Sunuyu indir
Sunum yükleniyor. Lütfen bekleyiniz

1
Introduction to Genetic Analysis
Anthony Griffiths • Susan Wessler • Richard Lewontin William Gelbart • David Suzuki • Jeffrey Miller Introduction to Genetic Analysis Eighth Edition Chapter 14: Mutation Copyright © 2005 by W. H. Freeman & Company

2
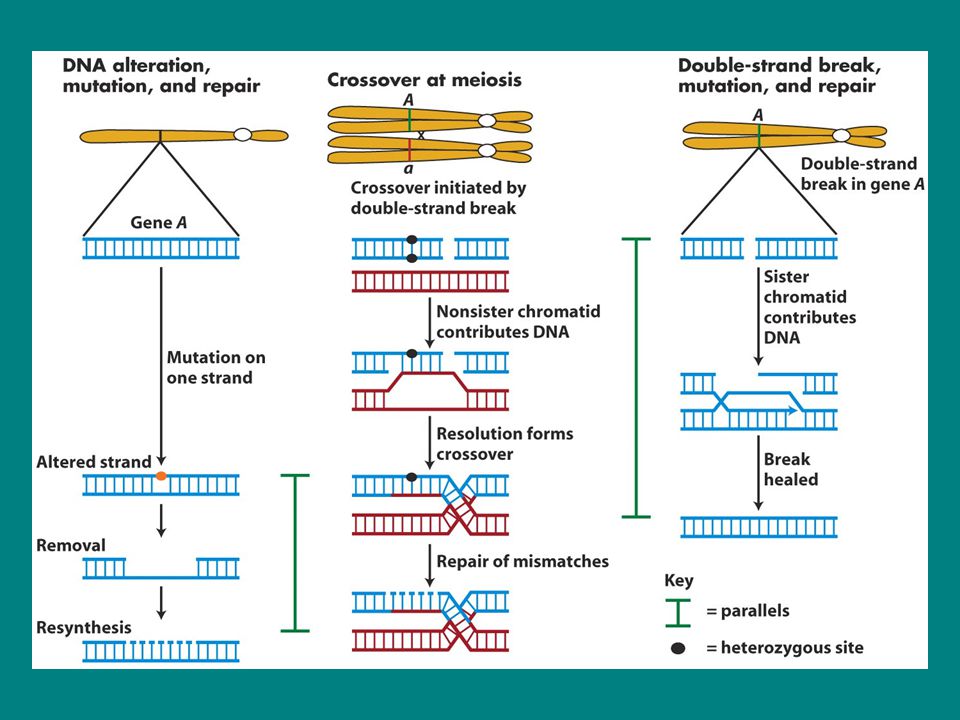
Holliday junction

3
MUTASYONLAR Bir genin DNA dizisindeki değişime mutasyon denir. Mutasyonlar evrimsel değişimin kaynaklarıdır. organizmalarda yeni alleller ortaya çıkar bunların bazıları spontane bazıları da radyasyon yada kimyasallar sonucu gerçekleşir. Mutasyon sonucu ortaya çıkan yeni allel 2. düzey çeşitlilik için hammadde olur çünkü rekombinasyondan etkilenir. Genetik çeşitlilik bireylere evrim için hammadde sağlar. Genetik çeşitlilik nasıl oluşur? Mutasyonlar Rekombinasyonlar

4
Rekombinasyon hücresel bir süreçtir farklı genlerin allellerinin yeni kombinasyonlarla yeni bir grup oluşturmasıdır. Bir benzetme yapacak olursak mutasyonlar yeni oyun kartları yapar rekombinasyonlar ise bu oyun kartlarının farklı ellere yeni kombinasyonlar oluşturacak biçimde dağıtılmasıdır. DNA hücrelerde kararlı değildir ve mutasyona uğrama olasılığı vardır.Bu derste kromozom mutasyonlarından öte gen mutasyonları anlatılacaktır. DNAda çeşitli gen mutasyonları görülür, bu mutasyonlar bir bazın değişimi kadar basit üçlü nt lerin tekrarları biçiminde karmaşık olabilir. Mutasyonlar mobil genetik elemanların genom içinde başka bir yere entegre olmasıyla da görülebilir. DNA üzerinde hücrede birçok savaş gerçekleşir. Bir yandan çeşitli moleküller DNA’da hasar oluştururken hücre onarım sistemi bu hasarları gidermeye çalışır. DNA yapısına mutasyonlar çoğunlukla çeşitli ajanlarla sokulur ki bunlara mutajen denir.Mutasyonlar spontane de olabilirler ancak daha az sıklıkta görülür.Ancak evrimsel açıdan bu mutasyonlar çok önemlidir.

5
Gen Mutasyonları genel anlamda iki başlık altında incelenebilir.
Hücreler hasarlı DNA’yı onaracak biçimde evrimleşmiştir. Hücrelerde çeşitli onarım sistemleri vardır ve bunların çoğu da DNA’nın komplementer olma özelliğine güvenerek onarımı gerçekleştirir. Yani bir zincir kalıp olarak kullanır ve diğerini onarır. Gen Mutasyonları genel anlamda iki başlık altında incelenebilir. DNA’nın tek bir baz çiftini etkileyen mutasyonlar (Nokta mutasyonlar) Bir gen içerisinde küçük tekrarlı dizilerin kopya sayılarını değiştiren mutasyonlar.
 Bir gen içerisinde küçük tekrarlı dizilerin kopya sayılarını değiştiren mutasyonlar.")
6
Nokta Mutasyonlar DNA’da tek baz çiftinin değişmesidir
Genin işlevi kaybetme olasılığı yeni bir genin işlev kazanmasından daha olasıdır. Ancak işlev kazanma olasılığı da vardır

7
Nokta mutasyonların Orijini
Yeni oluşan mutasyonlar iki gruba ayrılır Spontan mutasyonlar İndüklenen mutasyonlar Spontan mutasyonlar kendiliğinden oluşurken indüklenen mutasyonlar mutajen yada çevresel ajanlarla amaçlı olarak mutasyonun teşvik edilmesidir. Spontan mutasyonlar evrimsel çeşitlilik sağlarlar.

8
Spontan mutasyon frekansı oldukça düşüktür bir hücrede yaklaşık hücrede bir görülür. Dolayısıyla daha fazla mutasyon isteniyorsa mutlak indüklemek gerekir. Mutajenlere maruz bırakarak mutasyon üretmeye mutagenez denir ve bu organizmaya da mutagenize organizma denir.

9
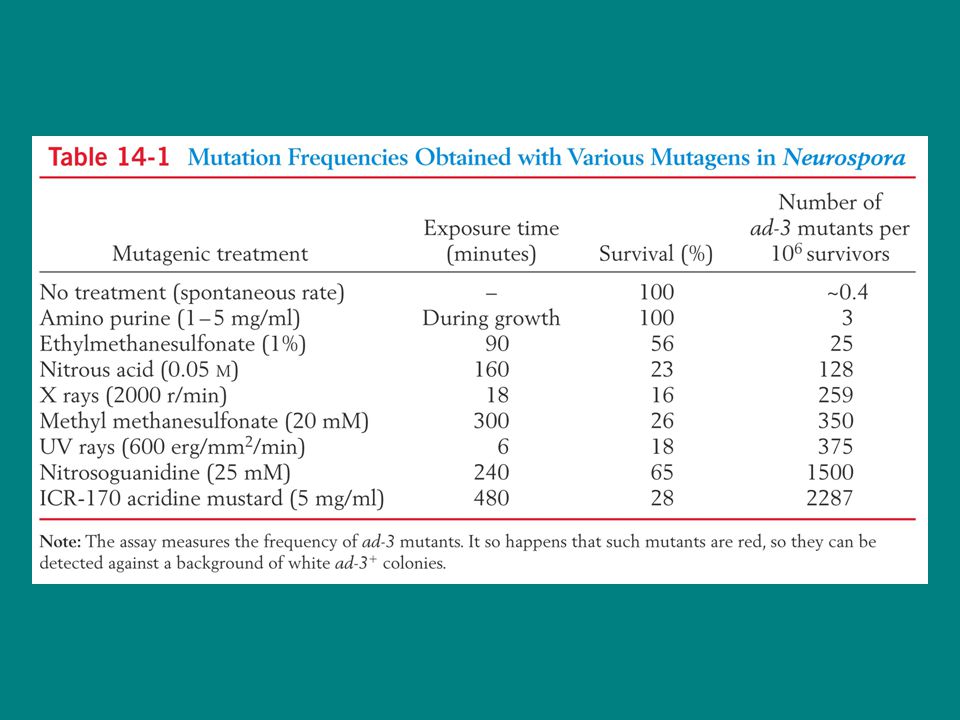
En sık kullanılan mutajenler Yüksek enerjili radyasyon
Çeşitli kimyasallar Mutajenin dozu nekadar yüksekse mutasyon freakansı da okadar yüksek olur

12
Nokta Mutasyon Tipleri I DNA düzeyinde
Transisyon Purin başka bir purinle yada primidin başka bir pirimidinle değişir Transversiyon Purin yerine pirimidin yada pirimidin yerine purin geçer İndel Bir yada birkaç bazın DNA dizisine eklenmesi yada çıkarılmasıdır.

13
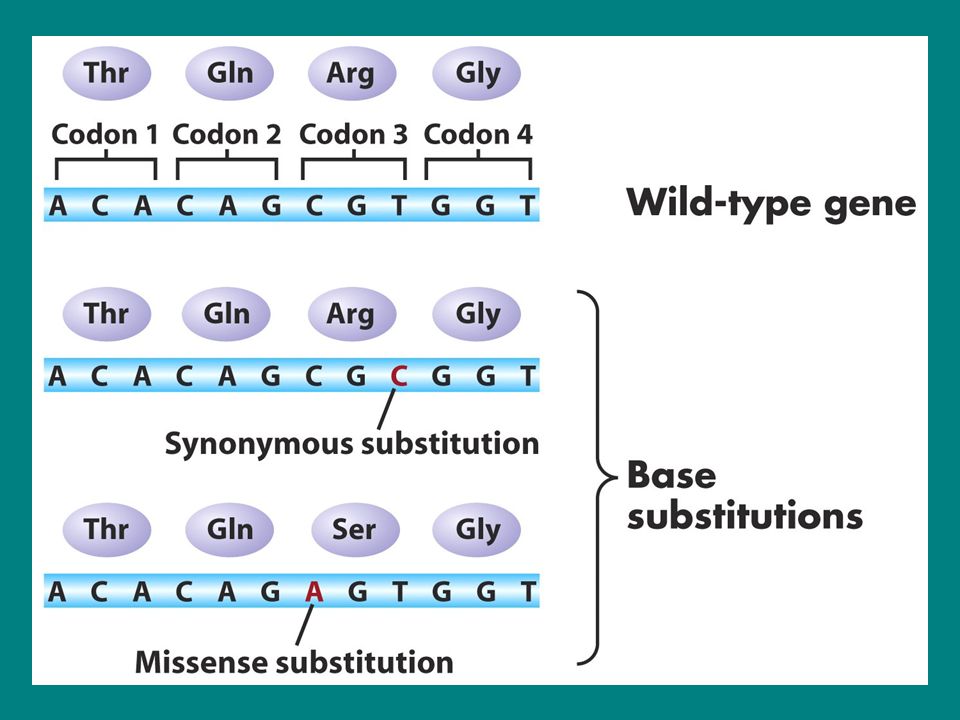
Nokta Mutasyon Tipleri II Protein düzeyinde
Sinonim mutasyonlar (Silent-sessiz mutasyon) Kodon aynı aa belirler AGG (Arg)-CGG(Arg) Missens mutasyon (non sinonim mutasyon) Konservatif missense mutasyon Kodon farklı ancak kimyasal olarak benzer aa belirler AAA(Lys-bazik)-AGA (Arg-bazik) Nonkonservatif missense mutasyon Protein işlevini değiştirmez ve kimyasal yapısı farklı aa belirler UUU(hidrofobik fenilalanin)-UCU(polar serin) Nonsens mutasyon Stop kodon oluşur Frameshift mutasyon Bir baz çifti eklenmesi AAG ACT CCT-AAG AGC TCC T… Bir baz çifti delesyonu AAG ACT CCT-AAA CTC CT…
 Kodon aynı aa belirler. AGG (Arg)-CGG(Arg) Missens mutasyon (non sinonim mutasyon) Konservatif missense mutasyon. Kodon farklı ancak kimyasal olarak benzer aa belirler. AAA(Lys-bazik)-AGA (Arg-bazik) Nonkonservatif missense mutasyon. Protein işlevini değiştirmez ve kimyasal yapısı farklı aa belirler. UUU(hidrofobik fenilalanin)-UCU(polar serin) Nonsens mutasyon. Stop kodon oluşur. Frameshift mutasyon. Bir baz çifti eklenmesi. AAG ACT CCT-AAG AGC TCC T… Bir baz çifti delesyonu. AAG ACT CCT-AAA CTC CT…")
14
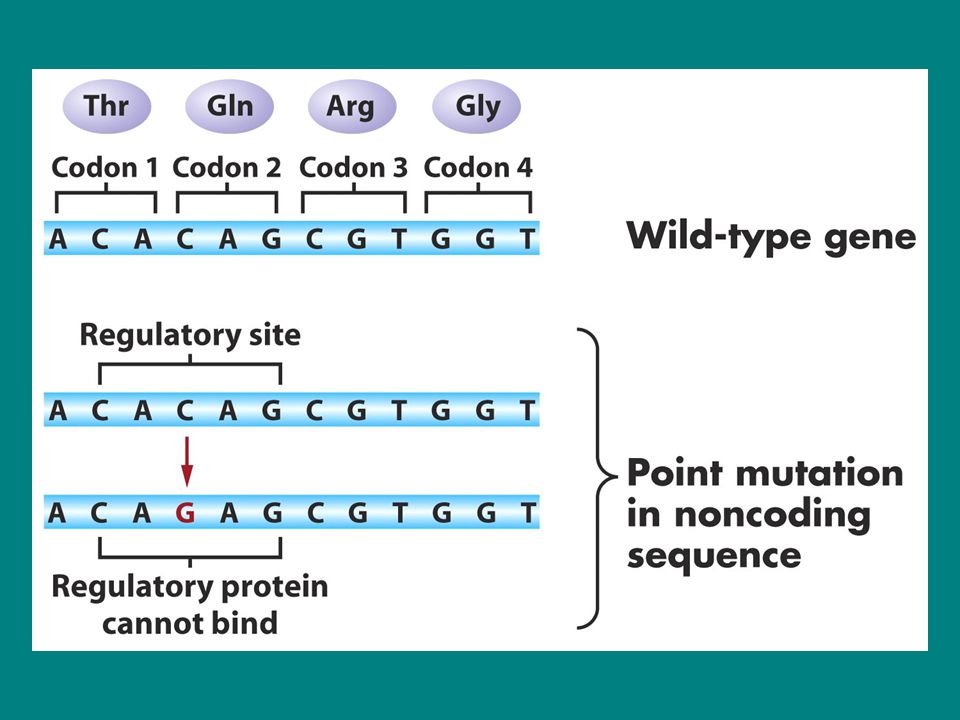
Doğal tip gen Baz değişimi Frameshift mutasyon Nonkoding bölgede
Nokta mutasyon Bir gende meydana gelen nokta mutasyonun sonuçları

18
Doğal tip gen Missens Mut Ör:G.C-A.T Nonsens mut Ör:CAA-TAA Frameshift mut Ör: +A Regülatör bölge mutasyonu Tahmin edilemeyen göç Northern blot karışık bir RNA populasyonu içerisinden istenilen bir RNA dizisinin belirlenmesini Western blot ise istenilen bir proteini bulmaya yarayan yöntemdir.Jel elektroforezi ise elektrik akımı verilerek hem DNA hem de proteinlerin boylarına yada büyüklüklerine göre belirli bir matrix üzerinde belirli bir noktaya göçüdür.

19
Nokta mutasyonların indüklenmesi
Her mutajen farklı bir mutasyon özgüllüğü yada önceliğine sahiptir DNA üzerindeki hedeflenen mutasyon bölgelerine yada mutasyona yatkınlık gösteren bölgelere sıcak nokta denir (hot spot) Mutajenler üç farklı mekanizmayla etkilerini gösterirler Baz başka bazla değişmesi Bazın değişmesi Bazın hasar görmesi
 Mutajenler üç farklı mekanizmayla etkilerini gösterirler. Baz başka bazla değişmesi. Bazın değişmesi. Bazın hasar görmesi.")
20
Bazın başka bazla değişmesi
Bazı kimyasal bileşikler normal azotlu bazlara benzerler ve normal bazın yerine geçerler bu bileşiklere baz analogları denir Bu analoglar normal bazlar gibi eşleşmezler Ve replikasyon sırasında yanlış nt lerin DNA yapısına eklenmesine neden olurlar

21
DNA yapısındaki her bir baz tautomer denen birkaç formda bulunur
Tautomerler izomerlerdir atomlarının pozisyonları ve atomları arasındaki bağları farklıdır Her form denge halindedir Keto form normal olarak DNA yapısında bulunan formdur İmino ve enol formları ise daha az rastlanan formlardır İmino yada enol tautomerler yanlış bazla eşleşirler ve mispair oluştururlar.

22
Bazların normal keto formlarının eşleşmesi

23
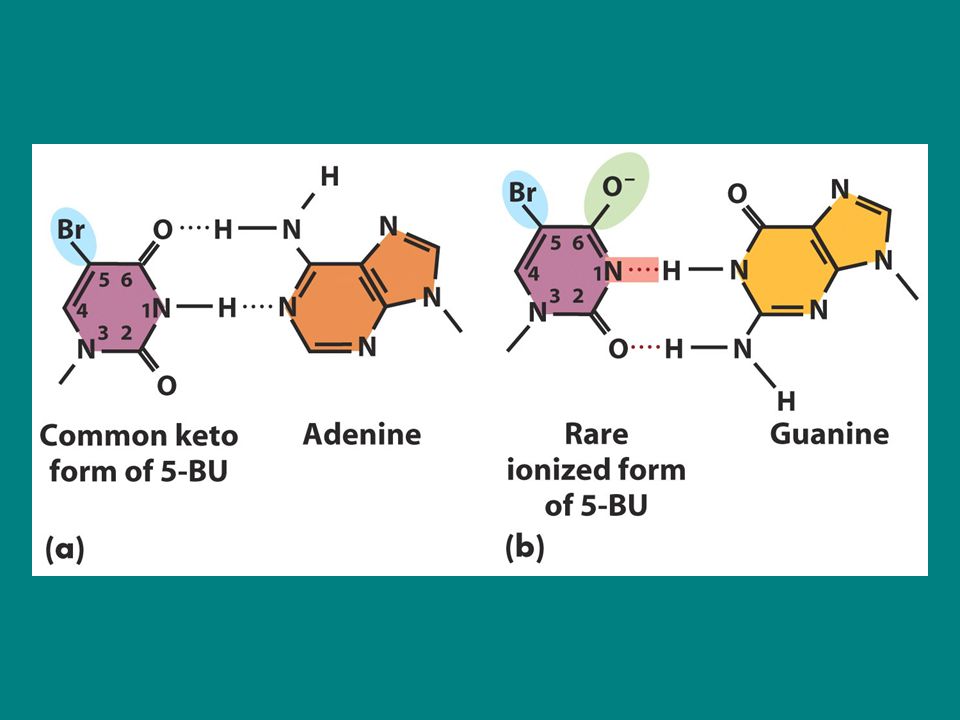
5-Bromo urasil Bazların yanlış eşleşmesi spontan yada bazlar iyonize olduklarında görülür 5-bromourasil (5-BU) timin analogudur ve timinde bulunan CH3 gurubu yerine karbon 5. pozisyonda bromin bulunur. 5-BU’nun mutajenik eylemi enolizasyon ve iyonizasyondan kaynaklanır. 5-BU’de bromin atomu baz eşleşmesi sırasında H bağı oluşturacak pozisyonda bulunmaz dolayısıyla 5BU timin bazı gibi adenin ile eşleşir ancak bromin atomu baz halkasında bulunan elektronların dağılımını önemli ölçüde değiştirir.çünkü 5BU ya enol yada iyonize forma dönüşür: enol ve iyonize form ise guanin ile eşleşir 5BU replikasyon sırasında enolizasyon yada iyonizasyonuna bağlı olarak G.C-A.T yada A.T-G.C transisyonuna neden olur.
 timin analogudur ve timinde bulunan CH3 gurubu yerine karbon 5. pozisyonda bromin bulunur. 5-BU’nun mutajenik eylemi enolizasyon ve iyonizasyondan kaynaklanır. 5-BU’de bromin atomu baz eşleşmesi sırasında H bağı oluşturacak pozisyonda bulunmaz dolayısıyla 5BU timin bazı gibi adenin ile eşleşir ancak bromin atomu baz halkasında bulunan elektronların dağılımını önemli ölçüde değiştirir.çünkü 5BU ya enol yada iyonize forma dönüşür: enol ve iyonize form ise guanin ile eşleşir. 5BU replikasyon sırasında enolizasyon yada iyonizasyonuna bağlı olarak G.C-A.T yada A.T-G.C transisyonuna neden olur.")
24
Az rastlanan sitozinin
İmino formu (C*) Az rastlanan timin İmino formu (T*) Mismatch ile sonuçlanan az rastlanan baz tautomerik formları
 Az rastlanan timin. İmino formu (T*) Mismatch ile sonuçlanan az rastlanan baz tautomerik formları.")
25
Az rastlanan adenin İmino formu (A*) Az rastlanan guanin İmino formu (G*)
 Az rastlanan guanin İmino formu (G*)")
27
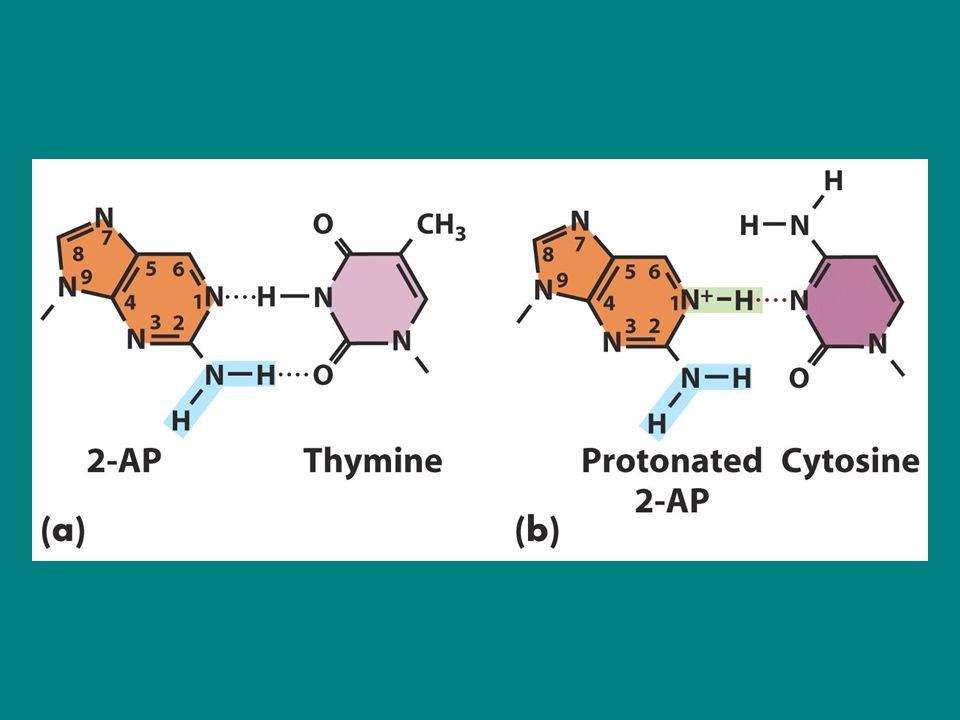
2-aminopurin (2-AP) 2AP adenin analogudur ve timinle eş oluşturur ve önemli mutajenlerdendir. Protonlandığı zaman 2AP sitozin ile yanlış eşleşir. Dolayısıyla timin ile eşleşen 2AP DNA yapısına katıldığında sonraki replikasyonda sitozin ile yanlış eşleşerek A.T-G.C transisyonuna neden olur. Yada eğer 2AP sitozin ile yanlış eşleşerek DNA yapısına katılmışsa G.C-A.T transisyonu görülür. Çünkü 2AP sonraki replikasyon döngüsünde timin ile eşleşir

29
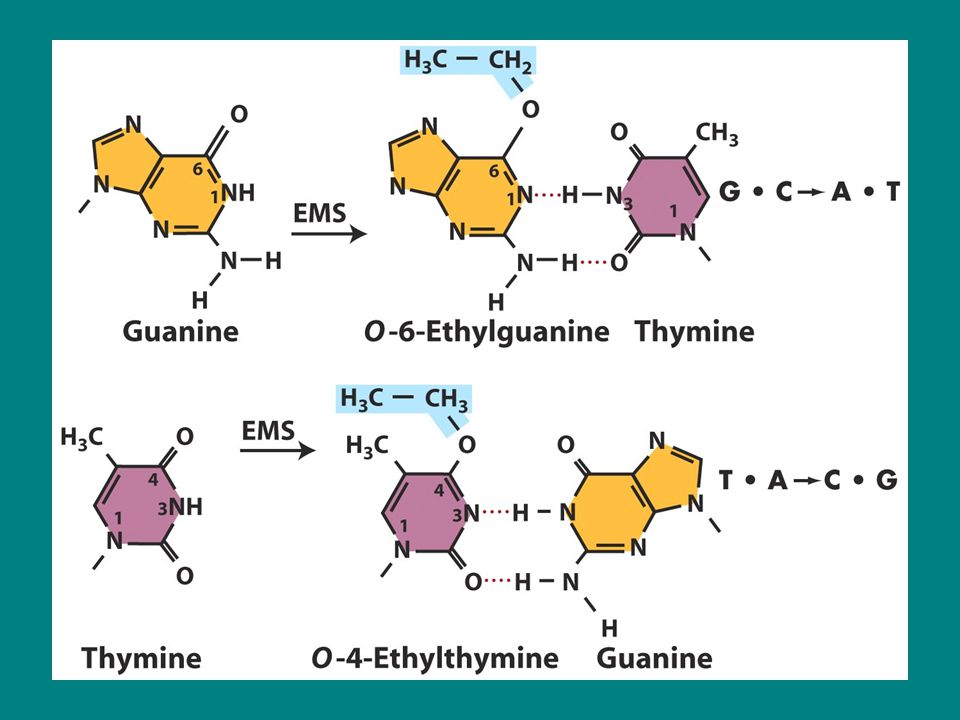
Baz değişimi Bazı mutajenler DNA yapısına katılmazlar ancak bazın kendisini değiştirirler ve yanlış eşleşmeye neden olurlar. Belirli alkile ajanlar bu yolakta kullanılır ör: Etilmetansülfonat (EMS) Nitrozoguanidin (NG) Bu ajanlar alkil gruplarını (EMS’de etil, NG örneğinde metil) her dört bazın çeşitli pozisyonlarına eklerler.guaninin 6.pozisyonunda yer alan oksijenine alkil grubu eklendiğinde O-6 alkil gunain meydana gelir bu da önemli mutasyonlardan biridir. Bu alkilasyon timin ile direkt yanlış eşleşmeye neden olur ve bir sonraki replikasyonda G.C-A.T transisyonuna yol açar Alkile ajanlar DNA sentezi sırasında eklenecek olan ntin bazını da modifiye ederler.
 Nitrozoguanidin (NG) Bu ajanlar alkil gruplarını (EMS’de etil, NG örneğinde metil) her dört bazın çeşitli pozisyonlarına eklerler.guaninin 6.pozisyonunda yer alan oksijenine alkil grubu eklendiğinde O-6 alkil gunain meydana gelir bu da önemli mutasyonlardan biridir. Bu alkilasyon timin ile direkt yanlış eşleşmeye neden olur ve bir sonraki replikasyonda G.C-A.T transisyonuna yol açar. Alkile ajanlar DNA sentezi sırasında eklenecek olan ntin bazını da modifiye ederler.")
31
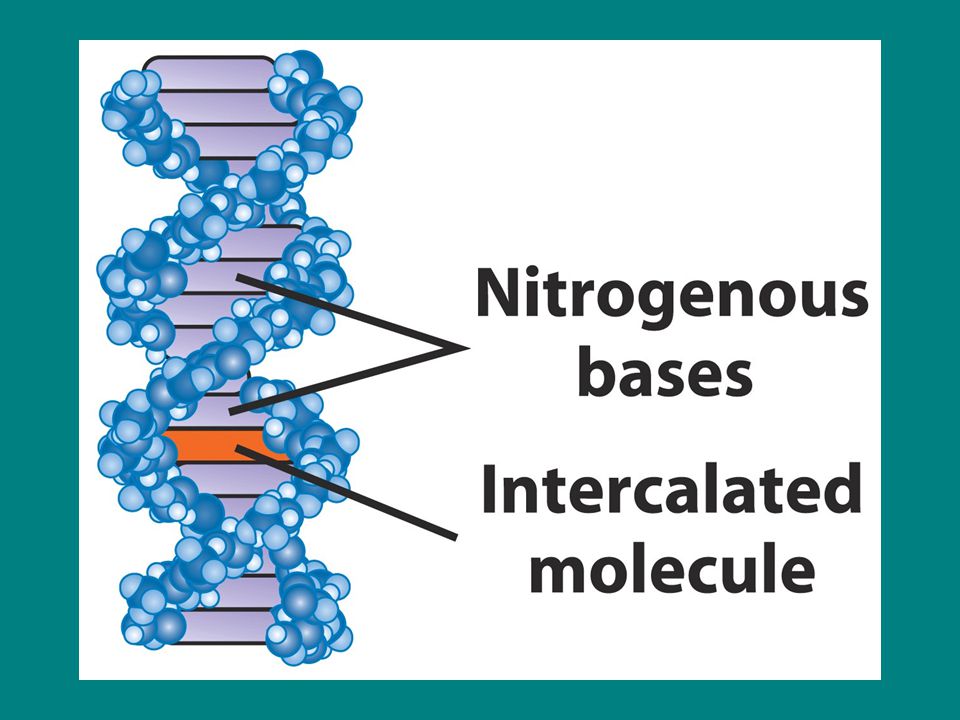
İntercalating ajanlar
Proflavin Akridin oranj ICR bileşikleri Bu bileşiklerin tamamı düz moleküllerdir ve baz eşleşmesini taklit eder ve DNA da bazlar arasına kayarak interkale olurlar. Bu interkalasyonda tek nt çifti insersiyon yada delesyonlarına neden olurlar.

34
Baz hasarları Birçok mutajen bir yada birkaç bazı hasara uğratır ve bazlar spesifik eşleşmelerini yapamazlar Sonuç replikasyonun bloke olmasıdır çünkü DNA pol DNA sentezini sürdüremez Prokaryot ve ökaryotlarda bu tür replikasyon blokajları nonspesifik bazlar eklenerek bypass edilir. E.colide bu süreç SOS sistemi aktivasyonunu gerektirir. Bu sistem ve diğer onarım sistemleri bir sonraki bölümde anlatılacaktır ancak burada kısa bir göz atılacaktır çünkü bu sistemlerin kendileri de DNA mutasyonuna neden olur.

35
SOS sistemi SOS adı önemli DNA hasarlarının varlığında hücrenin ölmesini önlemek için bir acil durumun varlığını tanımlamaktan gelir. Yani SOS hücrenin artık ölümden dönmek için son kozlarını oynamasıdır. 30 yıl boyunca SOS sisteminin nasıl mutasyon yarattığı merak edilmiştir çünkü DNA pol hasarlı noktayı bypass etmektedir.

36
UV birçok organizmada baz hasarlarına neden olur
UV’ye maruz kalan E.coli hücrelerinde Farklı bir E.coli mutant tipi 1970lerde belirlendi. Bu durum bu hücrelerin bazı genlerinin UVye maruz kaldığında mutasyon ürettiği sonucunu ortaya çıkardı. UV ile indüklenen mutasyonların DinB, UmuC yada UmuD’ genleri mutant olduğunda ortaya çıkmadıkları görüldü. Son zamanlarda bu genlerin iki tane hata-eğilimli DNA pol kodladıklarını ortaya koydu DinB DNA pol IV, UmuC ve UmuD’ DNA pol V altbirimlerini kodlar Bu polimerazlar replikasyon sırasındaki blokajın üstesinden hasarlı DNA’nın karşısındaki zincirinde ntleri ekleyerek gelirler. Hata-eğilimli pol lar (ErrorProne pol veya sloppy kopyalayıcılar)insandan mayaya kadar birçok canlıda tanımlanmıştır.Bu canlılarda hasarı tolere eden mekanizma diğer adıyla translezyon DNA sentezine katkıda bulunurlar ve bu sistemler SOS bypass sistemine benzer.
insandan mayaya kadar birçok canlıda tanımlanmıştır.Bu canlılarda hasarı tolere eden mekanizma diğer adıyla translezyon DNA sentezine katkıda bulunurlar ve bu sistemler SOS bypass sistemine benzer.")
37
İnsanda Hata-eğilimli DNA pol etki mekanziması:
translezyon DNA sentezi DNA pol ŋ (eta) ve DNA pol ι (iota)
 ve DNA pol ι (iota)")
38
Hata-eğilimli DNA pol duran replikasyon makinesine bağlanır

39
Bağlanma şekil değişimini artırır ve replikasyon başlar ve pol eta nt ekler

40
Pol eta ayrılır ve pol iota yanlış bazları hasarlı DNA’nın karşısındaki zincire eklemeye devam eder

41
SOS sisteminde ilk evre RecA denen proteinin sentezinin aktivasyonudur
DNA pol III hasarlı bölgede durduğunda polimerazın önünde kalan DNA bölgesi açılmaya devam eder ve SSBPler bu tek zincirli DNA’ya bağlanmaya devam eder Daha sonra RecA SSBP’ye bağlanır ve protein-DNA filamenti oluşturur.RecA filamenti bu proteinin biyolojik olarak aktif formudur ve bir sinyal görevi görerek hata-eğilimli pol’ın indüklenmesini sağlar ve bu durmuş çatal polimeraz için çekici hale gelir Kararlı baz çifti oluşmasını önleyen mutajenler SOS sistemine bağımlıdırlar çünkü yanlış bazların eklenmesi SOS siteminin aktive olmasına bağlıdır. SOS bağımlı mutajenler çok önemlidir çünkü birçoğu kansere neden olan ajanlardır (karsinojenler) ör: UV ve aflatoxin B1
 ör: UV ve aflatoxin B1.")
42
UV IŞIĞI UV foto-ürün denen DNA değişimlerine yol açar. İki farklı lezyon oluşturarak aynı zincirde komşu pirimidinleri birleştirir. Bu lezyonlar Siklobütan pirimidin fotodimeri ve 6-4 fotoürün oluşturmasıdır. Siklobütan pirimidin dimeri örneğinde UV dört üyeli siklobütil halkası oluşumunu aynı DNA zincirindeki komşu pirimidinler arasında 5,6 çift bağ üzerinde harekete geçerek stimüle eder. 6-4 fotoürün yapısı iki komşu pirimidinin C-6 ve C-4 atomları arasında oluşur (çoğunlukla 5’CC3’ ve 5’TC3’) Pirimidin dimerleri heliks yapısını bozar ve SOS sistemini aktive eder Dimerin 3’ pozisyonunda UV fotoürünlerinin karşısına yanlış bazlar eklenir. C-T transisyonu en sık rastlanandır ama UV transversiyon,frameshift büyük duplikasyonlar ve delesyonlarda yapabilir.
 Pirimidin dimerleri heliks yapısını bozar ve SOS sistemini aktive eder. Dimerin 3’ pozisyonunda UV fotoürünlerinin karşısına yanlış bazlar eklenir. C-T transisyonu en sık rastlanandır ama UV transversiyon,frameshift büyük duplikasyonlar ve delesyonlarda yapabilir.")
44
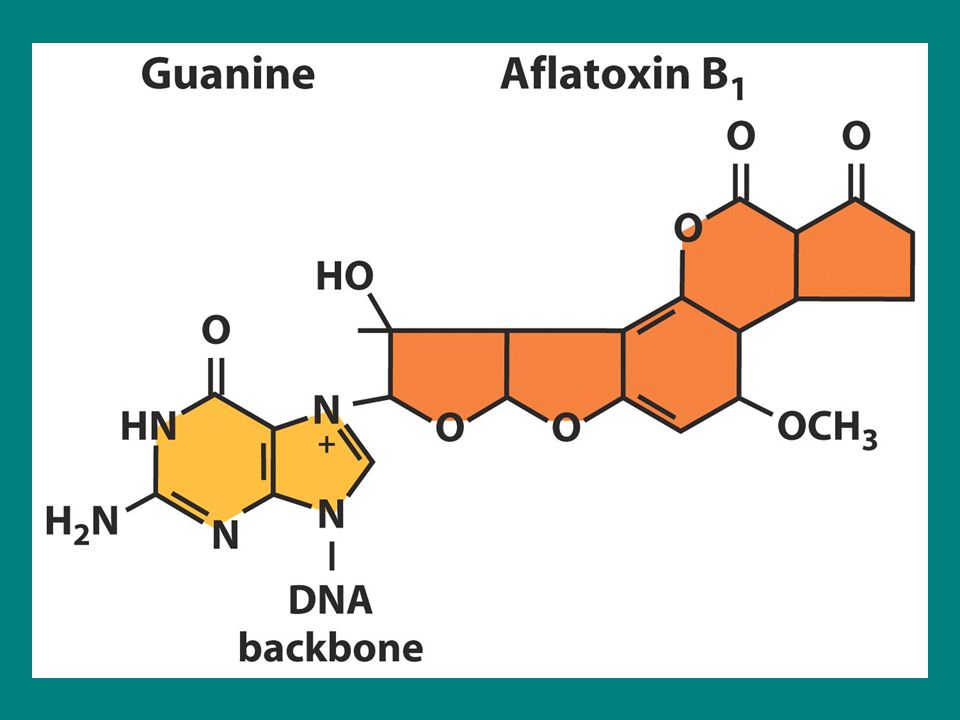
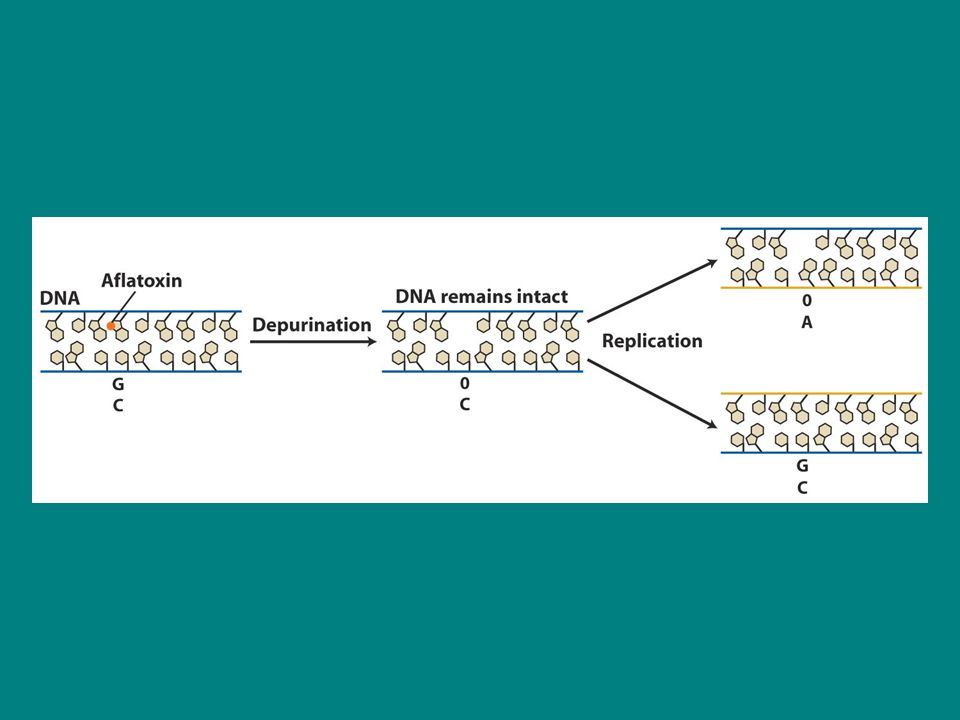
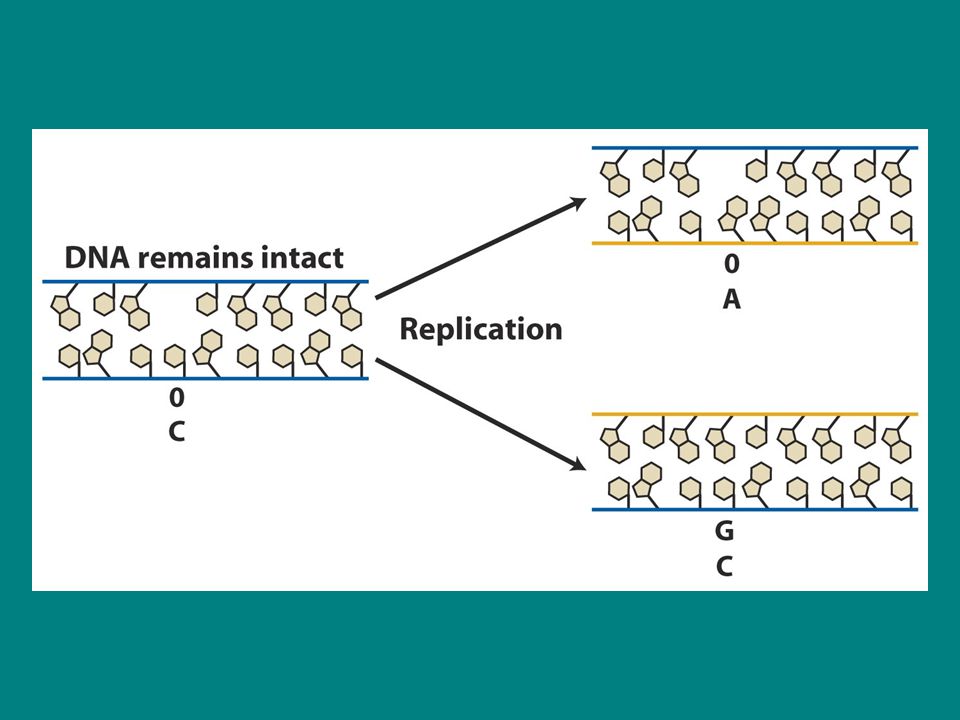
Aflatoksin B1 AFB1 güçlü bir karsinojendir ve bir mantarla enfekte fıstıklardan izole edilmiştir. AFB1 N-7 pozisyonunda guanine tutunur ve baz ile şeker arasındaki bağı koparır baz serbest kalır ve apurinik bölge oluşturur. SOS bypass sistemi böyle bir hasarın olduğu yerin karşısına adenin ekler Sonuç G.C-T.A transversiyonudur.

49
Spontan mutasyonlar Spontan mutasyonlar dış bir uyaran tarafından indüklenmektemidir? Yoksa gen varyantları düşük frekanstamı bulunmaktadır? Bu soruya yanıt bakterilerde spesifik çevresel faktörlere direnç gösteren genleri mutasyona uğratmakla elde edilmiştir. Normal doğal tip bakteriler bu tür mutasyonları tolere etmektedir.

50
Luria- delbrück yükseltme azaltma (fluctuation) deneyi (iniş çıkış deneyi)
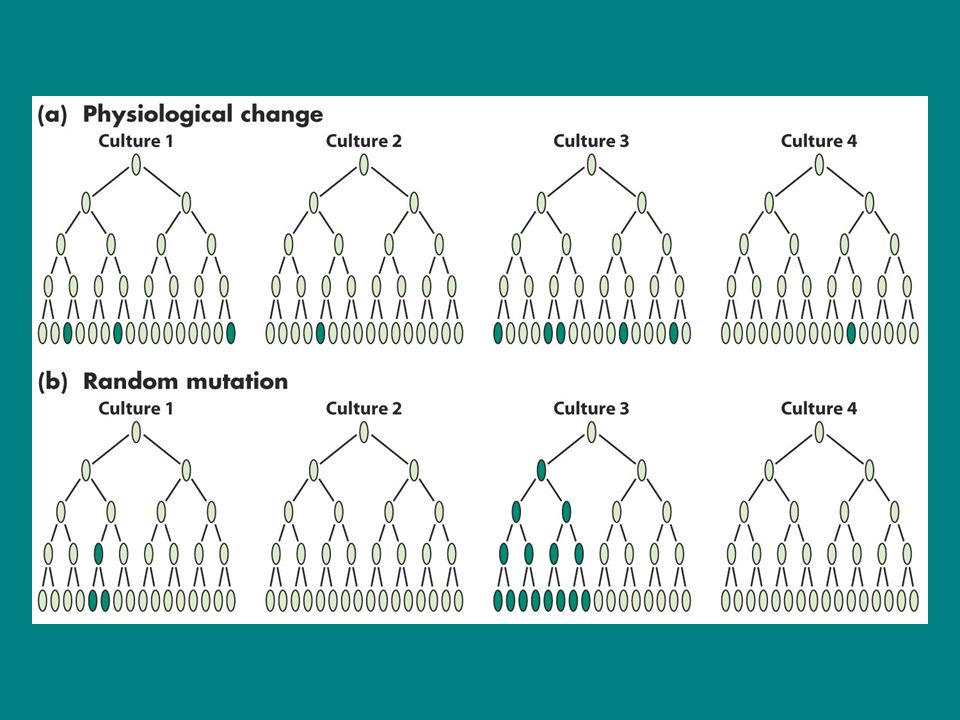
1943te araştırıcılar bu deneyi yaparlarken E.coli hücrelerinin besin ortamındaki agara ekimiyle birlikte T1 fajı da eklenirse bu fajlar bakterileri enfekte ettiği ve öldürdüğü biliniyordu Ancak nadir olarak bazı bakteriler faj enfeksiyonuna direnç göstermektedir bu koloniler kararlı ve gerçek mutantlardır. Ancak o dönemde bu mutantların spontanmı yoksa rastgele zamanla oluşup oluşmadıkları bilinmiyordu. Ayrıca fajların bu bakterilerden bazılarını resiztan hale getirip getirmedikleri de bilinmiyordu Luria’ya göre eğer mutasyon spontansa farklı kültürlerde farklı zamanlarda mutantlar oluşmalıydı ve kültür başına oluşan resistan koloni sayısı yüksek çeşitlilik göstermeliydi (yani inişli çıkışlı bir sayıda olmalıydı) Bu araştırıcılar deneyi şöyle kurdular 20 tane küçük kültürü herbiri birkaç koloni ile inoküle ettiler ve hücre sayısının 108 hücre/ml oluncaya kadar büyüttüler Aynı zamanda daha büyük bir kültürü bakteriyle inoküle edip aynı hücre sayısına ulaşıncaya kadar büyüttüler 20 küçük kültür ve büyük kültürden elde edilen 20 örnek faj bulunan ortama ekildi 20 küçük kültür önemli sayıda dirençli koloni oluştururken büyük kültürlerden elde edilen örneklerde varyasyon daha az görüldü. 20 küçük kolonili örnekten 11inde dirençli koloni görülmedi kalanlarda, 1, 1, 3,5, 5, 6, 35, 64 ve 107. büyük kültürde ise dirençli koloni sayısı arasında değişti Eğer faj mutasyonu indüklemiş olsaydı küçük kültürlerde aynı sayıda mutasyon görülürdü.en iyi açıklama rastgele mutasyon oluşmuştu.erken mutasyona uğrayan hücreler daha fazla sayıda mutant koloni verirken geç mutasyona uğrayan hücreler daha az mutant hücre vermiştir. Buna göre resistan hücreler çevresel ajanlar tarafından (burada faj) oluşturulmaktan öte onlar tarafından seçilirler
 Bu araştırıcılar deneyi şöyle kurdular. 20 tane küçük kültürü herbiri birkaç koloni ile inoküle ettiler ve hücre sayısının 108 hücre/ml oluncaya kadar büyüttüler. Aynı zamanda daha büyük bir kültürü bakteriyle inoküle edip aynı hücre sayısına ulaşıncaya kadar büyüttüler. 20 küçük kültür ve büyük kültürden elde edilen 20 örnek faj bulunan ortama ekildi. 20 küçük kültür önemli sayıda dirençli koloni oluştururken büyük kültürlerden elde edilen örneklerde varyasyon daha az görüldü. 20 küçük kolonili örnekten 11inde dirençli koloni görülmedi kalanlarda, 1, 1, 3,5, 5, 6, 35, 64 ve 107. büyük kültürde ise dirençli koloni sayısı arasında değişti. Eğer faj mutasyonu indüklemiş olsaydı küçük kültürlerde aynı sayıda mutasyon görülürdü.en iyi açıklama rastgele mutasyon oluşmuştu.erken mutasyona uğrayan hücreler daha fazla sayıda mutant koloni verirken geç mutasyona uğrayan hücreler daha az mutant hücre vermiştir. Buna göre resistan hücreler çevresel ajanlar tarafından (burada faj) oluşturulmaktan öte onlar tarafından seçilirler.")
51
Replika plate Seleksiyondan önce bir populasyonda mutantların varlığı direkt gösterilebilirmi Lederberg 1952-replika plating yöntemi Bakteriler seçici olmayan (faj olmayan) ortama ekilir ve herbir hücreden bir koloni büyür bu plate master plate olarak adlandırılır. Kadife bir kumaş bu platin üzerine konur ve kolonilerden birer kısmı alınarak başka bir plağa konur ve ana platein kopyası –replikası yapılır ancak bu plate seçici ortamdır yani faj içerir.beklendiği gibi dirençli koloniler replikalar üzerinde görülebilir. Eğer mutasyon seçici ortama konduktan sonra meydana gelmiş olsaydı herbir platein paterni rastgele olurdu Sonuç mutasyonlar seçici ortama maruz bırakmadan önce meydana gelmektedir. Mutasyonlar rastgele meydana gelirler herhangi bir zamanda herhangi bir hücrenin alleli mutasyona uğrayabilir.
 ortama ekilir ve herbir hücreden bir koloni büyür bu plate master plate olarak adlandırılır. Kadife bir kumaş bu platin üzerine konur ve kolonilerden birer kısmı alınarak başka bir plağa konur ve ana platein kopyası –replikası yapılır ancak bu plate seçici ortamdır yani faj içerir.beklendiği gibi dirençli koloniler replikalar üzerinde görülebilir. Eğer mutasyon seçici ortama konduktan sonra meydana gelmiş olsaydı herbir platein paterni rastgele olurdu. Sonuç mutasyonlar seçici ortama maruz bırakmadan önce meydana gelmektedir. Mutasyonlar rastgele meydana gelirler herhangi bir zamanda herhangi bir hücrenin alleli mutasyona uğrayabilir.")
55
Spontan mutasyon mekanizmaları
Replikasyon hataları Spontan lezyonlar Transpoze elementlerin insersiyonu Spontan mutasyonlar nadir görülür ve onların altındaki nedeni belirlemekde zordur

56
Spontan lezyonlar Doğal olarak DNAnın hasara uğramasına spontan lezyonlar denir ve bu hasarlar mutasyona neden olurlar En sık rastlananları Depurinasyon (daha sık gözlenen) deaminasyon
 deaminasyon.")
57
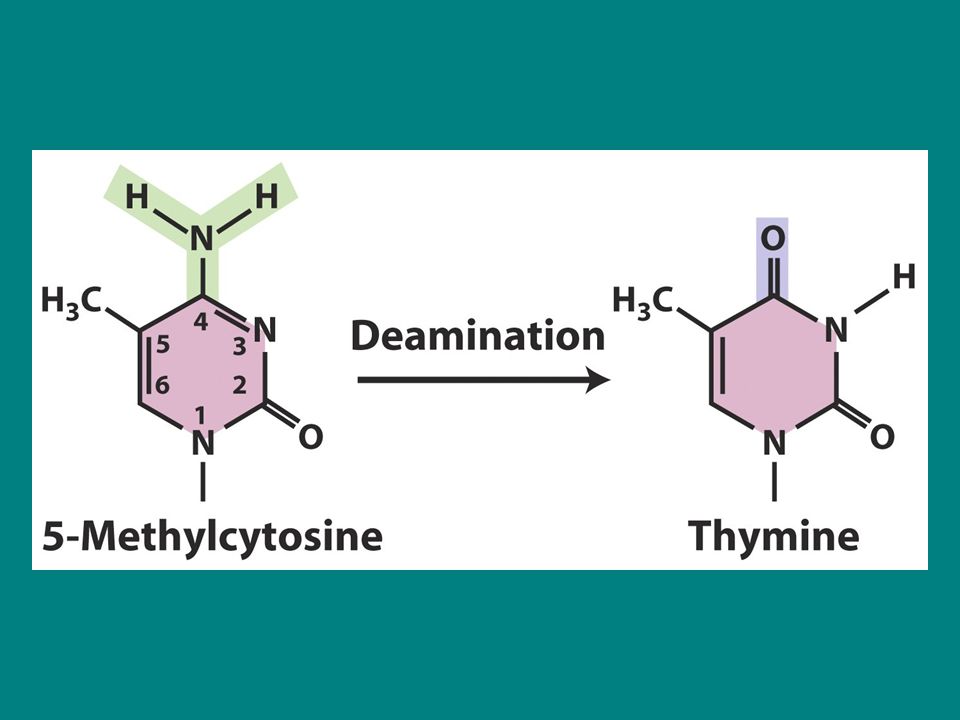
deaminasyon Sitozin deaminasyonu urasil üretir
Düzeltilmedikçe replikasyon sırasında urasil adenin ile eşleşir ve GC çifti AT ile değişir. 5-metil sitozin deaminasyonu da görülür çünkü pro ve ökaryotlarda bazı bazlar metillenirler. 5-metilC deaminasyonu timin üretir (5-metilurasil) dolayısıyla C-T transisyonu deaminasyonla üretilir.
 dolayısıyla C-T transisyonu deaminasyonla üretilir.")
58
Oksidatif hasarlı bazlar
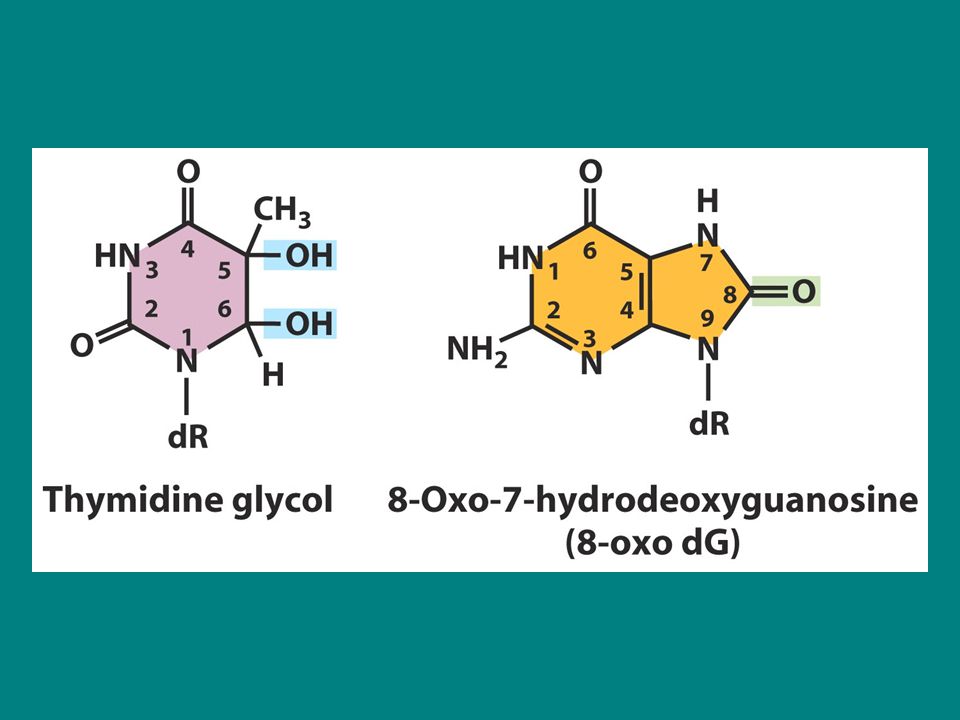
Bu bazlarda spontan mutasyonlara neden olur Aktif oksijen türleri süperoksit radikalleri (O2.-) hidrojen peroksit (H2O2) ve hidroksil radikalleri (.OH) normal aerobik solunumun yan ürünleridir. Bu yan ürünler oksidatif hasarlara neden olurlar ve bu mutasyonlar çeşitli hastalıklarda önemli rol oynarlar Oksidatif hasarlara iki örnek bozuk timidin birimi diğeri ise bozuk guanozin birimidir. Timidin glikol ve 8-oxo-7 hidroksi deoksiguanozin (8-oxodG yada GO). GO adenin ile yanlış eşleşir ve yüksek sıklıkta G-T transversiyonuan neden olur.
 hidrojen peroksit (H2O2) ve hidroksil radikalleri (.OH) normal aerobik solunumun yan ürünleridir. Bu yan ürünler oksidatif hasarlara neden olurlar ve bu mutasyonlar çeşitli hastalıklarda önemli rol oynarlar. Oksidatif hasarlara iki örnek bozuk timidin birimi diğeri ise bozuk guanozin birimidir. Timidin glikol ve 8-oxo-7 hidroksi deoksiguanozin (8-oxodG yada GO). GO adenin ile yanlış eşleşir ve yüksek sıklıkta G-T transversiyonuan neden olur.")
62
depurinasyon Aflatoksin depurinasyonu indükler (purin bazı kaybı) ancak depurinasyon spontan da olabilir Memeli hücreleri spontan olarak 37 derecede 20 saatlik bir üreme periyodunda yaklaşık purinini kaybeder Eğer bu mutasyonlar varsa replikasyon sırasında önemli DNA kayıplarına neden olur çünkü baz olmayan yerin karşısına baz eklenemez. Ancak daha önce anlatıldığı gibi buraya bir baz eklenir ama mutant bir baz olur.

64
DNA replikasyon Hataları
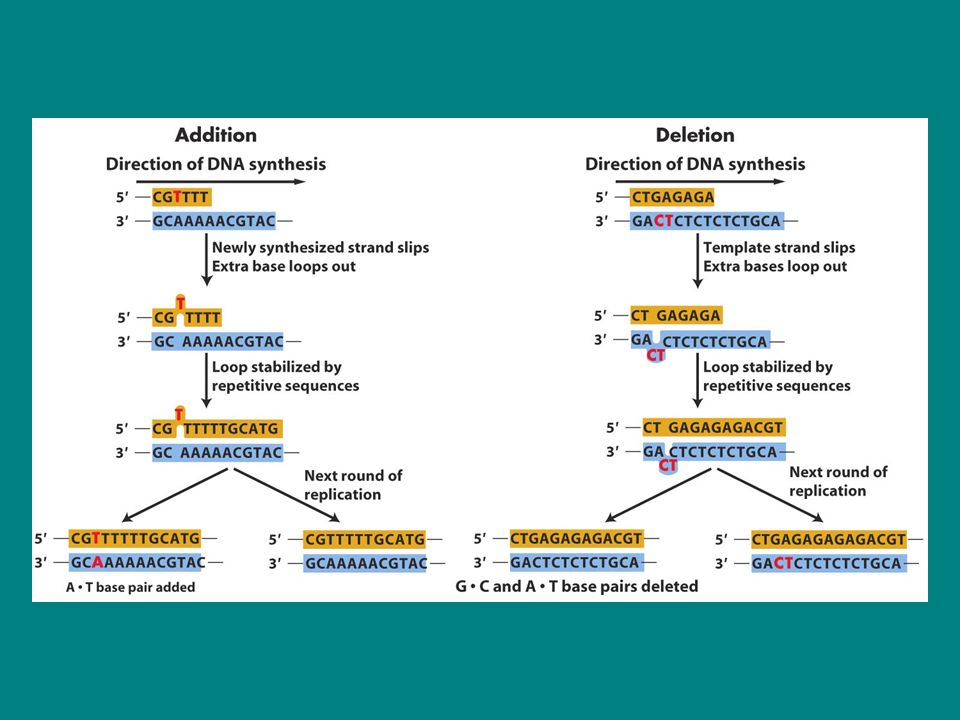
Baz değişimi Hiçbir kimyasal reaksiyon mükemmel değildir dolayısıyla DNA sentezi sırasında da yanlış nt çiftleri oluşturulabilir. Baz insersiyonu ve delesyonu Replikasyon sırasında indel mutasyonlarda görülür eğer üçlü nt biçiminde değilse frame kaymasına neden olur Bu eklenti yada çıkarılmalar replikasyon kayması olarak da bilinir. Genellikle art arda dizilmiş aynı nt lerin olduğu bölgelerde bu kaymalar meydana gelir.

66
İnsanda spontan mutasyonlar-trinükleotid tekrarları
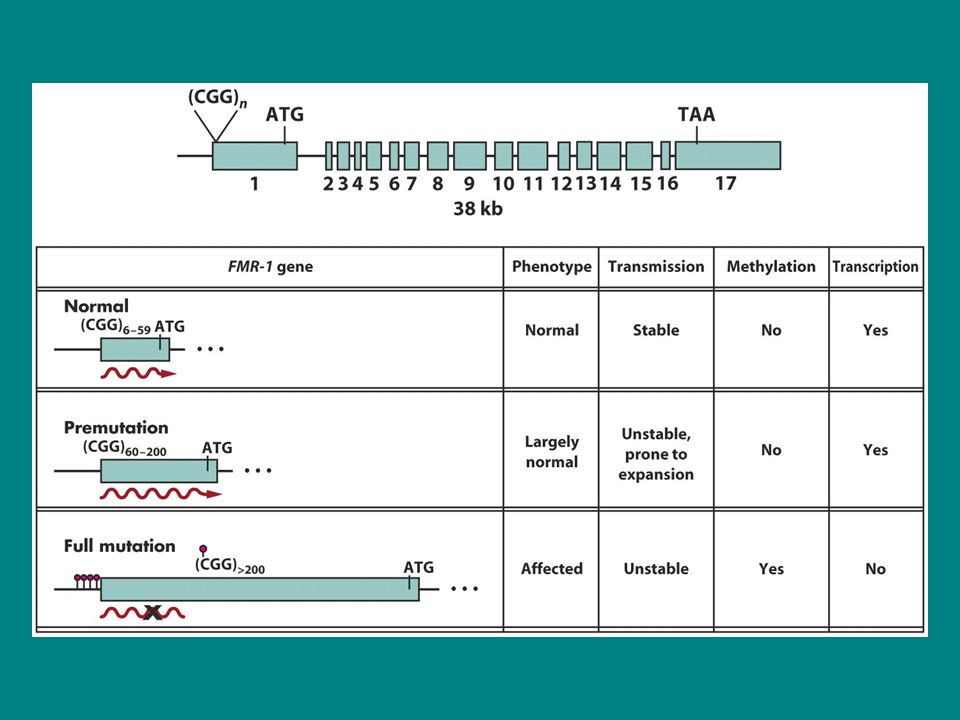
İnsanda görülen birçok hastalığın etkeni karmaşık mutasyonlardan kaynaklanır. Bunlardan en önemlilerinden biri kısa tekrarların duplikasyonlarıdır. Birçok kalıtsal hastalıkdan sorumlu genetik defektin altında yatan mekanizma 3lü nt dizilerinin tekrarıdır (trinucletid repeat) Fragil X sendromu buna örnektir 1/1500 erkek ve 1/2500 dişide görülür sitogenetik olarak kromozom incelenmesinde x kromozomunda kırılganlıklar görülebilir. Frajil X FMR1 genindeki CGG tekrarlarındaki değişim sonucu ortaya çıkar bu gen transkribe olur ama translase olamaz. Tekrar sayısı hastalık fenotipiyle nasıl doğru orantılı biçimde seyreder? İnsanlar doğal olarak FMR1 genindeki CGG tekrarlarında çeşitlilikler gösterir ve bu tekrarların sayısı 6-54 arasında değişir. Bazan etkilenmeyen ebeveynlerden hatta büyük baba/anneler kendinden sonraki kuşaklarda etkilenmiş çocuk sahibi olurlar.Bu hasta bireylerde tekrar sayıları e ulaşır. Etkilenmemiş ebevynlerde de bazan sayı arasında değişir. Dolayısıyla bu etkilenmemiş ebevynler premutasyonu taşımaktadırlar. Premutantlardaki Bu tekrarlar hastalık fenotipinin ortaya çıkması için yeterli değildir ancak bu kişiler oldukça yatkın kişilerdir.
 Fragil X sendromu buna örnektir. 1/1500 erkek ve 1/2500 dişide görülür sitogenetik olarak kromozom incelenmesinde x kromozomunda kırılganlıklar görülebilir. Frajil X FMR1 genindeki CGG tekrarlarındaki değişim sonucu ortaya çıkar bu gen transkribe olur ama translase olamaz. Tekrar sayısı hastalık fenotipiyle nasıl doğru orantılı biçimde seyreder İnsanlar doğal olarak FMR1 genindeki CGG tekrarlarında çeşitlilikler gösterir ve bu tekrarların sayısı 6-54 arasında değişir. Bazan etkilenmeyen ebeveynlerden hatta büyük baba/anneler kendinden sonraki kuşaklarda etkilenmiş çocuk sahibi olurlar.Bu hasta bireylerde tekrar sayıları e ulaşır. Etkilenmemiş ebevynlerde de bazan sayı arasında değişir. Dolayısıyla bu etkilenmemiş ebevynler premutasyonu taşımaktadırlar. Premutantlardaki Bu tekrarlar hastalık fenotipinin ortaya çıkması için yeterli değildir ancak bu kişiler oldukça yatkın kişilerdir.")
68
Tekrar mekanizması DNA sentezi sırasında eşleşme kaymasından kaynaklanır.insanda 50 tekrardan sonra DNA pol enziminin doğru sentez yapmadığı ileri sürülmektedir.yani 50 tekrar DNA pol için eşik değeridir.

69
Huntington Disease Bir gendeki trinükleotid tekrarlarının genişlemesinden kaynaklanır. HD geninde ekzonlarda tekrarlar görülür ve mutasyonlar tekrar sayısı arttıkça artar ve hastalığın prognozu tekrar sayısı arttıkça kötüleşir. HD ve Kennedy disease (X-bağlı spinal ve muskular atrofi) CAG dizilerinin genişlemesiyle meydana gelir Normal kişiler CAG tekrarına sahiptir hastalar ise ortalama 46 tekrara sahiptir KD da sürekli kas zayıflığı ve atrofisi görülür ve androjen reseptörünü kodlayan gendeki tekrar sayıları artmıştır. Tüm bu trint tekrar hatsalıklarında genel olarak nörodejenerasyon görülür (sinir sisteminde hücre ölümü) Tekrarlar okuma çerçevesi içerisinde yer alır FMR1 geninde ise tekrarlar genin 5’ ucuna yakın biryerde görülür transkripsiyona başlama noktasından önce. Dolayısıyla buradaki mekanizma bu dizilerin hipermetilasyonu mekanizmasıyla açıklanabilir.Hipermetilasyon genleri sessiz kılabilir. Tekrar genişlemesi kromatin yapısında değişime neden olarak mutant genin transkripsiyonunu baskılayabilir.
 CAG dizilerinin genişlemesiyle meydana gelir. Normal kişiler CAG tekrarına sahiptir hastalar ise ortalama 46 tekrara sahiptir. KD da sürekli kas zayıflığı ve atrofisi görülür ve androjen reseptörünü kodlayan gendeki tekrar sayıları artmıştır. Tüm bu trint tekrar hatsalıklarında genel olarak nörodejenerasyon görülür (sinir sisteminde hücre ölümü) Tekrarlar okuma çerçevesi içerisinde yer alır. FMR1 geninde ise tekrarlar genin 5’ ucuna yakın biryerde görülür transkripsiyona başlama noktasından önce. Dolayısıyla buradaki mekanizma bu dizilerin hipermetilasyonu mekanizmasıyla açıklanabilir.Hipermetilasyon genleri sessiz kılabilir. Tekrar genişlemesi kromatin yapısında değişime neden olarak mutant genin transkripsiyonunu baskılayabilir.")
Benzer bir sunumlar
















>")


 A) HÜCREDE YAPI VE CANLILIK OLAYLARININ YÖNETİMİ NASIL SAĞLANIR? Hücrede hücre yapısının oluşması ve devamlılığı ile canlılık olaylarının yürütülmesi.>")

>")





