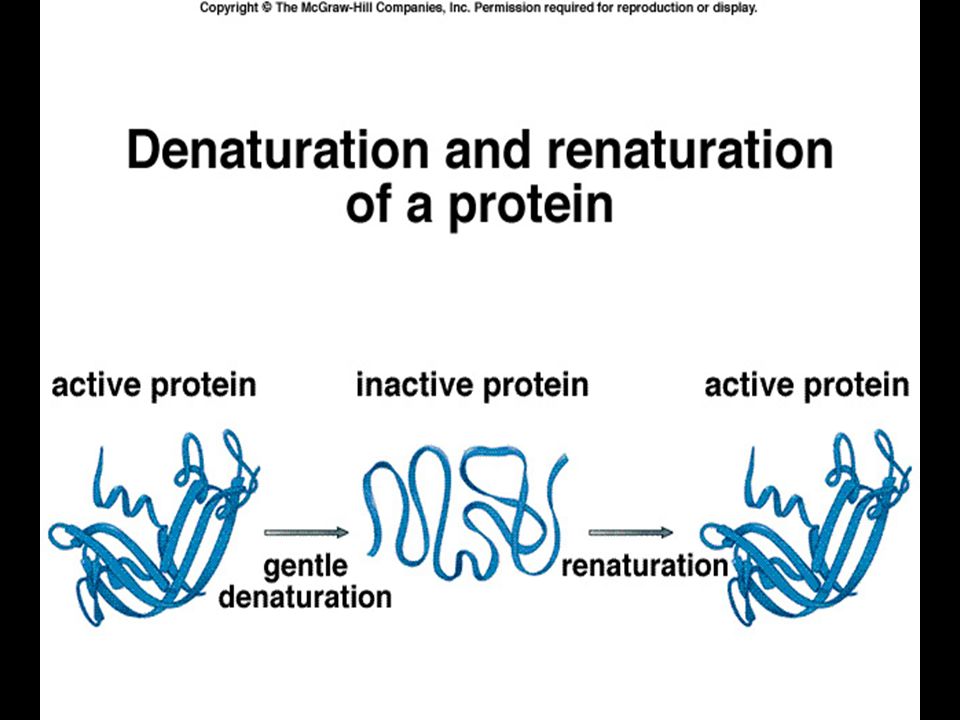
Denatürasyon Kimyasal, fiziksel, vb. nedenlerle proteinlerin sekonder, tersiyer ve kuarterner yapılarının bozulmasıdır. Yani primer aa dizisi korunmuştur (peptid bağları) ama konformasyon (3 boyutlu özgün yapıyı sağlayan non-kovalent etkileşimler) bozulmuştur. Konformasyon değişikliği biyolojik aktiviteyi engeller.
 ama konformasyon (3 boyutlu özgün yapıyı sağlayan non-kovalent etkileşimler) bozulmuştur. Konformasyon değişikliği biyolojik aktiviteyi engeller.")
140
Denatürasyona yol açan nedenler
Sıcaklık, asitler, bazlar, üre, Sodyum dödesil sülfat (SDS), guanidin, X-ışınları gibi non*kovalent etkileşimleri bozan maddelere kaotropik maddeler denilir. Bazı maddelerle oluşan denatürasyon geri dönüşümlüdür; renatürasyon denilir.
, guanidin, X-ışınları gibi non*kovalent etkileşimleri bozan maddelere kaotropik maddeler denilir. Bazı maddelerle oluşan denatürasyon geri dönüşümlüdür; renatürasyon denilir.")
142
Proteinlerin katlanması
Denatürasyona yol açan faktörlerin ortadan kaldırılması ile renatürasyonun kendiliğinden meydana gelmesi ve hatta bunun saniyeler içinde gerçekleşmesi dikkat çekmiştir. Proteinlerin 3 boyutlu yapısını belirleyen faktörün primer yapısı yani aa dizisi olduğu kararı verilmiştir: Buna Anfisen teorisi denilir

143
Sekonder yapının belirlenmesi
3 boyutlu yapı belirlenmesi Anfinsen, CB (1973) Principles that govern the folding of protein chains. Science 181,
 Principles that govern the folding of protein chains. Science 181,")
144
Ör: Orak hücreli anemi Orak hücreli anemi protein primer yapısında tek aa’lik değişim ile ilişkilidir. Hemoglobini oluşturan beta polipeptidinde Glutamik asit yerine valin bulunması işlevi ve yapıyı etkiler

145
Orak hücreli anemi

146
Proteinler doğru katlanma şeklini nasıl bulabilmektedir?
Proteinler enerjetik olarak katlanma eğilimindedir. Ama nasıl katlanacağını nasıl bilmektedir? Aslında proteinlerin katlanma konusunda fazla seçeneği yoktur. Çünkü...

147
(1) Peptid bağı düzlemseldir: bu düzlemde yer alan 4 atom ve 3 bağ için açılar sabittir.
 Peptid bağı düzlemseldir: bu düzlemde yer alan 4 atom ve 3 bağ için açılar sabittir.")
148
(2) R gruplarının yer kaplama etkisinden kaçınmak için transtır.
potential for steric hindrance

149
(3) zaten peptid ekseni üzerinde hareketli
olabilecek sadece 2 bağ vardır. a-C ile karbonil C’u arasındaki Psi (ψ) açısı a-C ile amino N’u arasındaki Phi (Φ) açısı
 açısı. a-C ile amino N’u arasındaki Phi (Φ) açısı.")
150
(4) Phi ve psi açıları ise aa yan gruplarının yer kaplama etkisi, van der waals itme mesafesi gibi nedenlerle tüm değerleri alamaz. Örneğin phi açısının belirli bir değerinde psi açısının alabileceği değerler sınırlıdır.

151
Ramachandran grafiği Ramachandran phi’ye karşı psi’nin
alabileceği değerleri grafik üzerinde göstermiştir.

152
Bu değerlerin belirli sekonder yapıları sağladığı anlaşılmıştır.

153
Katlanmanın Enerjisi Aslında katlanmış ve katlanmamış (denatüre) durumlar arasındaki enerji farkı çok küçüktür, DG = kcal/mol. DG’nin küçük olması protein konformasyonun çok stabil olmamasını sağlar ve proteine yapısal esneklik kazandırır. Çünkü genellikle proteinlerin işlevi için büyük konformasyonel esneklik gereklidir. Böylece işi biten veya eskiyen proteinlerin kolayca yıkımı da mümkün olur.

154
Proteinlerin esnek üç boyutlu yapısına
örnek olarak enzim

155
Geçiş formlarının enerjisi; doğru katlanma en düşük enerjili yani en sağlam protein konformasyonunu sağlar.

156
Levinthal paradoksu in vitro in vivo Denatüre protein 106 olası
konformasyon katlanma katlanma 1 stabil konformasyon t = saniyeler t =

Benzer bir sunumlar








 ile karakterize edilir. Verilen bir elementin tüm atomlarında.>")










>")






