Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Introduction to Genetic Analysis
Anthony Griffiths • Susan Wessler • Richard Lewontin William Gelbart • David Suzuki • Jeffrey Miller Introduction to Genetic Analysis Eighth Edition Chapter 7: DNA: Structure and Replication Copyright © 2005 by W. H. Freeman & Company

2
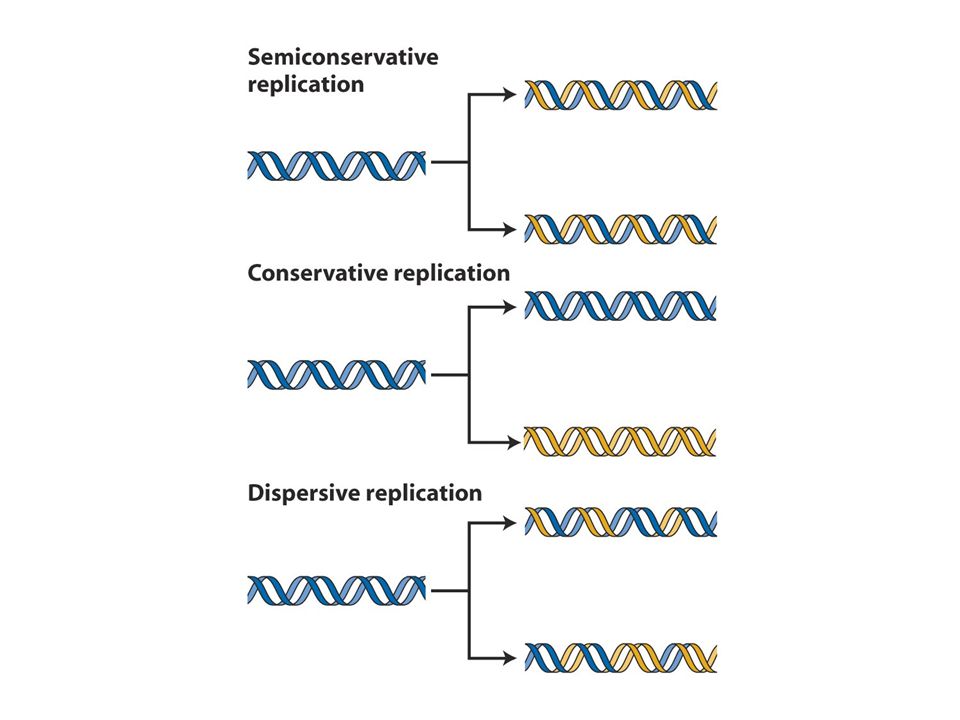
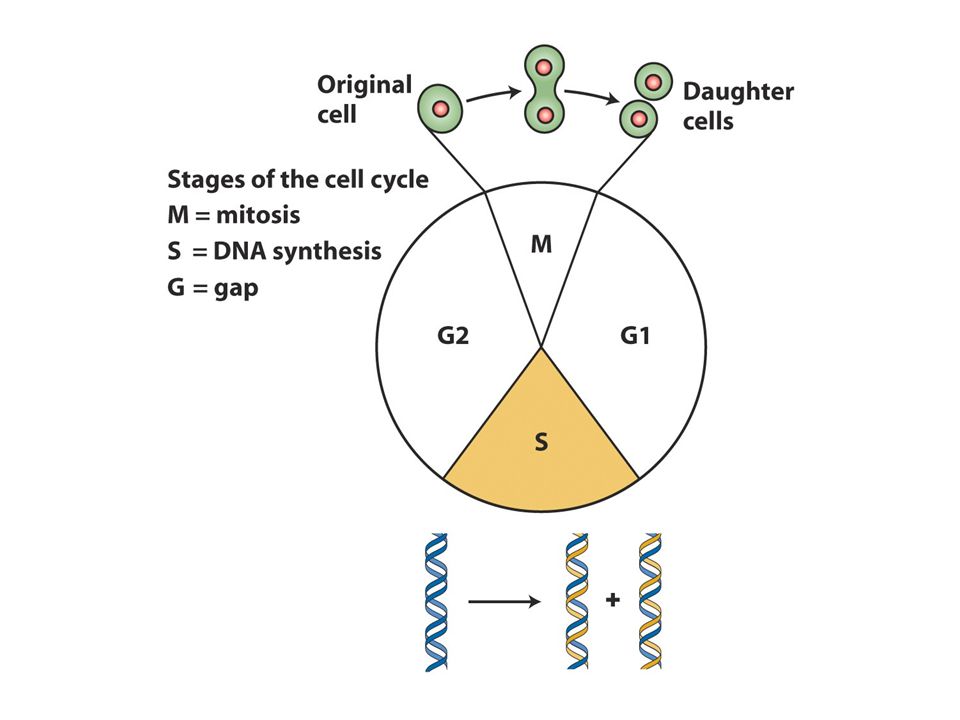
Genetik materyal DNA yapısı Semikonservatif replikasyon DNA replikasyonu Replizom:replikasyon makinesi Replikasyon başlangıcı Telomeraz

3
Yeni sentezlenen zincir
DNA çift heliksi Kalıp zincir DNA zincirinin herbiriyeni kopyalar üretmek üzere kalıp görevi üstlenir

4
Frederick Griffith-1928 Streptococcus pneumonia
Sıcaklıkla öldürülmüş S suşları Canlı R suşlarını canlı S suşuna dönüştürür Farelere virulan S suşu enjekte edildiğinde ölür R suşu enjekte edildiğinde yaşar Isıyla öldürülmüş S suşu enjekte edildiğinde yaşar Isıyla öldürülmüş S suşu ve canlı R suşu enjekte edildiğinde ölür Frederick Griffith-1928 Streptococcus pneumonia S suşundaki bir faktör R suşunu transforme ederek S haline dönüştürdü

5
R suşunu virulan yapan faktör DNA’dır.Oswald Avery-1944
Isıyla öldürülmüş S suşundaki hücre ekstrelerinde bulunan DNA bozulursa ve bu ekstre R suşuyla karıştırılırsa canlı nonvirulan R suşları fareyi öldürmez

6
İki set bakteriyofaj T2 kullanıldı
İlk sette protein kılıfı radyoaktif sülfür 35S ile (DNA yapısında bulunmaz) işaretlendi. İkinci sette DNA 32P ile işaretlendi (protein yapısında bulunmaz). Yalnızca işaretli DNA bakterilerden elde edildi. 1952-Hershey-Chase Deneyi: Faj genetik materyaliprotein değil DNA’dır
 işaretlendi. İkinci sette DNA 32P ile işaretlendi (protein yapısında bulunmaz). Yalnızca işaretli DNA bakterilerden elde edildi Hershey-Chase Deneyi: Faj genetik materyaliprotein değil DNA’dır.")
7
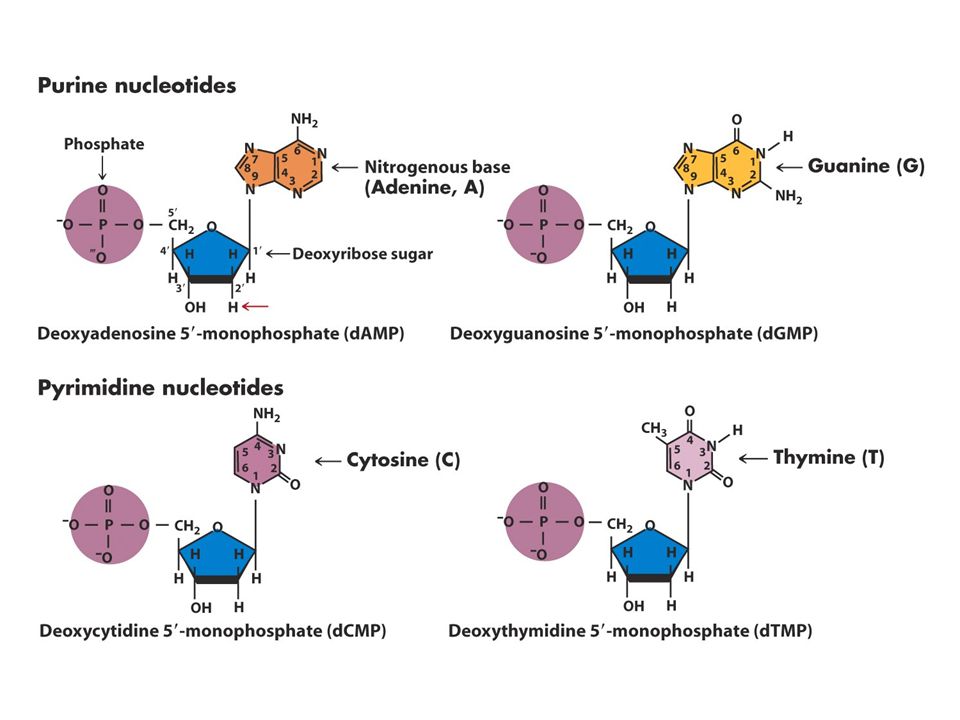
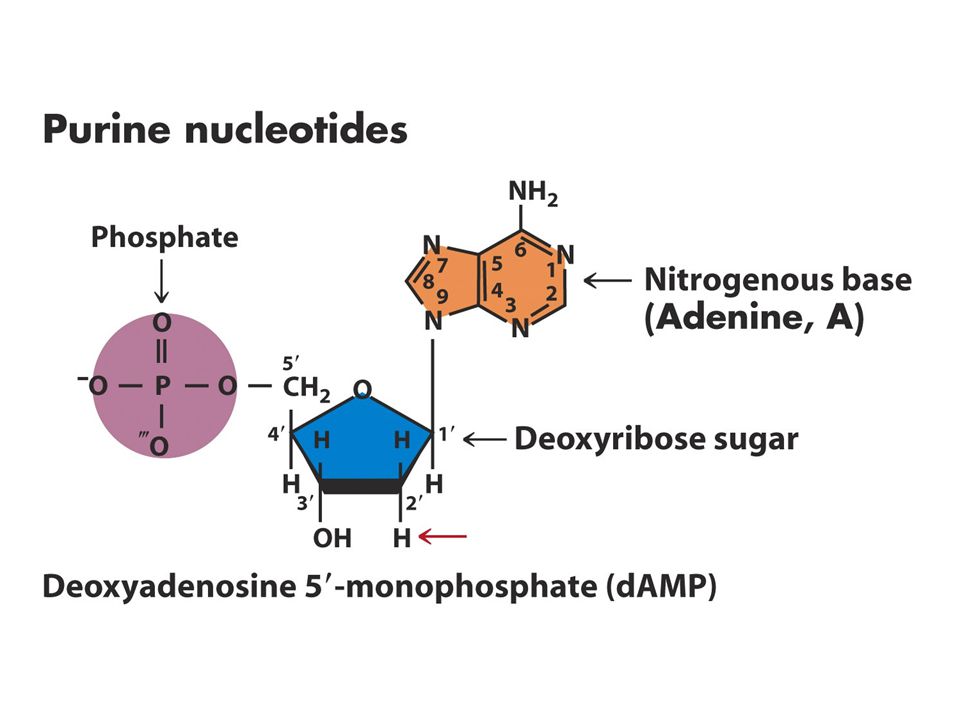
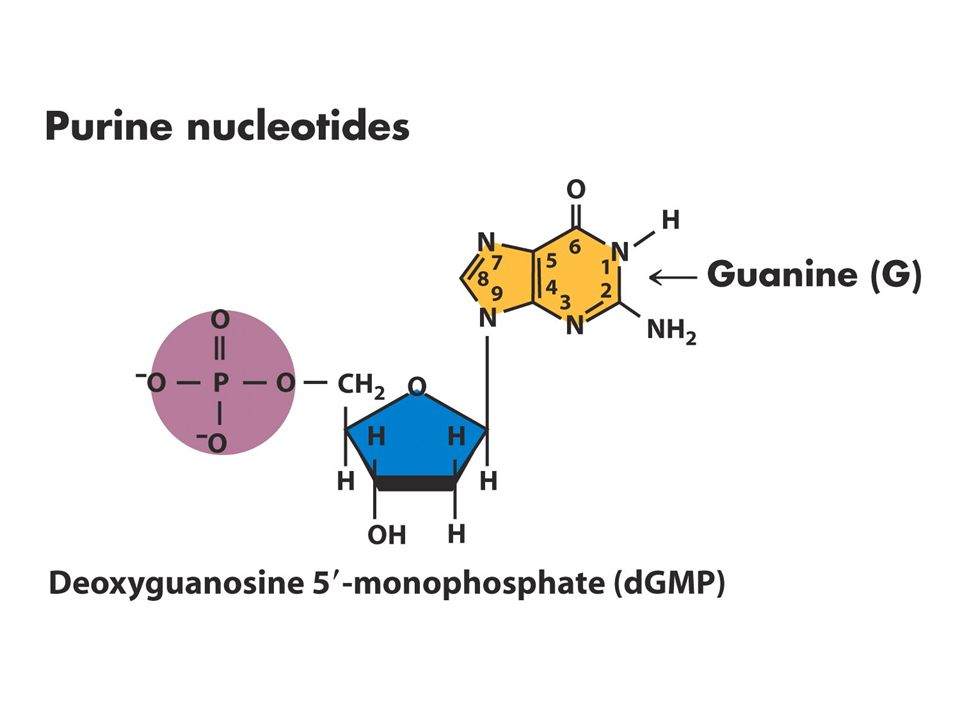
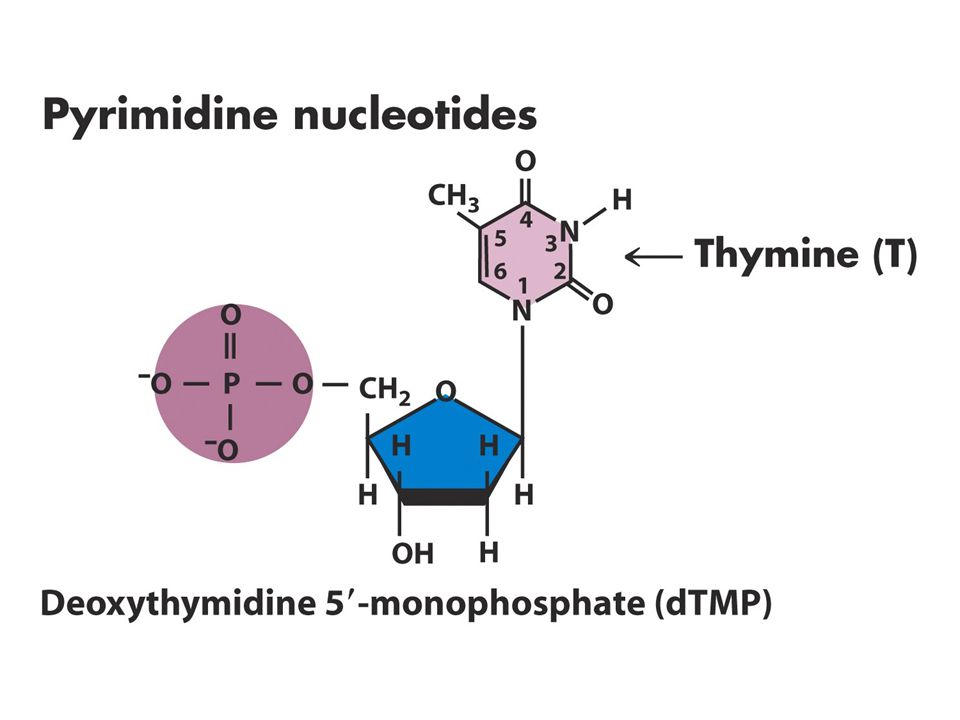
DNA Yapısı Watson-Crick 1953
DNA’nın üç temel bileşeni vardır Fosfat Deoksiriboz Azotlu 4 tane baz Bazlardaki karbon atom sayıları rakamlarla şekerdekiler 1’,2’ biçiminde gösterilir.

13
Erwin Chargaff deneyinde elde edilen sonuçlar Watson-Crick tarafından kullanıldı. DNA’da bulunan herbir nükleotidin miktarını çeşitli organizmalarda belirledi. Toplam primidin nt miktarı toplam purin miktarına eşittir. (T+C=A+G) T miktarı herzaman A miktarına eşittir C de G’ye eşittir. Ancak A+T miktarı G+C miktarına herzaman eşit olmayabilir. Bu oran çeşitli organizmalarda farklılıklar gösterir ama aynı organizmanın farklı dokularında eşittir.
 T miktarı herzaman A miktarına eşittir C de G’ye eşittir. Ancak A+T miktarı G+C miktarına herzaman eşit olmayabilir. Bu oran çeşitli organizmalarda farklılıklar gösterir ama aynı organizmanın farklı dokularında eşittir.")
14
Nükleozid ve nükleotidlerin adlandırılması

15
DNA’nın X- ışını diffraction analizi-Rosalind Franklin
Deneyde DNA fibrilleri üzerine X ışını tutuldu ve ışının serpintileri fotoğraf filminde tutuldu ve ışınlar nokta biçiminde görüntülendi.

16
Film üzerinde herbir noktayı temsil eden serpintinin açısı bir atomun yada DNA’da bulunan belirli atom gruplarının pozisyonu hakkında bilgi verir. Bu prosedur açıklaması zor bir yöntemdir. Buradan elde edilen veriler şöyledir. DNA uzun bir molekül ve birbirine paralel uzanan benzer yapı var. Bu yapı molekül boyunca uzanır (spiral benzeri).
..")
17
1953-James Watson ve Francis Crick Çift zincirli DNA Heliks yapısıyla görüntüleniyorlar.

18
Double helikste yanyana zincirler uzanır
İki nt zinciri herbir zincirdeki bazlar arasında bulunan zayıf etkileşimlerle birarada tutulur ve spiral bir merdivene benzer yapı oluşturur. Her zincirin omurgası fosfat ve deoksiriboz birimlerinin birbirini takibiyle oluşturulur ve bu birimler fosfodiester bağıyla birbirine bağlanır. Fosfodiester bağı bir deoksiribozun 5’ C atomu ile komşu deoksiribozun 3’ C atomu arasında oluşur.

19
Buna göre herbir şeker-P omurgası 5’-3’polarite ve yöne sahiptir
Çift zincirli helikste iki omurga birbirine antiparalel uzanır. Herbir baz omurgada deoksiribozun 1’C atomuna bağlanır ve içe doğru bakar ve karşı zincirdeki baza doğru uzanır. Bazlar arasında H bağları 2 zinciri birarada tutar. Her baz çifti bir purin bir pirimidinden oluşur G, C ile A ise T ile eş oluşturur. G-C içeriği fazla olan DNA daha kararlıdır daha geç denature olur.

20
Şeker-P omurgası ve 3D yapısı
Antiparalel zincirler mavi ile gösterilmiştir bazlar arası H bağları A-T arasında 2 G-C arasında 3 bağ bulunur. 3.34nm Helixin bir dönüşü Bazlar arası mesafe 0.334nm (3.34Angstrom)
")
21
Şeker ve bazdan oluşan birim nükleozid olarak adlandırılır.
Fosfodiester bağı iki komşu deoksiriboz arasındadır. 0.285nm 0.29nm 0.283nm 0.286nm 1.09nm

22
Eksene baz çiftinin 0.292nm dir
Büyük ve küçük oluklar DNA’ya proteinlerin bağlanmasını kolaylaştırırlar. DNA sağa dönümlü heliks yapıdadır. Solüsyondaki form B formudur A formuda sağa dönümlü formdur iki zincir 5’-3’ yönünde antiparalel olarak uzanır DNA %75 kurutulduğunda ve çift zincirli RNA yapısında görülür. Her dönüşte 10.9 baz çifti bulunur ve bazlar heliks eksenine 13 derecelik açıyla yerleşir. Eksene baz çiftinin 0.292nm dir DNA’nın B formundan daha derin büyük oluk görülür. DNA’nın Z formu ise sola dönümlü helikstir ancak her dönüşte 12 bç bulunur ve bazların eksene açısı 8.8 derecedir ve zikzaklı bir yapı biçimindedir. Genellikle CG tekrarlarının çok olduğu DNA bu formu alır. Küçük oluk derindir ve dardır ve büyük oluk yoktur.

23
DNA yapısında baz eşleşmesi
DNA yapısında baz eşleşmesi. Omurga arasındaki mesafe iki pirimidin için büyük iki purin için dardır. Purin-pirimidin çifti bu boşluğu doldurabilir. Kesik çizgiler DNA çapını göstwrmektedir.

24
DNA geridönüşümlü zincir ayrımına gidebilir

25
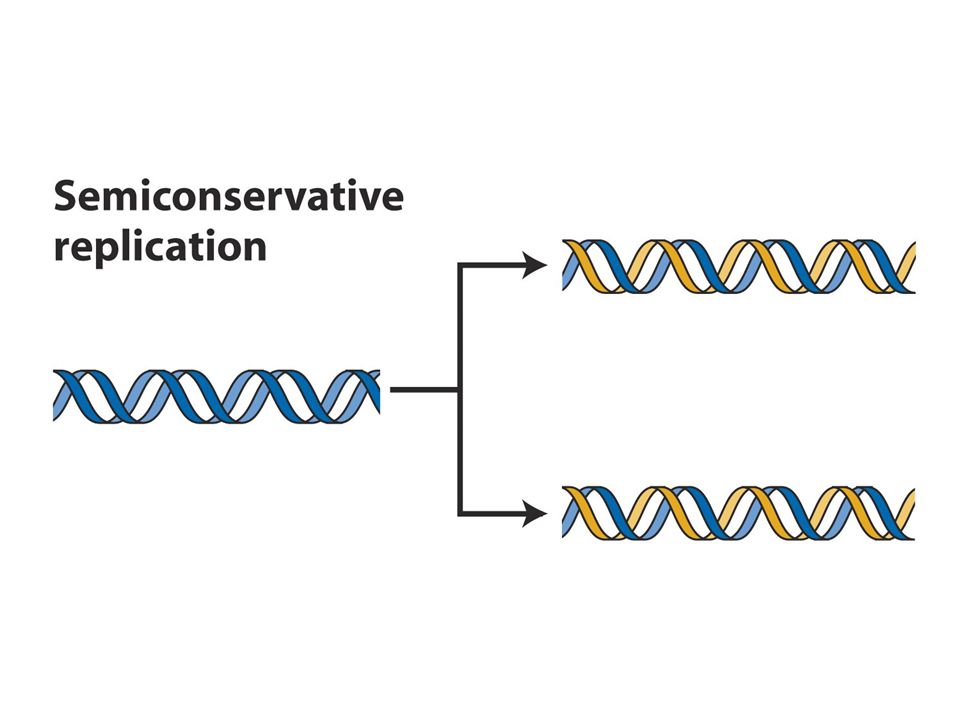
DNA replikasyonunda double heliksin iki zinciri ayrılır ve herbir zincir kalıp görevi görerek semikonservatif modelde replikasyonu sağlar

26
protein ve nükleik asitlerin sentezindeki genel prensipler
Protein ve nükleik asitler belirli sayıda farklı monomerlerin birbirine eklenmesiyle oluşur Herbir polipeptid ve polinükleotid zinciri spesifik bir başlama noktasına sahiptir ve büyüme tek bir yönde belirli bir uca kadar ilerler Primer olarak sentezlenen molekül genelliklşe modifiye edilir

27
Zincirlerin uzaması hem proteinlerde hemde nükleik asitlerde monomerik birimlerin birer birer eklenmesiyle gerçekleşir

28
Protein ve nükleik asitlerin zincirlerinin oluşmasında görünen olası modifikasyonlar

29
DNA degrade eden enzimler
Ekzonükleaz( dıştan içe tek zincirli DNA) Endonükleaz (içten hem tek hem çift zincirli DNA) Restriksiyon enzimler (spesifik diziden)
 Endonükleaz (içten hem tek hem çift zincirli DNA) Restriksiyon enzimler (spesifik diziden)")
30
Komplementer Zincirler
DNA zincirlerinin herbiri diğerinin tam komplementer eşi olduğundan herbir zincir yeni DNA zincirinin sentezi için kalıp olarak görev alır

35
Meselson-Stahl Deneyi.
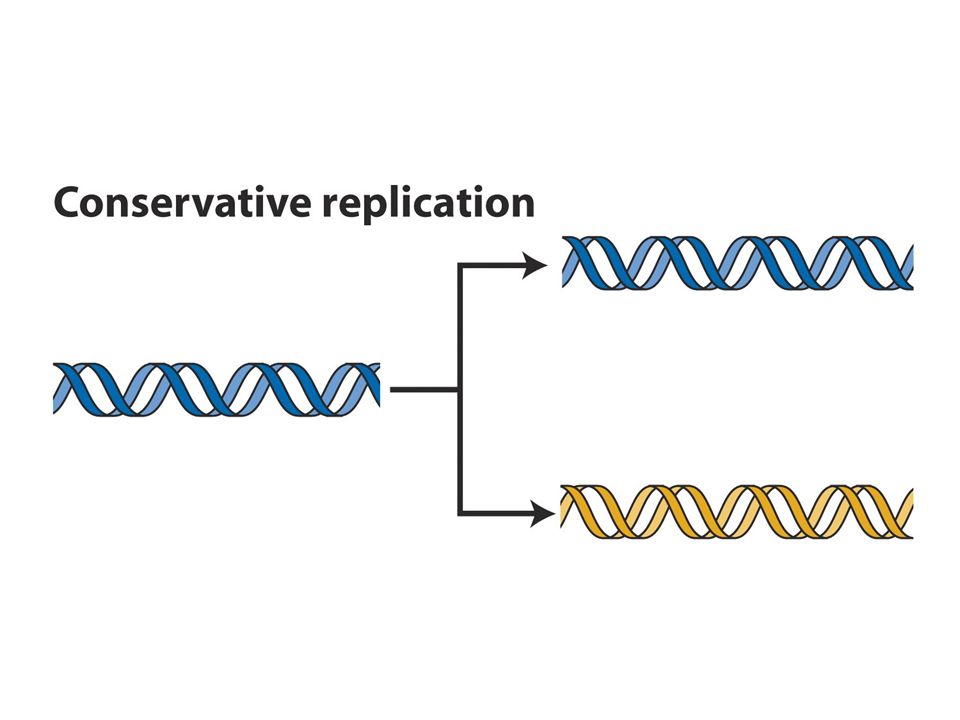
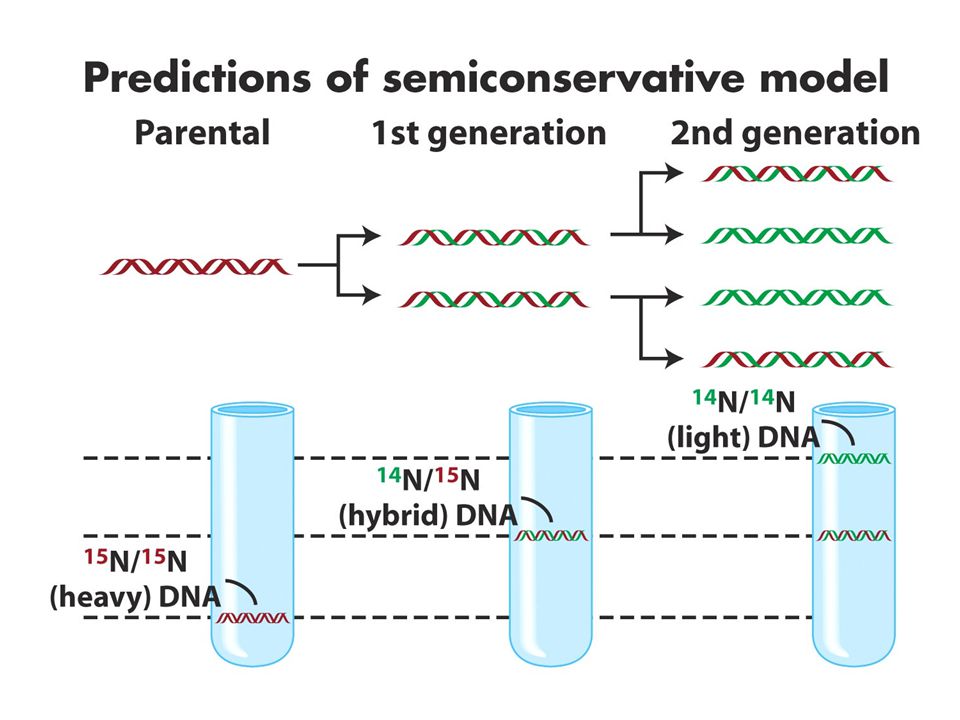
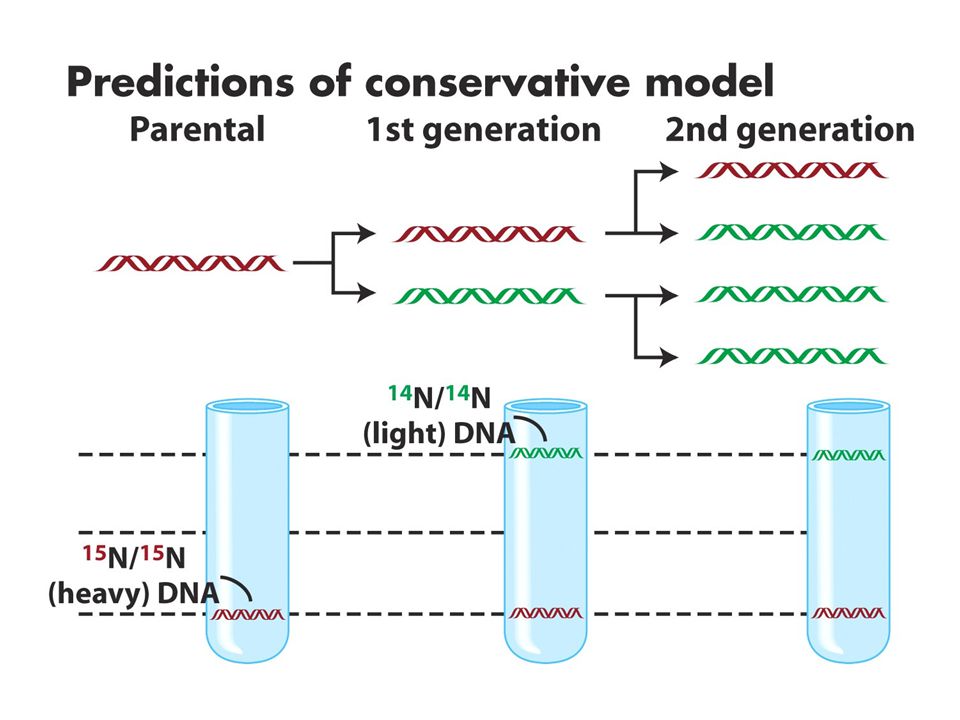
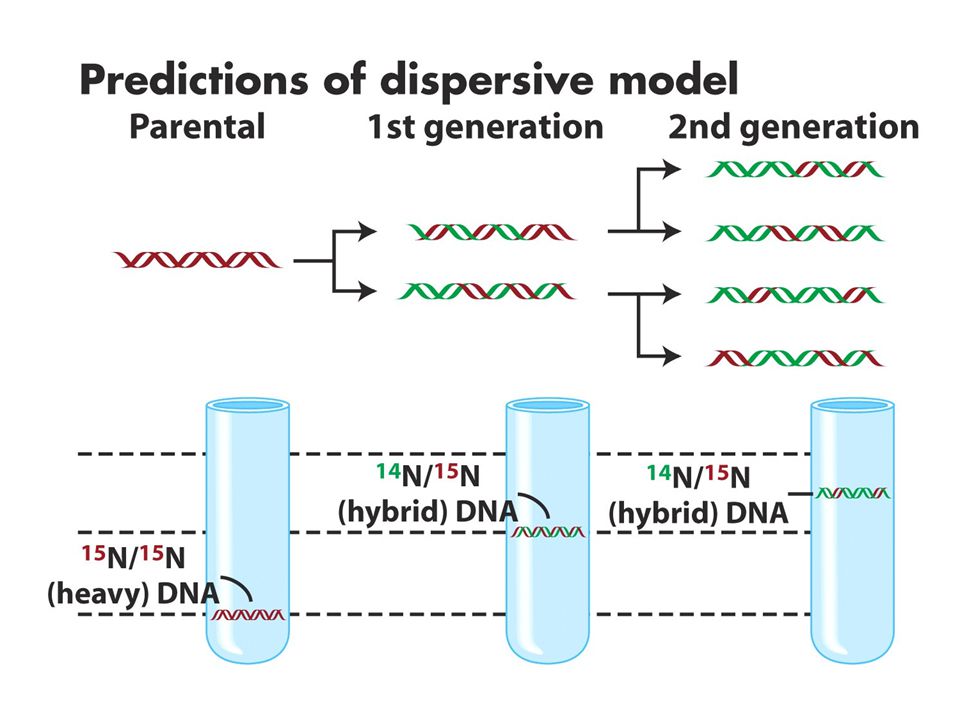
DNA replikasyonu semikonservatiftir. CsCl gradiyenti ile santrifüj dansiteye göre bantlar oluşturur.15N li ortamda çoğalan hücreler 14Nli ortama transfer edildiğinde ilk jenerasyonlar tek bir intermediate bant verirken 2. jenerasyonlar bir intermediate bir hafif bant verir. Bu sonuç semikonservatif modeli destekler. (b ve c konservatif ve dispersiv modeli anlatmaktadır.

39
John Cairns Trityumlu timidinli ortamda bakteri hücrelerini büyüttü (3Htimidin). Buna göre zincirlerden biri radyoaktifli zincir diğeri hafif 2H zincir içermeliydi.

40
Teta yapısı 2. replikasyon döngüsünde replikasyon çatalını göstermektedir

41
DNA polimerazlar 1959 arthur kornber DNA polimerazı izole etti
Enzim dNTPleri büyüyen tek zincirli DNA’nın 3’ ucuna atasal zinciri kalıp olarak kullanarak eklemektedir. Sübstratları dNTPlerdir. E.colide 3 tip pol vardır DNA pol I, II, ve III

42
DNA pol Özellikleri

43
DNA pol Özellikleri Maya DNA pol I, II, III pol α, β ve δ eşdeğerdir, I ve III hücrenin yaşamını sürdürmesi için önemlidir +pol β 20 nt açıklık bulunan DNA moleküllerinin sentezinde önemlidir ve onarımda görev alır FEN1 ökaryotlarda RNA primerini uzaklaştıran 5’-3’exonükelaz aktivitesine sahip enzimdir.E.coli DNA pol I enzimine benzer

44
5’-3’ yönünde zincir uzaması
5’-3’ yönünde zincir uzaması. Bu bağlanma için gerekli enerji trifosfattaki yüksek enerjili bağın kırılmasından elde edilir.

45
Replikasyon çatalı DNA sentezinde heliks boyunca ilerler
Kesintisiz zincir sentezi duraksamadan replikasyon çatalı yönünde gerçekleşir Kesintili zincir sentezi ters yönde ilerler ve replikasyon çatalından uzaklaşarak zincir büyür.

46
Primaz DNA’dan kopyaladığı kısa RNA primerleri sentezler
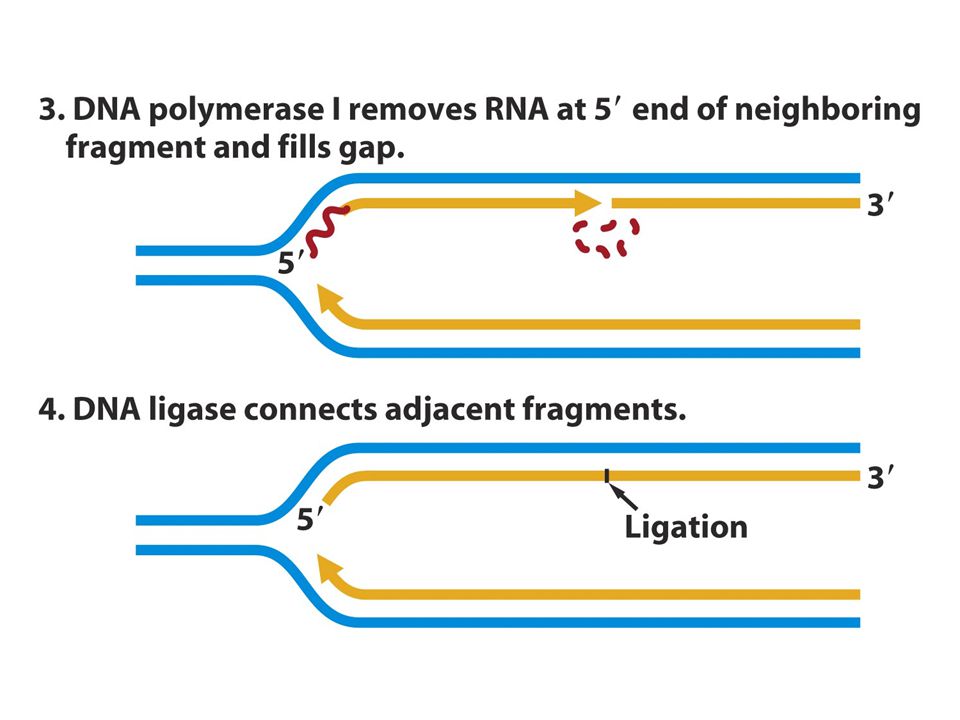
DNA pol IIIRNA primerlerinin ucuna yeni DNA ile uzatır DNA pol I komşu fragmentlerin 5’ ucundaki RNA’ları uzaklaştırır ve boşluğu doldurur. DNA ligaz komşu fragmentleri birleştirir.

47
Kesintili zincir sentezinde 1000-2000nt uzunluğunda okazaki fragmentleri sentezlenir

49
DNA ligaz iki ucu açık DNA zincirlerini yapıştıran tek enzimdir.

50
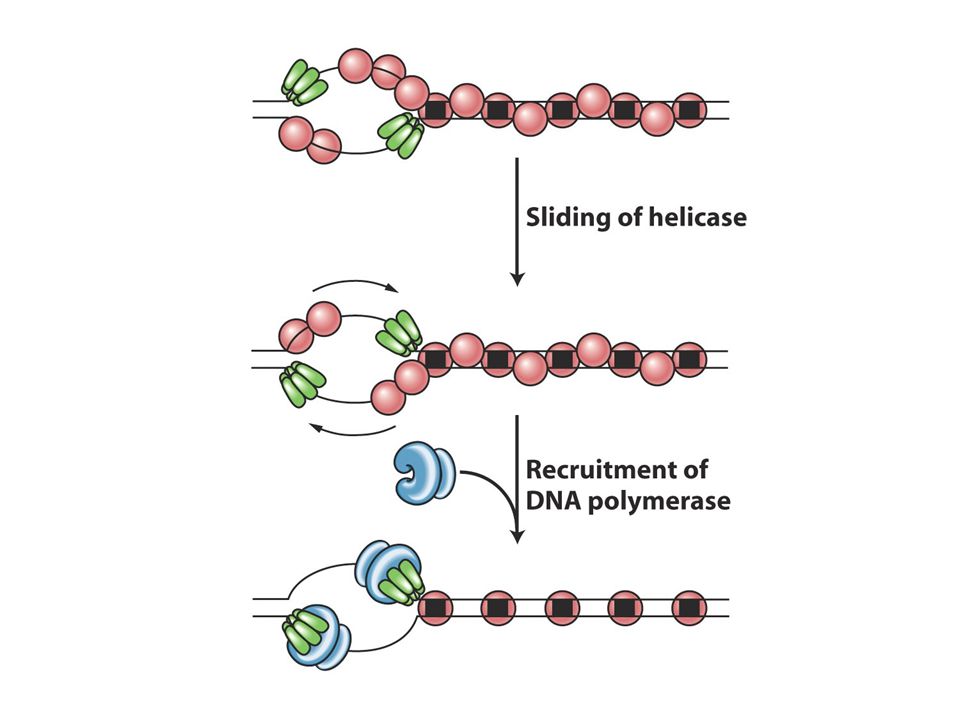
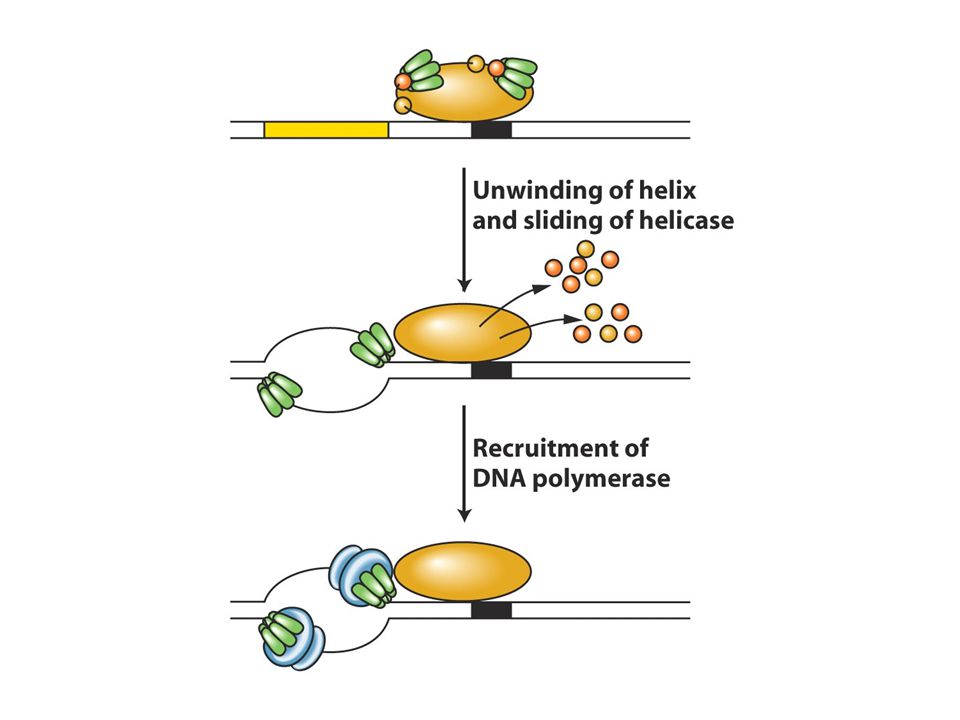
Replizom:DNA replikasyon Makinesi
Topoizomeraz ve helikaz double heliksi açar Açılan tek zincirlere ssbp bağlanır ve iki zincirin yenşden birbirine bağlanmasını önler Primaz kısa RNA primerleri sentezler DNA pol III DNA sentezini gerçekleştirir. E.Colide saniyede 2000 nt eklenir ve genom büyüklüğü 5 milyon bp dir. Herbir replikasyon çatalı saniyede 1000 nt hızda açılır. Pol III çok alt birimli bir yapıdır ve enzimin çekirdeği katalitik aktiviteye sahiptir ve diğer yardımcı proteinlerle birlikte (accessory proteins) buna holoenzim adı verilir. Katalitik çekirdeğin biri kesintili diğeri kesintisiz zincir sentezini yapar. Bazı yardımcı proteinler iki katalitik kor arasında köprü görevi görerek koordineli biçimde kesintili ve kesitntizis sentezi gerçekleştirir. Sliding clamp DNA’yı sarar ve DNA pol IIIün DNA’dan kopmamasını sağlar. Onun sayesinde DNA’dan düşmeden 10 nt ekleyen DNApol III (distributive) düşmeden 10binlerce nt ekleyen bir forma (processive) dönüşür. Primaz clampa dokunmaz dolayısıyla primaz distrubitve enzimdir
 buna holoenzim adı verilir. Katalitik çekirdeğin biri kesintili diğeri kesintisiz zincir sentezini yapar. Bazı yardımcı proteinler iki katalitik kor arasında köprü görevi görerek koordineli biçimde kesintili ve kesitntizis sentezi gerçekleştirir. Sliding clamp DNA’yı sarar ve DNA pol IIIün DNA’dan kopmamasını sağlar. Onun sayesinde DNA’dan düşmeden 10 nt ekleyen DNApol III (distributive) düşmeden 10binlerce nt ekleyen bir forma (processive) dönüşür. Primaz clampa dokunmaz dolayısıyla primaz distrubitve enzimdir.")
51
Replizom:DNA replikasyon Makinesi

52
Açılan atasal dublex Aşırı sarmal bölge

53
Bir tür topoizomeraz olan DNA giraz süpersarmallı yapıda kesik oluşturur ve replikasyon çatalı önündeki gerilmeyi azaltır. Sonra kırığı yeniden onarıır. Topoizomerazlar DNA yapısına süpersarmallar ekleyen yada onları kırarak DNA gerilimini azlatan enzimlerdir. Kırıklar tek yada çift zincirde oluşturulabilir.

54
Replikasyon sırasında nükleozomların toplanması:
Ökaryot ve prokaryotlarda replikasyon semikonservatiftir ve herikisinde de kesintili/siz zincir sentezi vardır. Replizomdaki bileşenler prokaryotlarda 13 ökaryotlarda 27 tanedir (bugüne kadar belirlenmiş olan). Ökaryot DNA’sı çekirdekte kromatin olarak bulunur ve kromatinin temel birimi nükleozomdur. Sentez sırasında nükleozomlar atasal zincirden ayrılır bir yandan da yeni zincir için yenileri sentezlenir. Replizomda bir yandan eski histonlar rastgele yeni sentezlenen zincirler üzerine dağıtılır ve yeni sentezlenen histonlar ise chromatin assembly factor I aracılığıyla gönderilir. CAF1 histonlara bağlanır ve onları replikasyon çatalına hedefler ve orada yeni sentezlenen DNA ile bağlanır. CAF1 ve taşıdığı histonlar replikasyon çatalına clamp proteininin ökaryotik versiyonu olan proliferating cell nuclear antigen denen bir proteine bağlanarak replikasyon çatalına ulaşır.
. Ökaryot DNA’sı çekirdekte kromatin olarak bulunur ve kromatinin temel birimi nükleozomdur. Sentez sırasında nükleozomlar atasal zincirden ayrılır bir yandan da yeni zincir için yenileri sentezlenir. Replizomda bir yandan eski histonlar rastgele yeni sentezlenen zincirler üzerine dağıtılır ve yeni sentezlenen histonlar ise chromatin assembly factor I aracılığıyla gönderilir. CAF1 histonlara bağlanır ve onları replikasyon çatalına hedefler ve orada yeni sentezlenen DNA ile bağlanır. CAF1 ve taşıdığı histonlar replikasyon çatalına clamp proteininin ökaryotik versiyonu olan proliferating cell nuclear antigen denen bir proteine bağlanarak replikasyon çatalına ulaşır.")
55
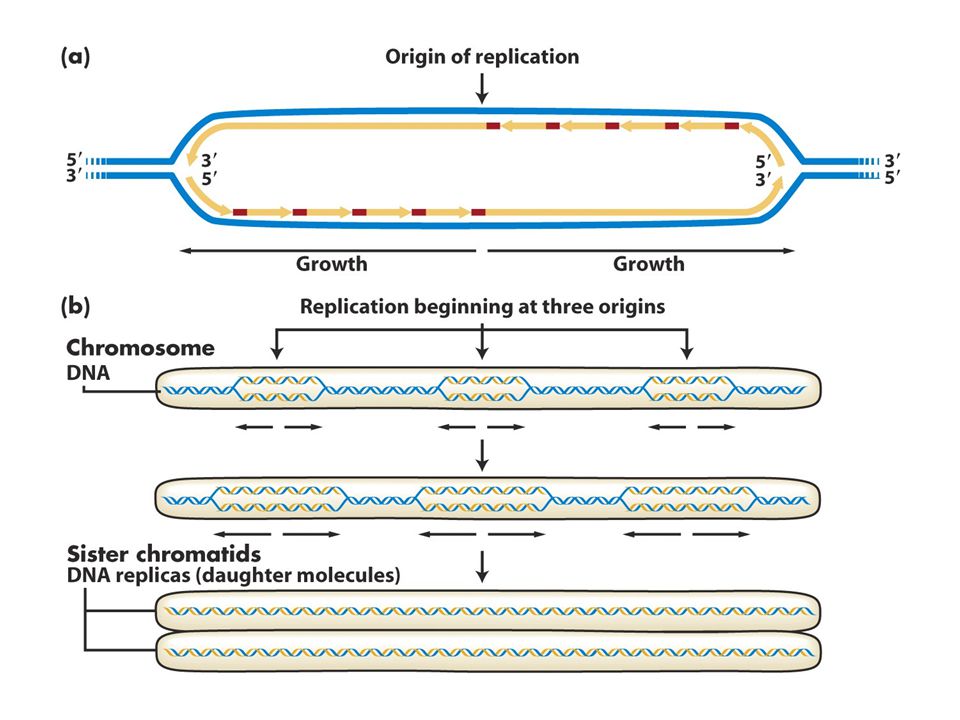
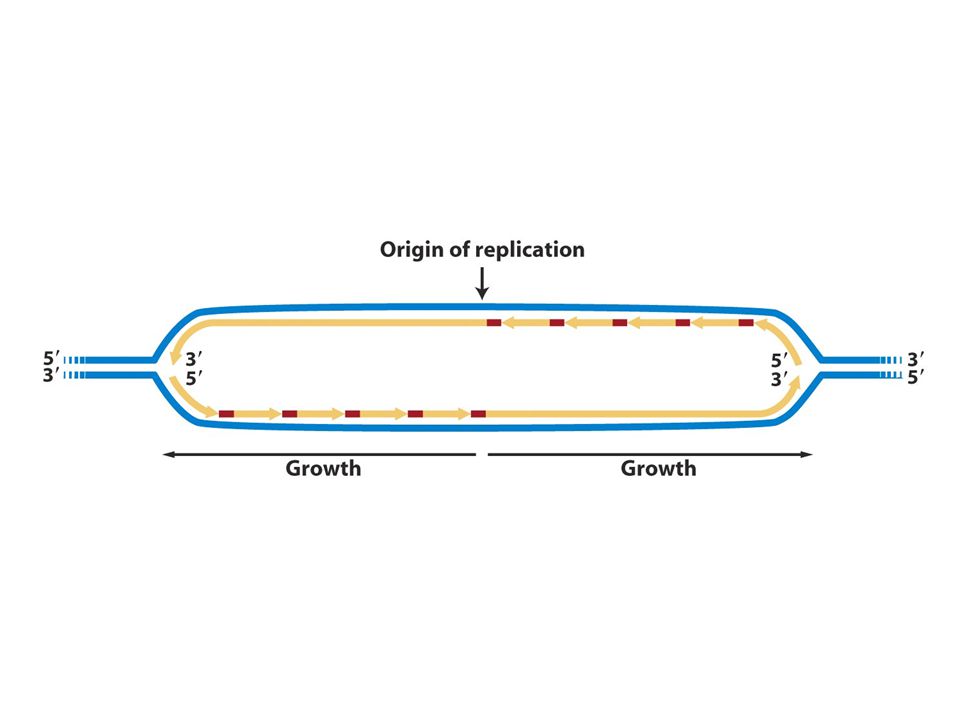
Replikasyonun başlangıcı
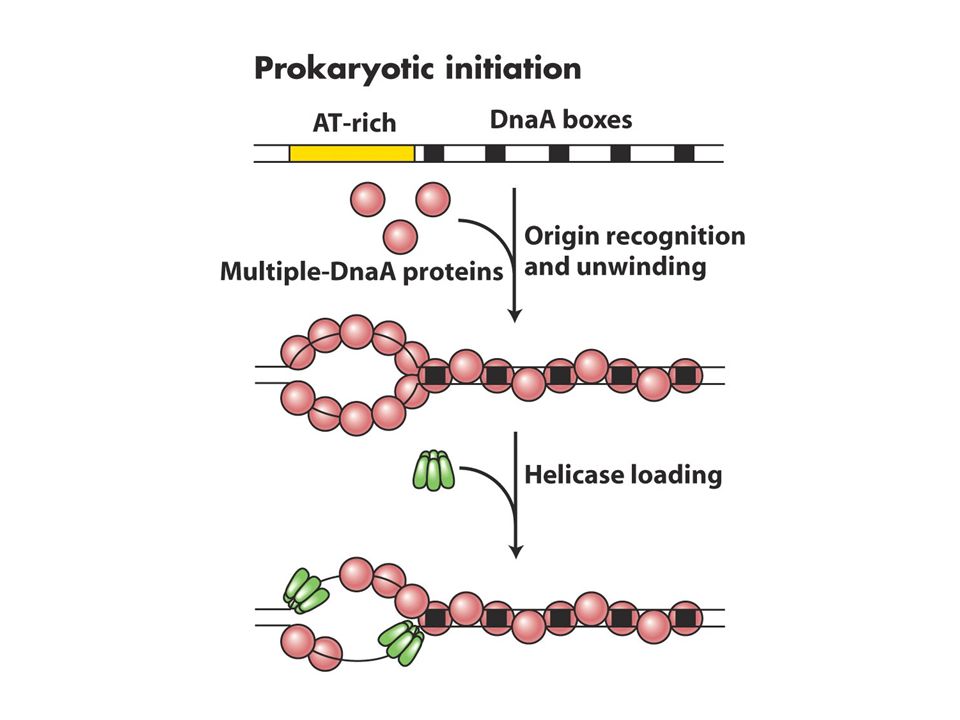
Pro ve ökaryotlardaki replizom kümeleşmesi kromozomda belirli bir noktadan (orijin) başlar ve hücrenin hayatında belirli bir zamanda gerçekleşir. E.Coli replikasyonu sabit bir orijinden başlar ve buaraya oriC adı verilir ve iki yönde ilerler. DnaA kutusu denen 13bç lik bölgeye DnaA proteininin bağlanması replizom oluşumundaki ilk evredir. Bu 13bç lik çerçeve oriC’de 5 kez yinelenir. A ve T nt lerince zengin bölgede DnaAnın bağlanması buranın açılmasına neden olur. Açılma ilerledikçe daha fazla DnaA tek zincirli DNA’ya bağlanır. DnaA oriC’yi kapladığında iki tane helikaz (DnaB proteini) bağlanır ve 5-3’ yönünde kaymaya başlar ve replikasyon çatalında heliksi açmaya başlar. DNA pol III holoenzim ve primaz protein-protein etkileşmlerini kullanarak replikasyon çatalına bağlanır sentez başlar.
 başlar ve hücrenin hayatında belirli bir zamanda gerçekleşir. E.Coli replikasyonu sabit bir orijinden başlar ve buaraya oriC adı verilir ve iki yönde ilerler. DnaA kutusu denen 13bç lik bölgeye DnaA proteininin bağlanması replizom oluşumundaki ilk evredir. Bu 13bç lik çerçeve oriC’de 5 kez yinelenir. A ve T nt lerince zengin bölgede DnaAnın bağlanması buranın açılmasına neden olur. Açılma ilerledikçe daha fazla DnaA tek zincirli DNA’ya bağlanır. DnaA oriC’yi kapladığında iki tane helikaz (DnaB proteini) bağlanır ve 5-3’ yönünde kaymaya başlar ve replikasyon çatalında heliksi açmaya başlar. DNA pol III holoenzim ve primaz protein-protein etkileşmlerini kullanarak replikasyon çatalına bağlanır sentez başlar.")
56
Prokaryotlardaki ve ökaryotlardaki replikasyon orijinleri (bakteri-maya)
")
59
Bakterilerde replikasyon döngüsü 20-40dk da biter ama ökaryotlarda 1
Bakterilerde replikasyon döngüsü 20-40dk da biter ama ökaryotlarda 1.4 saat mayada 24 saat kültüre hücrelerde ve saat diğer hücrelerde sürer.Çünkü sadece birtek kromozomda sentez olmaz. Mayada oriC bakteridekine benzer AT zengin bölgeye başlangıç proteinleri bağlanır ve sentez başlar. Yüksek yapılı organizmalarda orijin tam olarak belirlenmemiştir.Ancak bin hatta onbinlerce nt uzunlukta olabilir. Bakteri aksine birçok replikasyon orijini bulunur. İnsanın 23 kromozomunda binlerce replikasyon çatalı belirlenmiştir. Dolayısıyla birçok orijinden iki yönde birçok replikasyon çatalında sentez sürer.Ökaryotlardaki DNA sentezi sadece hücre döngüsünün S fazında olur.

63
DNA pol mayada 3 noktada senteze başlar çift yönde ilerler zincir uzar sonra kardeş kromatidler oluşur. Replizomda kümeleşme için 3 protein gerekir. Origin recogniton complex (ORC) maya orijinine bağlanan ilk proteindir. (DnaA proteini gibi) Daha sonra cdc6 ve Cdt1 buraya bağlanır. ORC ve bu 2 protein daha sonra MCM kompleksi denen replikatif helikazın bağlanmasına aracılık eder. cdc6 ve Cdt1 mitozun geç evrelerinde ve G1de sentezlenir sentez başladıktan sonrada yıklır. Sentez başladıktan sonra orijinde yeni replizomlar oluşmaz.
 maya orijinine bağlanan ilk proteindir. (DnaA proteini gibi) Daha sonra cdc6 ve Cdt1 buraya bağlanır. ORC ve bu 2 protein daha sonra MCM kompleksi denen replikatif helikazın bağlanmasına aracılık eder. cdc6 ve Cdt1 mitozun geç evrelerinde ve G1de sentezlenir sentez başladıktan sonrada yıklır. Sentez başladıktan sonra orijinde yeni replizomlar oluşmaz..")
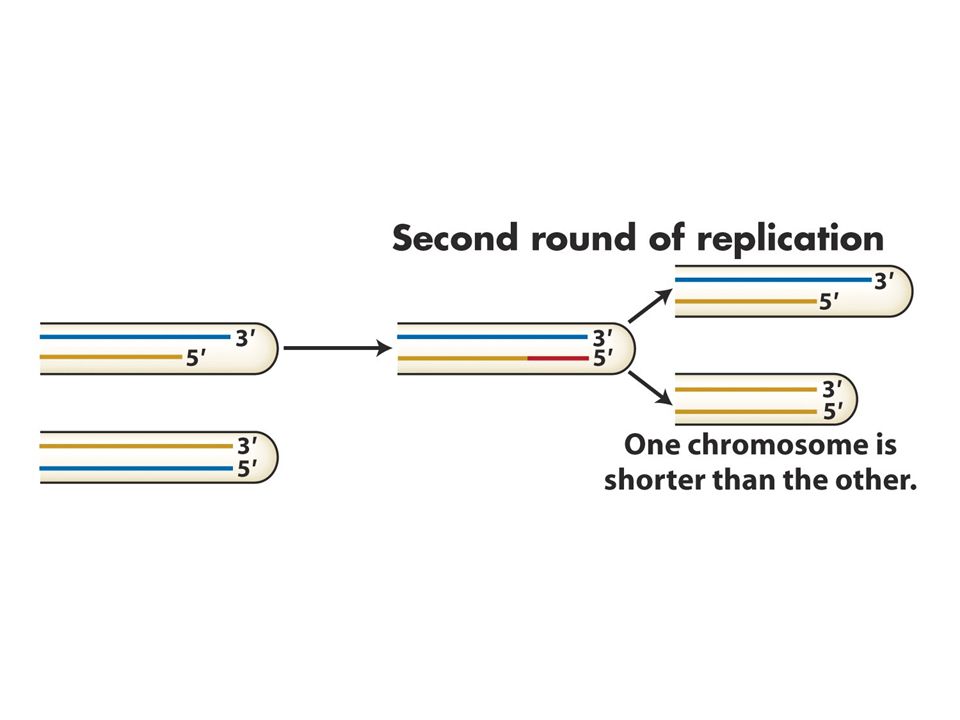
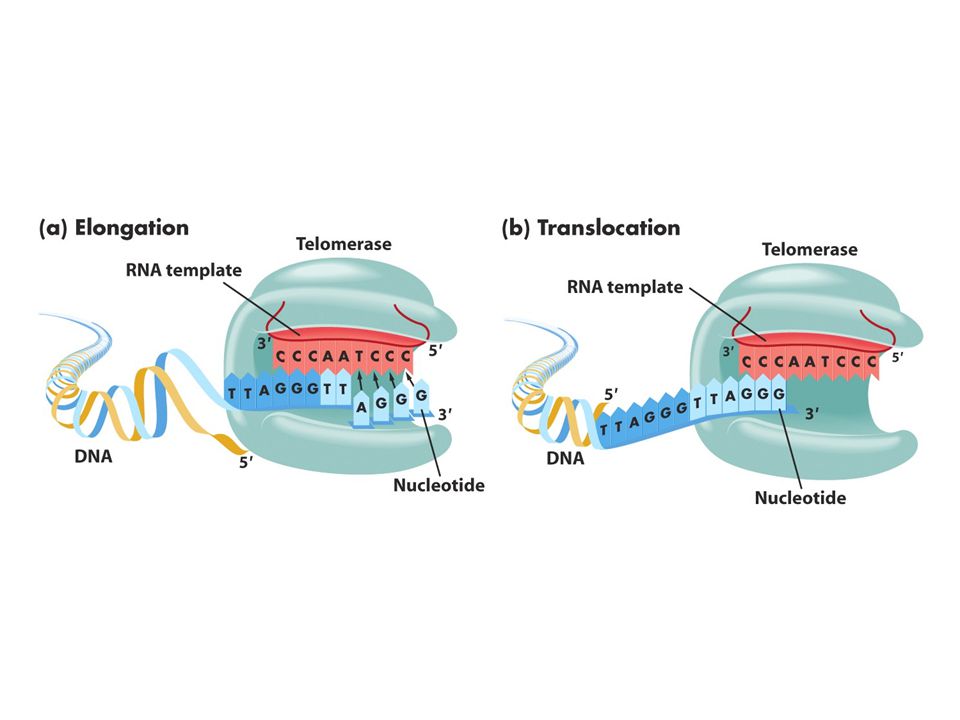
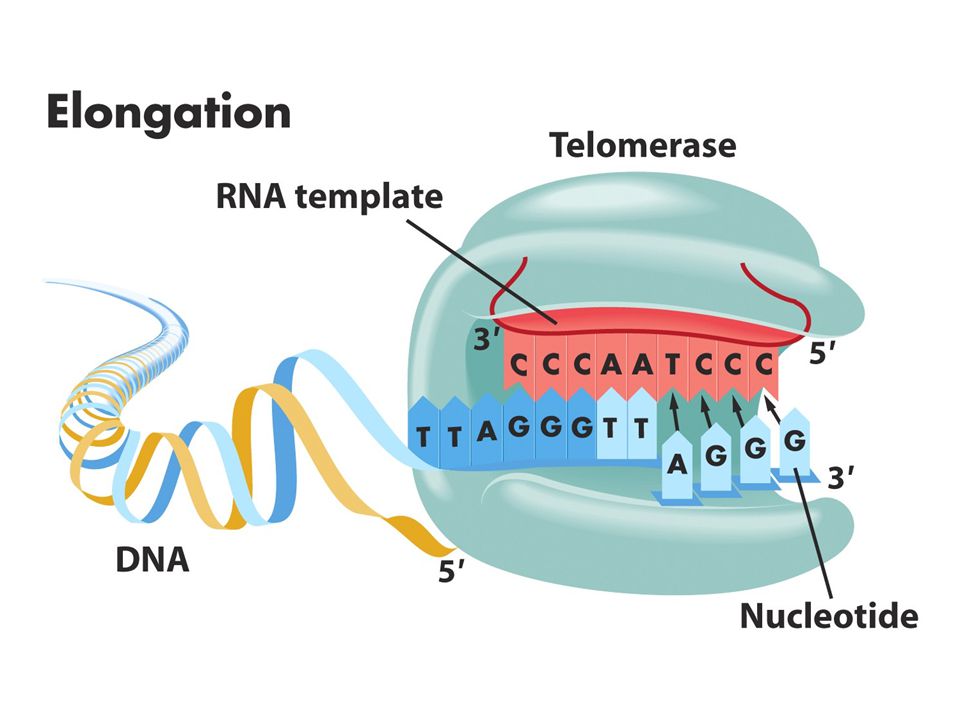
65
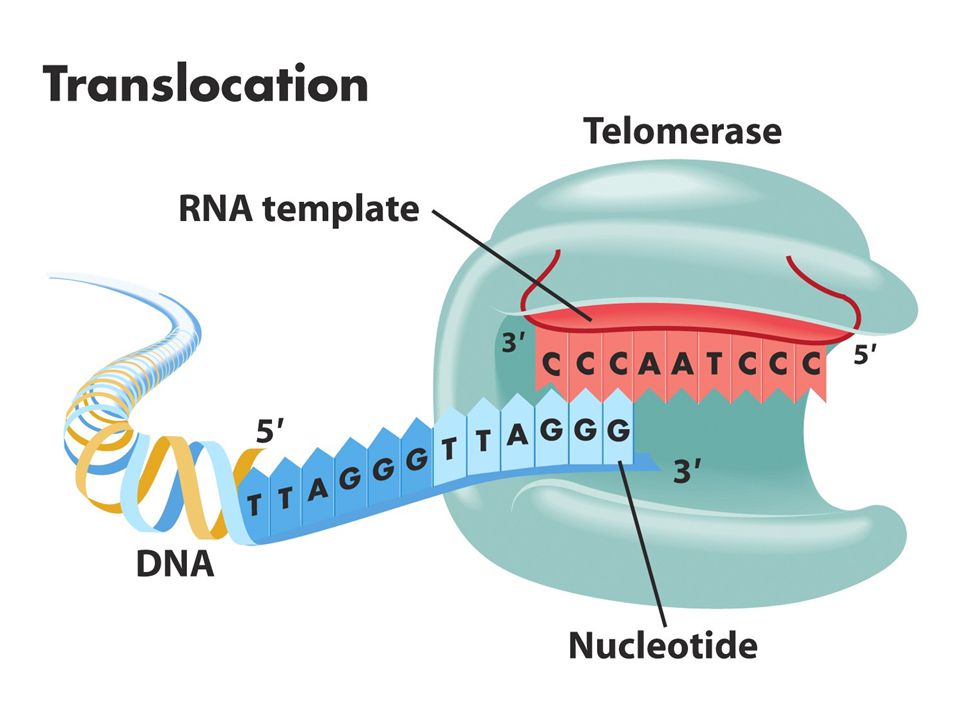
Kromozom uçlarında replikasyon sorunu: kromozon uçlarında (telomer) replikasyon sırasında kesintili zincir sentezinde kromozomun bir kısmı kaybolur. Kesintisiz zincir sentezi kromozomun ucuna kadar sürer.Kesintili zincir sentezinde kromozomun ucunda yer alan RNA primeri uzaklaştırılınca 2. replikasyon döngüsünde koromozomlardan biri kısalır.Sonunda o bölgede bulunan çeşitli genler kaybolur.Hücreler bu sistemi düzeltecek mekanizmaya sahiptir.Bir enzim (telomeraz) kodlanmayan bir diziyi kromozoun ucuna ardıl biçimde ekler. Tetrahymena denen birhücreli organizmada TTGGGG dizisinin birçok kopyası her kromozomun 3’ ucuna eklenir. İnsanlardaki bu dizi TTAGGG’dir. Bu dizi nonkoding DNA’dır. Telomeraz enzimi bu diziyi ekler. Telomeraz küçük bir RNA molekülü taşır ve bu RNA dizisinin bir kısmı telomerik tekrar dizileri için kalıp görevi görür. İnsanlardaki bu RNA dizisi 3’AATCCCdir.
 kodlanmayan bir diziyi kromozoun ucuna ardıl biçimde ekler. Tetrahymena denen birhücreli organizmada TTGGGG dizisinin birçok kopyası her kromozomun 3’ ucuna eklenir. İnsanlardaki bu dizi TTAGGG’dir. Bu dizi nonkoding DNA’dır. Telomeraz enzimi bu diziyi ekler. Telomeraz küçük bir RNA molekülü taşır ve bu RNA dizisinin bir kısmı telomerik tekrar dizileri için kalıp görevi görür. İnsanlardaki bu RNA dizisi 3’AATCCCdir..")
Benzer bir sunumlar











>")



 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")
>")

>")



